
贝那鲁肽对高糖诱导的胰岛β细胞功能障碍和凋亡的影响及机制研究
2022-03-08 05:42辛丽娟宁建峰唐宏霞左源渊王新婷尚宏博鲍喜静张家口市第一医院内分泌二科检验科内分泌代谢病门诊内分泌一科河北张家口075000
现代检验医学杂志 2022年1期
辛丽娟,宁建峰,唐宏霞,左源渊,王新婷,彭 一,尚宏博,鲍喜静(张家口市第一医院.内分泌二科;.检验科;.内分泌代谢病门诊;.内分泌一科,河北张家口,075000)
近年来糖尿病的发病率迅速增加[1]。糖尿病的高死亡率和心血管并发症仍然是目前卫生保健系统面临的难题,严重影响患者的生活质量,增加社会负担[2]。研究表明,胰岛β细胞的丢失、破坏或功能障碍可导致糖尿病和高血糖,而促进胰岛β细胞的再生和减少胰岛β细胞的凋亡是近年来治疗糖尿病的研究热点[3-4]。贝那鲁肽是全球首个人源氨基酸序列的GLP-1 药物,广泛用于治疗2型糖尿病。研究表明GLP-1 可刺激胰岛β细胞复制、再生和分化,并通过减少应激来抑制胰岛β细胞的凋亡[7],但具体机制尚不完全清楚。本研究旨在探究贝那鲁肽减轻高糖诱导的胰岛β细胞系(INS-1)功能障碍和凋亡的分子机制,扩展对贝那鲁肽抗凋亡作用的理解,并进一步支持了GLP-1 类似物在2型糖尿病患者治疗中的应用。
1 材料与方法
1.1 研究对象 大鼠胰岛素瘤细胞系INS-1 购自上海抚生实业有限公司。将INS-1 胰岛β细胞培养于含有10ml/dl 胎牛血清,100U/ml 青霉素和100μg/ml 链霉素的RPMI-1640 中,置于37℃含有5ml/dl CO2的细胞培养箱中,每2 天换一次液。
1.2 仪器与试剂 电泳仪与转膜仪(美国,伯乐);曝光仪(美国,UVP),酶标仪及细胞培养箱(赛默飞世尔科技有限公司,美国)。贝那鲁肽购自上海仁会生物制药股份有限公司,RPMI-1640 培养液、胎牛血清、96 孔板和24 孔细胞培养皿购自Gbico公司;青链霉素混合液购自上海碧云天生物技术有限公司;MG-132,ISO-1,二甲基亚砜和噻唑蓝(MTT)试剂盒购自美国Sigma-Aldrich 公司;大鼠/小鼠胰岛素酶联免疫吸附测定(ELISA)试剂盒购自美国LINCO research 公司;巨噬细胞迁移抑制因子(MIF)ELISA 试剂盒购自Abcam 公司;鼠抗鼠Bax和IκB抗体 、兔抗鼠cleaved caspase3抗体、HRP 标记山羊抗鼠 lgG( H + L),HRP 标记山羊抗兔 lgG( H + L)购自美国 Cell Signaling Technology公司;兔抗鼠p-IκB,NF-κB p65 和β-actin 购自Abclonal 公司;蛋白质 Marker 购自 Fermentas 公司;总蛋白提取试剂盒购自南京凯基公司;磷酸盐缓冲液(PBS)、Tris 缓冲盐溶液(TBS)购自武汉博士德公司;BCA 蛋白浓度测定试剂盒购自美国Pierce 公司;甲醇购自国药集团化学试剂有限公司;Tween-20 购自北京普利莱基因技术有限公司;脱脂奶粉购自伊利乳业股份有限公司;5×上样缓冲液、SDS-PAGE 凝胶配制试剂盒、RIPA 细胞裂解液、免疫印迹化学发光试剂(ECL),抗荧光淬灭封片剂购自碧云天生物技术有限公司。
1.3 方法
1.3.1 分组:对照组(Control group):向培养液中加入5.5mmol/L 葡萄糖,培养24h。高糖组(HG组):向培养液中加入30mmol/L 葡萄糖,培养24h。贝那鲁肽处理组(HG+BL 组):在高糖组的基础上,向培养液中加入1nmol/L 贝那鲁肽,培养24h。MG-132 预处理组:在高糖组的基础上,提前1h 给予20 μmol/L MG-132 预处理,培养24h。ISO-1 预处理组:在高糖组的基础上,提前1h 给予50 μmol/L ISO-1 预处理,培养24h。
1.3.2 INS-1 细胞活力测定:采用比色MTT 测定法评估细胞活力,将细胞(2×104细胞/孔)接种在96孔板中,设置对照组、高糖组和贝那鲁肽处理组,孵育48h,随后将MTT 溶液(100 µl,1 mg / ml)添加到96 孔板的每个孔中,并在37℃下孵育4h 至有紫蓝色结晶析出,离心弃上清。加入100 μl 二甲基亚砜震荡。使用酶标仪在540 nm 处测定每个孔的内容物的吸光度。根据公式计算细胞存活率。细胞存活率(%)=(样本孔A540nm/对照孔A540nm)×100%。
1.3.3 葡萄糖刺激胰岛素分泌(GSIS)实验:将细胞(2×104个/孔)接种在24 孔板中,设置对照组、高糖组、贝那鲁肽处理组、MG-132 预处理组和ISO-1预处理组,在无葡萄糖Krebs-Ringer缓冲液(129 mmol/L NaCl,4.8 mmol/L KCl,2.5 mmol/L CaCl2,1.2 mmol/L MgSO4,1.2 mmol/L KH2PO4,5 mmol/L NaHCO3)中孵育1h 后,小心除去培养液,PBS 洗涤细胞,在37℃下用含有5 或25mmol/L 葡萄糖的Krebs-Ringer 缓冲液刺激1h。然后收集细胞培养液分析胰岛素的分泌。使用大鼠/小鼠胰岛素酶联免疫吸附测定试剂盒测定胰岛素分泌。
1.3.4 Western blot:收集各组INS-1 胰岛β细胞,加入适量RIPA 裂解液,冰上研磨裂解后提取总蛋白,使用BCA 蛋白浓度测定试剂盒检测蛋白浓度,加入上样缓冲液,沸水煮5min 使蛋白变性,取30 μg 蛋白电泳分离蛋白,而后将蛋白转移至PVDF 膜上,5ml/dl 脱脂奶粉室温封闭1h,加入Bax(1:500)、cleaved caspase3(1:1 000)、p-IκB(1:500)、IκB(1:1 000)、NF-κB p65(1:300)及β-actin(1:1 000)一抗,4℃过夜。TBST 洗膜3 次,37℃二抗孵育1.5h,TBST 洗膜3 次,使用 UVP 凝胶成像系统,ECL 化学发光显色,采用 Image J 图像分析系统软件分析条带光密度值。
1.4 统计学分析 统计学处理采用 GraphPad Prism 5.0 统计软件,数据采用均数±标准差表示,组间比较采用t检验。P<0.05 为差异有统计学意义。
2 结果
2.1 贝那鲁肽对高糖诱导的INS-1 胰岛β细胞活力和胰岛素分泌的影响 见表1。与对照组相比,高糖组INS-1 胰岛β细胞的存活率和胰岛素的分泌均显著降低(P<0.05);与高糖组相比,贝那鲁肽处理组INS-1 胰岛β细胞的存活率和胰岛素的分泌均可显著增加(P<0.05)。
表1 贝那鲁肽对高糖诱导的INS-1 胰岛β细胞活力和胰岛素分泌的影响(±s)

表1 贝那鲁肽对高糖诱导的INS-1 胰岛β细胞活力和胰岛素分泌的影响(±s)
项目对照组HG 组HG+BL 组FP细胞存活率(%)97.28±4.549.49±9.60*75.08±4.83#12.63<0.001 5mmol/L 葡萄糖(ng/3×105cells/hr)11.27±1.545.53±0.95*14.82±2.12#8.484<0.05 25mmol/L 葡萄糖(ng/3×105cells/hr)18.01±1.4210.00±1.23*20.63±2.15#11.30<0.01
2.2 贝那鲁肽对高糖诱导的INS-1 胰岛β细胞凋亡的影响 见图1。Western blot 检测结果发现,与对照组相比,高糖组INS-1 胰岛β细胞Bax 蛋白和cleaved caspase 3 表达明显增加(t=2.923,P<0.05;t=3.141,P<0.05);与高糖组相比,贝那鲁肽处理组Bax 蛋白和cleaved caspase 3的表达显著降低(t=2.500,2.558,均P<0.05),差异均有统计学意义。

图1 Western blot 检测贝那鲁肽对高糖诱导的INS-1 胰岛β细胞Bax 蛋白和cleaved caspase 3 表达的影响
2.3 贝那鲁肽对高糖诱导的INS-1 胰岛β细胞内NF-κB 信号通路相关蛋白表达的影响以及MIF 分泌的影响 见图2。与对照组相比,高糖组MIF 分泌明显增加(t=3.024,P<0.05);与高糖组相比,贝那鲁肽处理组MIF 分泌明显降低(t=2.415,P<0.05),差异均有统计学意义。Western blot 检测结果显示,与对照组相比,高糖组可增加磷酸化IκB(p-IκB)和NF-κB p65的表达(t=3.544,4.658,均P<0.01),减少IκB的表达(t=3.062,P<0.05);与高糖组相比,贝那鲁肽可抑制p-IκB 和NF-κB p65的表达(t=2.959,3.047,均P<0.05),促进IκB的表达(t=3.453,P<0.05)。
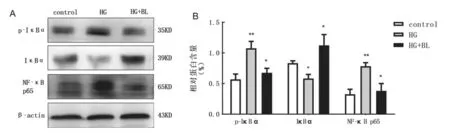
图2 Western blot 检测贝那鲁肽对高糖诱导的INS-1 胰岛β细胞p-IκB,IκB 和NF-κB p65 表达的影响
2.4 高糖诱导NF-κB/MIF 途径激活对INS-1 胰岛β细胞凋亡和功能障碍的影响 见图3。Western blot 结果表明,与高糖组相比,MG-132 和ISO-1预处理均可显著降低Bax(t=4.022,2.554,均P<0.05)和cleaved caspase 3(t=4.022,2.760,均P<0.05)的表达,差异均有统计学意义。
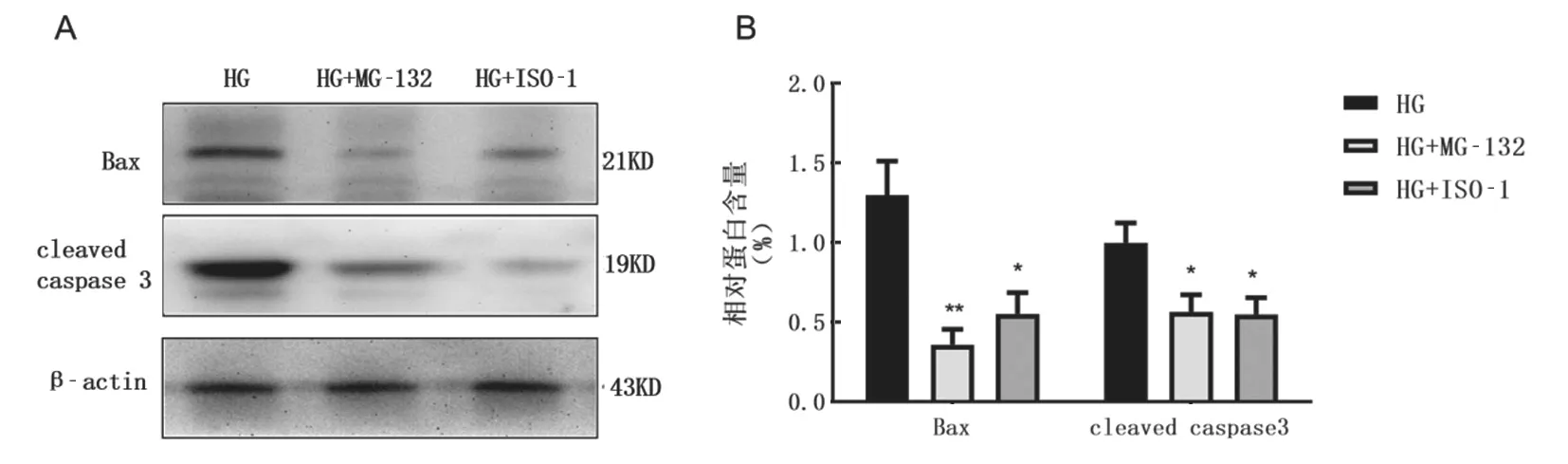
图3 Western blot 检测MG-132 和ISO-1 预处理对INS-1 胰岛β细胞Bax 蛋白和cleaved caspase 3 表达的影响
2.5 MG-132 和ISO-1 预处理组对高糖诱导的INS-1 胰岛β细胞胰岛素分泌的影响 见表2。与高糖组相比,MG-132 和ISO-1 预处理均可促进胰岛素的分泌(P<0.05)。

表2 MG-132 和ISO-1 预处理对高糖诱导的INS-1 胰岛β细胞胰岛素分泌的影响
3 讨论
慢性高血糖症是糖尿病的标志,它具有β 细胞毒性,可加剧β 细胞衰竭,形成恶性循环[8]。研究表明,高糖可增加人类胰岛细胞分泌IL-1β,导致NF-κB 转录因子活化、FasL 上调和胰岛β细胞功能障碍[9];高糖还可通过NF-κB 依赖性方式诱导软骨细胞的分解代谢和炎症反应[10]。而过度炎症反应是诱导细胞凋亡和功能障碍的重要因素。本研究发现了高糖不仅可抑制INS-1 胰岛β细胞的活性和胰岛素的分泌功能,还可增加INS-1 胰岛β细胞NF-κB 炎症通路相关蛋白的表达,而给予NF-κB 抑制剂MG-132 处理后,可降低凋亡相关蛋白的表达,还可改善胰岛β细胞胰岛素分泌功能,进一步说明了NF-κB 炎症通路在胰岛细胞凋亡和功能障碍中的重要调节作用。
MIF 即巨噬细胞迁移抑制因子,是一种包括胰岛β细胞在内的多种免疫细胞和非免疫细胞产生的促炎细胞因子,其分泌过多可以增加机体的炎症[11]。此外,MIF 还是NF-κB 下游促炎因子,可调节机体的固有免疫和适应性免疫。研究表明,MIF 可募集免疫细胞至炎症位点,进而增加促炎因子TNF-α,IL-1β,IL-17 和IFN-γ的表达[12-14]。相关临床研究显示,糖尿病患者血清MIF 明显升高,与糖耐量受损有关[15]。MIF 可在胰岛β细胞内实现适当的胰岛素构象,并发挥其全部活性,MIF 表达的固有或获得性变化可能导致不同的胰岛素加工过程和自身免疫的开始[16]。此外,MIF 还被证实通过增强炎性环境并使靶细胞无法响应胰岛素,从而间接地和负面地影响胰岛素的作用[16]。本研究发现高糖诱导的INS-1 胰岛β细胞培养上清液中MIF 明显增高,而给予MIF 抑制剂ISO-1 预处理后,可改善高糖诱导的胰岛β细胞活性和胰岛素分泌功能,降低凋亡蛋白Bax 和cleavedcaspase 3的表达,表明了MIF参与胰岛细胞凋亡和功能障碍。
贝那鲁肽是2型糖尿病治疗的主要药物,其主要活性成分rhGLP-1 具有控制血糖的作用。研究表明,GLP-1 受体激动剂可通过急性促胰岛素作用以及在某些情况下通过长期作用来改善血糖控制,从而通过刺激β 细胞增殖和抑制凋亡来保持β 细胞质量[17-18]。这与本研究结果贝那鲁肽干预可以改善胰岛β细胞的活性并减少其凋亡一致。此外,贝那鲁肽还具有抗炎作用。研究表明,GLP-1 受体激动剂exendin-4(50 nmol/L)处理培养的人胰岛细胞可抑制炎症基因NFκB1(p105)、 NFκB2(p100)以及TNF 受体超家族成员1A的表达[5]。exendin-4还可抑制ICAM-1 和MIF的表达,改善氧化型低密度脂蛋白对巨噬细胞迁移的抑制作用[19]。本研究发现贝那鲁肽可抑制高糖诱导的INS-1 胰岛β细胞MIF 的分泌和NF-κB 信号通路相关蛋白p-IκB和NF-κB p65的表达,促进IκB的表达,而NFκB/MIF 抗炎通路可调控高糖诱导的胰岛β细胞凋亡和功能障碍。因此,贝那鲁肽可能通过抑制NF-κB/MIF 依赖的炎症途径减轻高糖诱导的胰岛β细胞功能障碍和凋亡。
猜你喜欢
中国动物检疫(2022年10期)2022-10-14
法医学杂志(2022年1期)2022-06-21
安徽医药(2021年4期)2021-04-09
旗帜文摘(2020年5期)2020-06-09
科学大观园(2020年7期)2020-04-08
保健与生活(2020年6期)2020-03-20
中国卫生产业(2018年12期)2018-05-14
中国新闻周刊(2016年32期)2016-10-27
现代园艺(2016年17期)2016-10-17
家庭科学·新健康(2016年6期)2016-06-30

- 现代检验医学杂志的其它文章
- 2型糖尿病肾病患者外周血micorRNA-155水平与肾小球滤过率的相关性研究
- 子痫前期并发幽门螺旋杆菌感染患者血清miR-31-5p表达与不良妊娠结局的相关性研究
- 子宫肌瘤患者血清和组织中hsa-miR-15b-5p,hsa-miR-21-5p的水平与组织中ER,PR蛋白表达的相关性研究
- 妊娠糖尿病患者血清、脐血及胎盘组织中Visfatin和Chemerin的表达与胰岛素抵抗发生机制的研究
- 完全性川崎病患儿血清25-羟基维生素D水平检测对预测IVIG抵抗的价值研究
- 子宫内膜异位症患者血清中PAPP-A,IGF-I水平及PAPP-A基因rs7020782 A/C位点多态性与临床分期相关性研究