
基于转录组测序的文心兰花香形成分析
2022-03-11 05:41:04陈艺荃方能炎叶秀仙罗远华钟淮钦黄敏玲樊荣辉
核农学报 2022年3期
陈艺荃 方能炎 叶秀仙 罗远华 钟淮钦 黄敏玲,* 樊荣辉,*
(1 福建省农业科学院农业工程技术研究所,福建 福州 350003;2 福建省农业科学院作物研究所,福建 福州 350013)
花香挥发物能吸引昆虫进行授粉繁殖,有利于防御病原体、寄生虫和食草动物的侵害[1-2],其主要由花器官在特定花发育时期释放产生[3]。迄今已有来自100种植物的大约1 700种花香挥发物被鉴定[2]。
花香挥发物主要由萜类化合物、苯丙酸类/苯类和脂肪酸衍生物这三类化合物组成[2-3]。萜类化合物是第一大类花香挥发物,主要由单萜(C10),倍半萜(C15)和二萜(C20)组成[4-5]。挥发性萜类化合物由两种常见的异戊二烯前体合成,即异戊烯焦磷酸酯(isopentenyl diphosphate, IPP)和二甲基烯丙基焦磷酸酯(dimethylallyl pyrophosphate, DMAPP)。在细胞溶质中,IPP和DMAPP由甲羟戊酸(mevalonic acid, MVA)途径的乙酰辅酶A生成,并在法呢基二磷酸合酶(farnesyl diphosphate synthase, FPPS)的催化下形成法呢基二磷酸(farnesyl diphosphate, FPP)。然后,细胞溶质/线粒体中的萜类合酶(terpene synthase, TPSs)将FPP转化为倍半萜[6-7]。在质体中,IPP和DMAPP由甲基赤藓醇磷酸(mevalonate pathway, MEP)途径中的丙酮酸和甘油醛-3-磷酸合成,并通过香叶基焦磷酸合酶(geranyl pyrophosphate synthase, GPPS)转化为香叶基焦磷酸(geranyl pyrophosphate, GPP), 随后,质体中的TPS催化GPP转化为各种单萜和二萜[8]。TPS基因家族被分为7个亚家族,分别为TPS-a~g[9-11],目前,已在拟南芥(Arabidopsisthaliana)[12]、玫瑰(Rosehybrida)[13]、薰衣草(Lavandulaangustifolia)[14]、金鱼草(Antirrhinummajus)[15]、烟草(Nicotianasuaveolens)[16]和依兰(Canangaodorata)[17]等植物中获得了TPS。
高等植物中萜类挥发物的释放受到TPS基因表达模式的调控[6]。金鱼草中,花香挥发物的释放量随TPS表达水平的升高而逐渐增加[16];晚香玉中,挥发性萜类的昼夜释放节律与TPS基因的表达密切相关[18]。TPS被认为是萜类化合物形成的关键基因。
文心兰(Oncidium)又名跳舞兰,原产墨西哥、西印度群岛、巴西等地,是兰科中的一大属,原种多达 750 种以上[19]。文心兰中多数品种无香气,而一些盆栽品种,如香水文心、梦香等香气浓郁。目前对文心兰花香的研究只集中在香水文心等花香成分的分析[20-22],鲜见花香形成分子水平等更深入的解析。本研究以白梦香,黄梦香和香水文心为材料,探索香气形成的分子机制,旨在为文心兰花香分子育种奠定基础。
1 材料与方法
1.1 试验材料
试验材料为福建省农业科学院花卉种质资源圃种植的文心兰。取香水文心(Onc.Sharry Baby)、白梦香(Onc.Twinkfe White Fantasy)、黄梦香(Onc.Twinkfe Yellow Fantasy)盛花期花朵用于转录组测序,取香水文心、白梦香、黄梦香花蕾前期、花蕾中期、盛花期花朵用于花香成分测定及实时荧光定量PCR(quantitative real-time PCR, qRT-PCR)检测。

表1 文心兰3个品种花香等级及描述Table 1 Aroma level of floral scents from three varieties of Oncidium orchid
1.2 顶空-固相微萃取和气相色谱分析
使用顶空-固相微萃取(headspace solid-phase microextraction, HS-SPME)方法收集挥发性化合物。称取1 g花朵样品密封于20 mL提取瓶中并在50℃条件下平衡10 min。使用SPME纤维[聚二甲基硅氧烷(polydimethylsiloxane,PDMS),直径65 μm]提取挥发性化合物并吸附30 min。然后,使用Shimadzu GCMS-TQ8040系统(日本岛津)对捕获的花香化合物进行气相色谱分析(gas chromatography-mass spectrometry, GC-MS)。使用Rxi-5Sil MS毛细管柱(30 m×0.25 mm×0.25 μm)分离挥发性化合物,以氦气作为载气,流速为1 mL·min-1。柱操作条件如下:初始50℃保持2 min,再以4℃·min-1的速率升至170℃,然后以20℃·min-1的速率升至270℃,最后保持2 min。选用α-Terpinene为标准品,采用内标法。每个样品3次重复。
1.3 RNA提取、文库构建和RNA-Seq
收集盛花期的整朵花进行转录组测序,每个样品3次重复。使用通用RNA提取试剂盒(Bioteke Corporation,大连)提取总RNA,应用NanoDrop 2000 UV-vis分光光度计(Thermo Scientific,美国)和Agilent 2100生物分析仪(Agilent Technologies,美国)进行RNA质量和浓度测定。文库由北京百迈克生物科技有限公司使用mRNA-Seq样品制备试剂盒(Illumina,美国)构建。最后,使用Illumina HiSeq TM 4000平台对文库进行测序。
1.4 组装和功能注释
收集四组样品的原始数据(raw reads),去除低质量数据(reads)。应用Trinity软件通过重叠区域将高质量过滤后数据(clean reads)延伸为更长的重叠群(contigs),通过对端连接进一步组装成转录本(transcripts),然后聚类成单基因(unigenes)[23]。通过序列相似性,将所有组装的unigenes与公共数据库进行比对,E值阈值为10-5。七大数据库包括Nr (NCBI non-redundant protein), Nt (non-redundant nucleotide databases), GO(gene Ontology), KOG (eukaryotic orthologs groups), KEGG(kyoto encyclopedia of genes and genomes), Pfam(protein family), Swiss-Prot(Swiss-Prot protein database)。
1.5 表达注释
使用Bowtie2软件将修剪后的reads对齐到组装的转录组[24]。应用RSEM(RNA-Seq by expectation maximization)评估每个基因的丰度。应用每千个碱基的转录每百万映射读取的碎片(fragments per kilobase of exon model per million mapped reads,FPKM)计算每个基因的数据计数(read counts)[25]。应用DESeq软件鉴定文库中的差异基因[26]。绝对错误率(Error rate)<0.01和倍数变化值(Fold change)≥2作为阈值以确认表达水平的显著差异。
1.6 进化树构建
在NCBI里查找其他物种的TPS核苷酸序列,利用MEGA软件对TPS进行系统进化树的构建,参照Trapp等[27]和Fan等[28]的方法,对TPS家族进行分类。
1.7 qRT-PCR分析
使用荧光染料SYBR Green(TaKaRa, 大连)在ABI 7500实时PCR系统(Applied Biosystems,美国)上进行qRT-PCR分析。应用PrimerPremier 5.0软件设计引物。Actin作为内参基因。qRT-PCR反应条件及体系按TB Green©Premix Ex TaqTMⅡ (Tli RNaseH Plus)(TaKaRa, 大连)试剂盒方法,每个反应3次重复。使用2-ΔΔCt方法计算相对表达量。

表2 qRT-PCR所用引物序列Table 2 Primer sequences of qRT-PCR
2 结果与分析
2.1 主要花香挥发物鉴定
GC-MS检测结果表明,香水文心、黄梦香和白梦香3个品种中,萜类挥发物及其衍生物是主要花香挥发物(表3)。3个品种花香挥发物总相对含量均随花的发育逐渐上升,到盛花期含量最高(图1)。白梦香、黄梦香和香水文心3个品种的主要花香挥发物存在差异,白梦香的主要成分为芳樟醇(15.76 μg·h-1)和反式橙花醇(11.50 μg·h-1),均在盛花期含量最高(图2、表3);黄梦香的主要成分为芳樟醇(27.13 μg·h-1)、 (E)-2-甲基巴豆酸异戊酯(27.08 μg·h-1)和反式橙花醇(23.49 μg·h-1);香水文心的主要成分为β-罗勒烯(31.84 μg·h-1)和1,8-桉叶素(16.34 μg·h-1),含量均随花的发育上升,盛花期含量最高(图2)。

注:不同小写字母表示花香释放量显著差异(P<0.05)。Note: Statistical significance of the difference in the amount of floral scent released is indicated by different lowercase letters (P<0.05). The same as following.图1 不同品种文心兰不同花发育时期花香挥发物总相对含量Fig.1 Total floral volatile relative contents during different flower development stages in different varieties of Oncidium orchid
2.2 转录组数据分析
从9个文库中共组装获得459 756个unigenes,平均长度为475 bp(表4)。在这459 756个unigenes中,186 711个被七大公共数据库注释,占总数的40.61%。为了鉴定unigenes分属的代谢途径,进行KEGG注释,结果显示有42 157(9.17%)个unigenes分属于123个KEGG途径。图3标注的是差异基因最多的代谢途径,包括初级代谢和次级代谢。3个品种中共发现34 313个差异基因,白梦香和黄梦香的差异基因较少,共5 431个,与香水文心的差异基因较多,分别有17 098和24 312个(表5)。
2.3 MVA和MEP途径关键基因分析
文心兰的转录组中共鉴定出14个MEP途径基因,包括3个OnDXS、1个OnDXR、3个OnMCT、2个OnCMK、1个OnMDS、1个OnHDS、1个OnHDR和2个OnGPPS,这14个基因中,多数基因在黄梦香中的表达最高,在香水文心中的表达略低,在白梦香中的表达最低(图4-A)。
共鉴定出的8个MVA途径基因中,包括1个OnAACT、1个OnHMGS、1个OnHMGR、1个OnMK、1个OnPMK,1个OnMDC和2个OnFPPS,这8个基因中,多数基因在黄梦香中的表达最高,在香水文心中的表达略低,在白梦香中的表达最低(图4-B)。
2.4 萜类关键基因分析
本研究从转录组中筛选获得5个显著差异的TPS基因,聚类分析表明,OnTPS1(DN124402_c1_g1)、OnTPS3(DN124003_c0_g3)和OnTPS4(DN99721_c2_g1)聚为TPS-b亚家族,主要由单萜合成酶(mono-TPSs)组成,OnTPS2(DN136754_c1_g1)和OnTPS5(DN126828_c0_g1)是TPS-f亚家族的成员(图5)。OnTPS4在3个品种中均高表达,qRT-PCR结果显示,随着花的发育,3个品种的OnTPS4表达量均上升,且在盛花期达到最高,说明OnTPS4在3个品种中对香
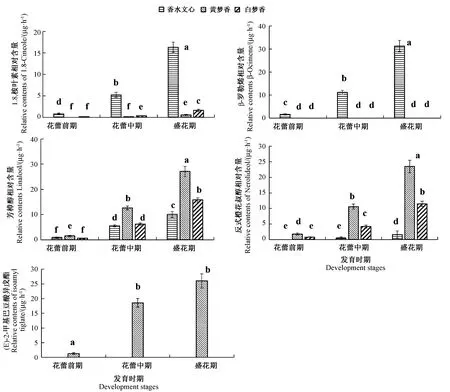
图2 文心兰3个品种主要花香挥发物相对含量Fig.2 Relative contents of main floral volatiles in three varieties of Oncidium

表3 不同文心兰品种盛花期主要花香成分Table 3 Main floral components of different varieties during blooming period in Oncidium orchid
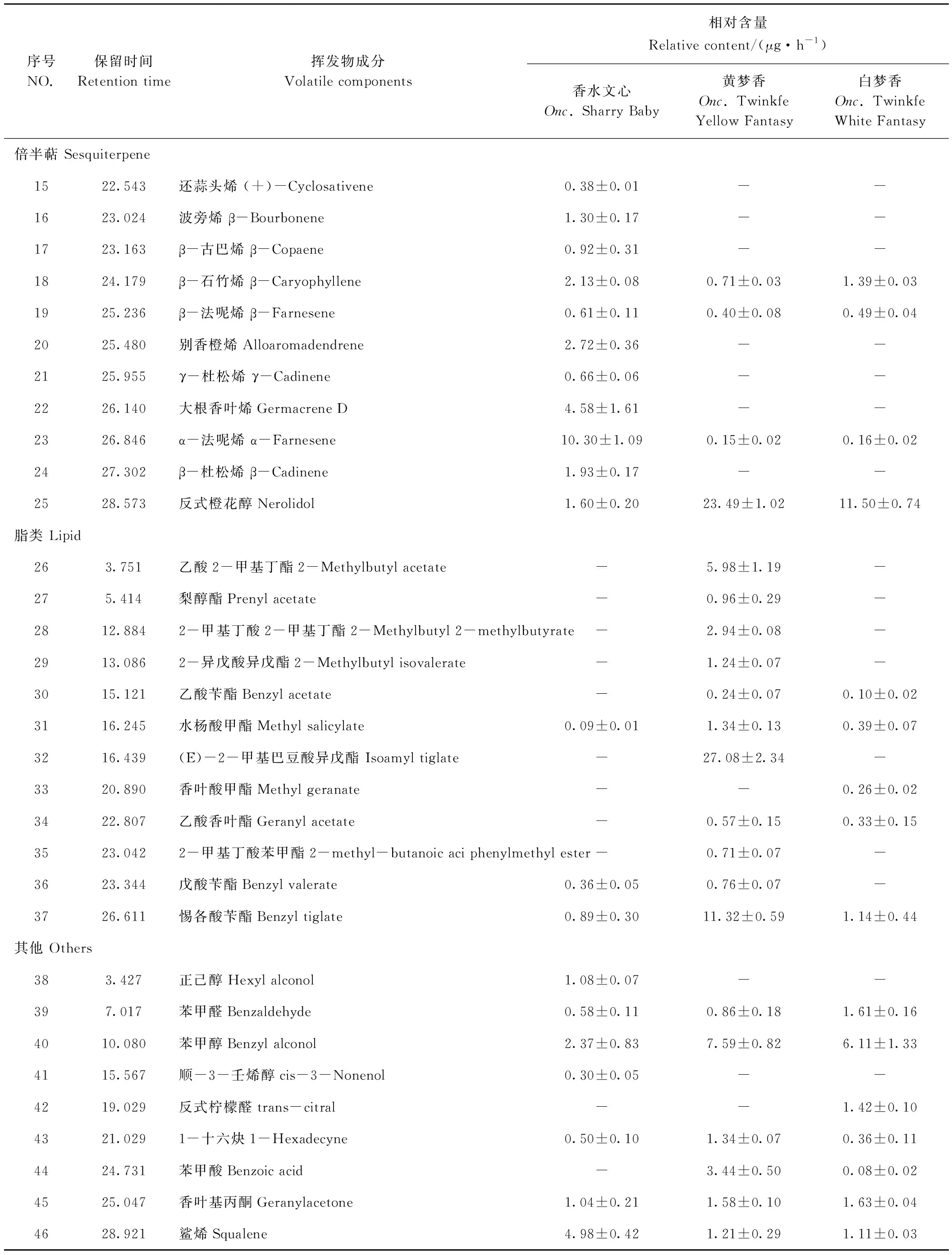
表3(续)

注:A: 细胞过程; B: 环境信息处理; C: 遗传信息处理; D: 代谢; E: 有机系统。Note: A: Cellular processes. B: Environmental information processing. C: Genetic information processing. D: Metabolism. E: Organismal systems.图3 香水文心、黄梦香和白梦香的KEGG注释Fig.3 KEGG pathway in Onc. Sharry Baby、Onc. Twinkfe Yellow Fantasy and Onc. Twinkfe White Fantasy
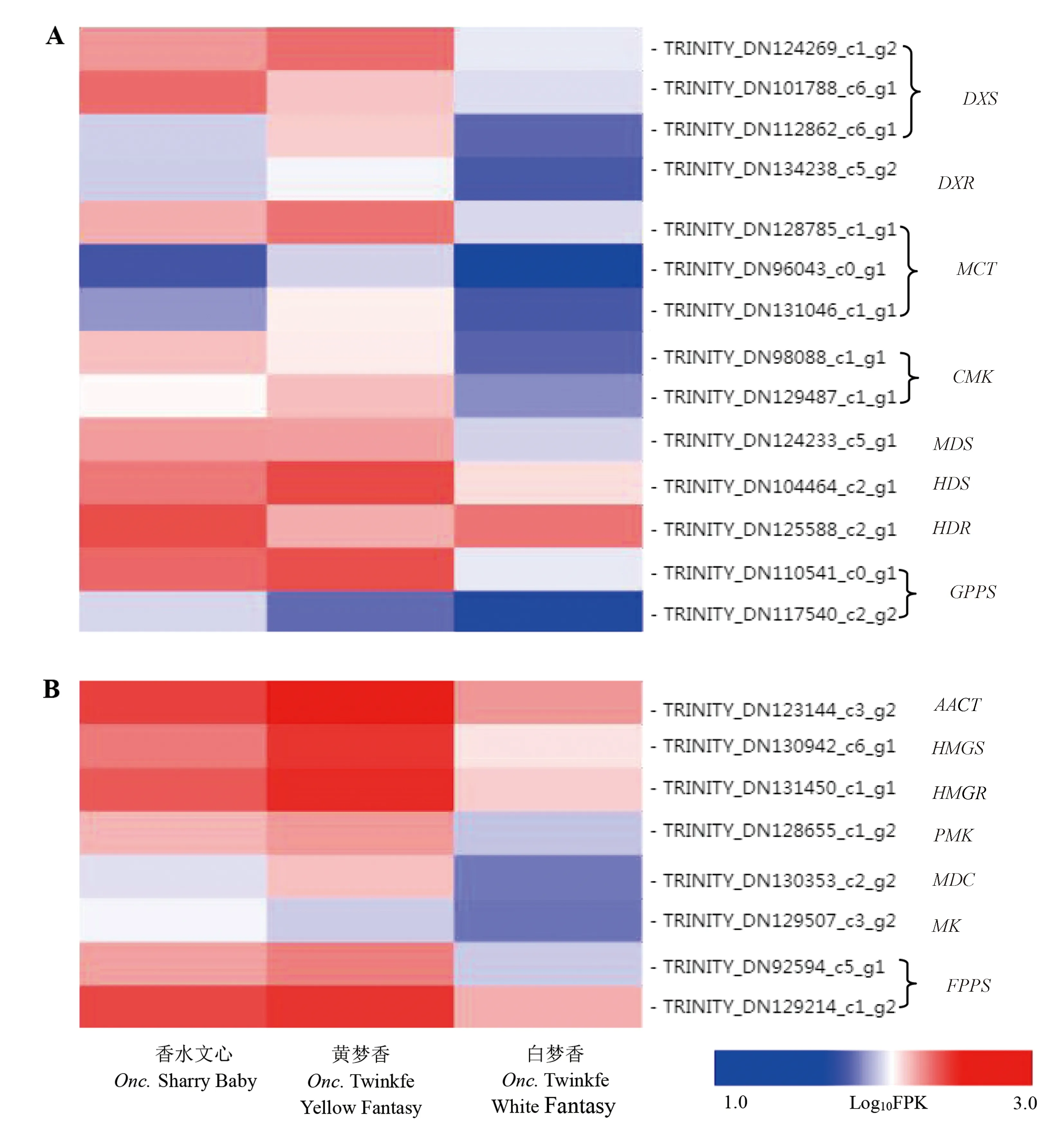
注:A:甲基赤藓醇磷酸途径;B: 甲羟戊酸途径。Note: A: 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway. B: Mevalonate(MVA) pathway.图4 单萜和倍半萜合成相关基因表达谱Fig.4 Expression profiles of the genes involved in the monoterpene and sesquiterpene biosynthesis

注:AaLIS: 黄蒿芳樟醇合酶(AAF13356); CrGES: 长春花香叶醇合酶(AFD64744); AaADS: 黄蒿紫穗槐-4,11-二烯合酶(AFA34434); AmMS: 金鱼草月桂烯合酶(AAO41727); AtCPS1: 拟南芥柯巴基焦磷酸合(NP_192187); CmCPS: 笋瓜柯巴基焦磷酸合酶(AAD04292); AgPS: 大冷杉蒎烯合酶(AAB71085); AgMS: 大冷杉月桂烯合酶(AAB71084); AgLS: 大冷杉柠檬烯合酶 (AAB70907); CbLIS: 仙女扇芳樟醇合酶(AAC49395); AtGA2: 拟南芥贝壳杉烯合酶(AAC39443)。Note: AaLIS: Artemisia annua linalool synthase (AAF13356). CrGES: Catharanthus roseus geraniol synthase (AFD64744). AaADS: Artemisia annua amorpha-4,11-dienesynthase (AFA34434). AmMS: Antirrhium majus myrcene synthase (AAO41727). AtCPS1: Arabidopsis thaliana copalyl diphosphate synthase (NP_192187). CmCPS: Cucurbita maxima copalyl diphosphate synthase (AAD04292). AgPS: Abies grandis pinene synthase (AAB71085). AgMS: Abies grandis myrcene synthase (AAB71084). AgLS: Abies grandis 4S-limonene synthase (AAB70907). CbLIS: Clarkia breweri S-linalool synthase (AAC49395). AtGA2: Arabidopsis thaliana ent-kaurene synthase (AAC39443).图5 文心兰TPS进化树分析Fig.5 Phylogenetic analysis of the TPS in Oncidium orchid

注:P1: 花蕾初期; P2: 花蕾中期; P3: 盛花期;F: TPS基因在3个文心兰品种盛花期的FPKM值;a: 香水文心; b:黄梦香; c:白梦香。Note: P1: Early bud period. P2: Mid bud period. P3: Blooming period. F: FPKM value of TPS gene in blooming period of three Oncidium cultivars. a: Onc. Sharry Baby. b: Onc. Twinkfe Yellow Fantasy. c: Onc. Twinkfe White Fantasy.图6 萜类合成酶基因表达分析Fig.6 Expression analysis of terpene synthase genes

表4 转录组测序和组装数据Table 4 Summary of sequencing and assembly data

表5 转录组差异表达基因Table 5 Differentially expressed genes of transcriptome sequencing
气的形成起重要作用。OnTPS1、OnTPS2和OnTPS5在香水文心中表达显著高于黄梦香和白梦香,qRT-PCR结果显示,随着花的发育,这3个基因在香水文心中均逐渐升高,在黄梦香和白梦香中随花的发育表达量均未上升,说明这3个基因在香水文心花香挥发物的形成中起重要作用。OnTPS3在黄梦香中表达量最高,香水文心次之,白梦香最低(图6)。
3 讨论
花香作为观赏植物品质的重要特征,其评价也是植物学研究的热点。花香的组成和释放模式已在许多植物中研究和明确,但其生物合成与调控机制尚未被揭示。本研究以香水文心、黄梦香和白梦香为材料,通过转录组测序探索花香形成的分子机制,共获得459 756个unigenes,40.61%的unigenes被公共数据库注释,这些信息为挖掘文心兰花香关键基因和功能提供了重要资源。
花香主要由萜类化合物和苯类/苯丙酸类化合物组成,该类物质由低分子量挥发性的复杂混合物组成。张莹等[21-22]对香水文心、蜜糖和飞鸟花香成分的研究中,同样发现萜类为主要挥发物,但酯类含量很少。单萜类化合物在多种植物中为花香的主要成分,如小苍兰(Freesiahybrida)[29]、百合(Lilium)[30]、姜花(Hedychiumcoronarium)[31]等。本研究GC-MS结果表明,香水文心和梦香系列的花香挥发物均以单萜为主,但种类差异较大,这与前人研究结果基本一致。本研究中,黄梦香酯类含量最高,达53 μg·h-1。与挥发物种类相近的白梦香相比,黄梦香香气更加浓郁,这可能与其主要香气成分的嗅觉阈值偏低及香气成分总含量高有关,酯类含量高可能是香气更浓郁的另一因素。梦香系列与香水文心的花香成分差异较大,可能是不同萜类合成酶基因表达差异的结果。
MEP和MVA途径分别为单萜和倍半萜的上游途径[32-33]。在百合中,MEP途径中的多数基因在有香气的Siberia中表达量显著高于无香气的Novano[32]。本研究中,MEP和MVA途径中的多数基因在黄梦香中的表达最高,在香水文心中表达比黄梦香中略低,在白梦香中表达最低,花香挥发物总量在黄梦香中最高,白梦香中最低,这两者是相吻合的,说明黄梦香在MEP途径的激活水平高于香水文心和白梦香。TPSs通过一步反应将GPP或FPP转化为各种萜类化合物,植物中含有多样的TPS。TPS通常被分为7个进化枝,而在一些植物中TPS仅有2个或3个进化枝,揭示了特定基因类型的扩增[34]。文心兰转录组数据中发现了5个显著差异表达的TPS,3个属于TPS-b, 主要由单萜合成酶(mono-TPSs)组成,2个属于TPS-f。前人研究结果表明,萜类合成酶(TPS)的高表达是花香挥发物释放量高的主要原因[34-35]。在花发育的过程中,金鱼草挥发性萜类的释放(如β-罗勒烯和月桂烯)与TPS基因的表达密切相关[15]。OnTPS4 在3个品种中均高表达,表明该基因在3个品种单萜合成中起关键作用;芳樟醇在3个品种中含量均较高,且均随着花的发育含量上升,OnTPS4的表达与芳樟醇的释放规律接近。OnTPS1在香水文心中表达偏高,在黄梦香和白梦香中低表达,并随着花的发育表达量上升,说明OnTPS1是香水文心单萜挥发物形成的重要基因;β-罗勒烯和1,8-桉叶素在香水文心中含量高,在黄梦香和白梦香中含量很低,且释放规律与OnTPS1的表达接近,推测OnTPS1可能是β-罗勒烯合成酶或1,8-桉叶素合成酶。OnTPS3在在黄梦香中表达偏高,说明OnTPS3是香水文心单萜挥发物形成的重要基因。OnTPS2和OnTPS5 在香水文心中表达偏高,对香水文心萜类挥发物的形成起到重要作用,但确定上述2个基因的具体类型和分布,仍需要进行亚细胞定位或体外酶活性测定。
4 结论
本研究确定了3种文心兰的主要花香成分为萜类化合物,并从转录组数据中共获得459 756个单基因(unigenes),从上述单基因中筛选出5个显著差异表达的TPS基因(OnTPS1~5),验证了上述5个基因均对文心兰花香挥发物的释放起重要作用。在此基础上,后续研究可对5个TPS基因进行酶活性测定,以确定具体基因类型,并通过遗传转化,对无香文心兰进行改良。
猜你喜欢
中草药(2022年21期)2022-11-05 07:08:14
中国农业科学(2021年1期)2021-01-14 03:04:10
核农学报(2020年4期)2020-03-13 14:19:58
中成药(2018年11期)2018-11-24 02:57:04
天然产物研究与开发(2018年7期)2018-08-21 02:04:02
石油炼制与化工(2018年6期)2018-03-27 03:18:40
学苑创造·A版(2018年12期)2018-03-04 06:57:06
时代人物(新教育家)(2017年10期)2017-12-18 06:42:48
中国篆刻(2017年5期)2017-07-18 11:09:30
学苑创造·A版(2017年6期)2017-06-23 17:57:12
