
Conifer establishment after the eruption of the Paricutin volcano in central Mexico
2022-03-08 02:18JesEdurdoenzCejBlnLizethenzCejTriniddenzReyesDiegoRfelrezSlirup
Forest Ecosystems 2022年1期
Jesús Edurdo Sáenz-Cej ,Bln Lizeth Sáenz-Cej ,J.Trinidd Sáenz-Reyes ,Diego Rfel Pérez-Slirup,*
aInstituto de Investigaciones en Ecosistemas y Sustentabilidad,Universidad Nacional Aut'onoma de M'exico,Antigua Carretera a P'atzcuaro 8701 Col.Ex hacienda San Jos'e de la Huerta,58190,Morelia,Mexico
b Escuela Nacional de Estudios Superiores Unidad Morelia,Universidad Nacional Aut'onoma de M'exico,Antigua Carretera a P'atzcuaro 8701 Col.Ex hacienda San Jos'e de la Huerta,58190,Morelia,Mexico
cInstituto Nacional de Investigaciones Forestales,Agrícolas y Pecuarias,Campo Experimental Uruapan,Av.Latinoamericana 1101,Col.Revoluci'on,60500,Uruapan,Mexico
Keywords:Age structure Dendrochronology Lava flow Primary succession Tropical montane forest
ABSTRACT Background: Volcanic eruptions have large effects on forest ecosystems and create new substrates,triggering primary succession processes.The Paricutín volcano,born in central-western Mexico,erupted between 1943 and 1952.After the cessation of the eruptive activity,plant colonization began on the lava flows and tephra deposits,including the conifer species that dominate the surrounding mature forests.This study aims to reconstruct the history of the establishment of conifer trees on the substrates created by the Paricutín eruption.Methods: 16 sampling plots were established along three transects with northern,southern,and south-western aspects,located every 250 m from the preserved forest to the volcanic cone.Increment cores from 400 conifer trees were extracted and their age was determined by cross-dating annual tree rings.The order of the species colonization and the tree establishment,abundance,and dominance patterns were characterized.Also,the influence of the distance from the mature forests and the inter-annual climatic conditions on the temporal tree establishment pattern was evaluated.Results:Eight pine and one fir species have been established since 1970,only 18 years after the end of the eruptive period.However,tree establishment increased by 12.9% annually after 1995,with the youngest tree in our sample getting established in 2015.We did not find a well-defined temporal and spatial pattern of species arrival,which suggests that colonization occurred randomly,although the four pine species that were established early became the most abundant and dominant.Tree establishment was not influenced by the distance from the mature forest,and wet inter-annual conditions did not enhance pulses of tree recruitment,exhibiting a continuous tree establishment pattern.Conclusions: Conifer species have shown a great capacity for colonizing volcanic substrates created by the Paricutín eruption,which suggests that tropical montane conifers can regenerate rapidly under high-magnitude disturbances.These findings support the use of these forest species for ecological restoration.
1.Background
Volcanic eruptions are large disturbances that substantially modify environmental conditions,resource availability,substrates,and the structure of ecosystems and landscapes (Del Moral and Grishin,1999).The deposition of ash and volcanic sand (tephra),including subsequent lava flows,can cover large areas,which represent a barrier for plant establishment (Titus and Tsuyuzaki,2003) since these new substrates experience high levels of insolation,extreme temperature fluctuations,low organic matter content,low water retention,and high porosity(Chang et al.,2019).
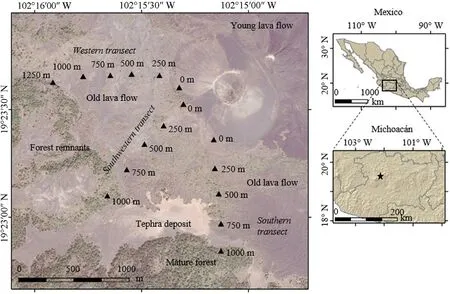
Fig.1.Location of the Paricutín volcano in Mexico and sampling plots to reconstruct the age of conifer establishment after the eruptive period.

Fig.2.(a) View of the Paricutín volcano and vegetation near the mature forest;(b) state of the vegetation in the rocky ground at an intermediate elevation;(c)vegetation near the top of the volcano;(d) extraction of the increment cores.
The time from eruption to the arrival of vegetation is known as the establishment span or ecesis (Pierson,2007).The most common successional pattern in volcanic areas is the colonization by pioneer species such as lichens and mosses,followed by grasses and shrubs,and then eventually by tree species(Cutler et al.,2008).Plant succession depends strongly on the ability of seeds to germinate,establish and grow in the new substrates (Vilmundardóttir et al.,2018).In addition,the distance from mature forests to the gaps covered by volcanic substrates influences the arrival of colonizing vegetation(Holle and Tsuyuzaki,2018).Climate variability also plays a fundamental role in plant establishment since moisture availability and suitable temperatures influence tree recruitment(Del Moral and Grishin,1999).
The substrates formed by volcanoes represent an opportunity to characterize the trajectories of primary succession,the establishment span,the order of species arrival,the ability of plants to survive under extreme environments,the effect of distance from the mature forest at the time of establishment,and spatial patterns of colonization(Del Moral and Bliss,1993).This information is crucial in providing guidelines to manage disturbed areas and inform ecological restoration efforts,even by other disturbances such as large fires (Robinson and Handel,2000).In temperate ecosystems,the formation of annual growth rings in conifer species offers the possibility to reconstruct the history of population establishment and evaluate the capacity of these species to regenerate after this kind of disturbances(Stoffel and Bollschweiler,2008).
The Paricutín volcano emerged in 1943 in western-central Mexico and caused the devastation of towns,agricultural lands,and conifer forests,including the fall of volcanic ash over 300 km2(Pioli et al.,2008).The eruption also affected the growth of pine trees in surrounding forests(Sheppard et al.,2008).In areas covered by petrified magma and tephra(24.5 km2),primary succession began rapidly in 1950 in the oldest substrates,mainly by mosses and lichens;vascular plants arrived in 1957,reaching up to 28 species among herbs and shrubs in 1960 (Eggler,1963).In 1977,25 years after the end of the eruption,39 vascular plants were recorded(Réjmanek et al.,1982).Later,50 years after the eruption,49 plant species had colonized the new substrates(Lindig-Cisneros et al.,2006).
Field-based observations have reported the presence of conifer trees established on the tephra deposits and lava flows of the Paricutín volcano,which are the dominant species in the forest surrounding the volcanic substrates (Medina,2003).However,the history of the establishment of these conifer trees has not been documented.Since trees represent the latest seral stage in high-magnitude disturbances(Cook and Halpern,2018),this information could be useful to characterize the successional pattern of dominant trees in these volcanic substrates.
The aims of this study were:(1) to reconstruct the history of the establishment of conifer trees in tephra and lava flows from the Paricutín volcano,(2)to characterize the order of the species colonization and the tree establishment,abundance,and dominance patterns,(3)to evaluate the effect of the distance from the preserved forest on the temporal and spatial establishment patterns,and(4)to evaluate the effect of climate on tree establishment.
2.Methods
2.1.Study area
The Paricutín volcano is located at coordinates 19°29′39′′N,102°15′05′′W in the state of Michoacán,in central Mexico,belonging to the Tran-Mexican Volcanic Belt (Fig.1).Its volcanic activity began on February 20,1943,and ended on March 4,1952,emerging on a montane plain covered originally by cornfields at 2,400 m of elevation (Wilcox,1954).It was the second volcano whose emersion was historically recorded in that portion of Mexico,after the El Jorullo volcano,which emerged in 1759,100 km southeast of Paricutín(Bullard,1947).
The Paricutín volcano is a monogenetic cineritic cone,formed by a Strombolian eruption which reached a height of 424 m,at a maximum altitude of 2,782 m (Fig.2a),expulsing pyroclastic bombs,ash,sand(tephra),incandescent material,and magma during three stages:the formation of the cone,the opening of the adventitious cone Sapichu,and the release of lava flows(Foshag and González,1956;Pioli et al.,2008).It spewed a total of 1.4 million m3of lava over 24.5 km2(2,450 ha)(Wilcox,1954).
Due to the geomorphological characteristics of the terrain where the Paricutín volcano emerged,the first lava flows were concentrated on the southern and southwestern cone faces,while the later lava flows advanced down the slope towards the northern and eastern cone faces,destroying the towns of Paricutín and San Juan Parangaricutiro (Krauskopf,1948).The area covered by tephra was 233 km2(23,300 ha),with an average depth of 25 cm,while on the slopes and in the valleys,it reached up to 3 m (Inbar et al.,1994).
The climate is humid subtropical with summer rains,an average annual temperature of 15°C,and average annual rainfall of 1,500 mm(Medina-García et al.,2000).The soils of the surrounding forests are Andosols,which were covered by a layer of tephra (Siebe et al.,2003).The dominant vegetation is pine-oak forest,with a high degree of human disturbance (Fregoso et al.,2003).A floristic richness of 108 families,306 genera,and 716 species was recorded in the mature forests,of which 12 are pine species and two fir species,including the endangeredPinus martineziiLarsen (ocote pine) andAbiesflinckiiRushforth (Jalisco fir)(Medina,2003).Currently,the volcanic cone and the land south of the lava flows belong to the Caltzontzin indigenous community,ofPur'epechaorigin,which has owned these lands since 1599 by a royal decree during the colonial period,currently managing a surface of approximately 6,700 ha(Corona-Chávez,2018).

Fig.3.Age structure of the sampled plots according to the transect(southern,southwestern,and western)and the distance to the Paricutín volcano(0,250,500,750,1000,and 1250 m).
2.2.Dendrochronological sampling
By analyzing a remote-sensing image from Google Earth Pro version 7.3.4.8248 (Google,2019),16 sampling plots were established in April 2019 along three transects (western,southwestern,and southern) with sampling plots every 250 m from the forest fragments to the volcanic cone.The western transect had a length of 1,250 m(6 plots),1,000 m in the southwestern(5 plots),and 1,000 m in the southern transect(5 plots)(Fig.1).In each sampling plot,the presence of conifers was verified and the elevation,slope-aspect,and slope of the terrain,substrate condition(sandy or rocky),and the density of coniferous trees in a circular area of 100 m2were recorded.
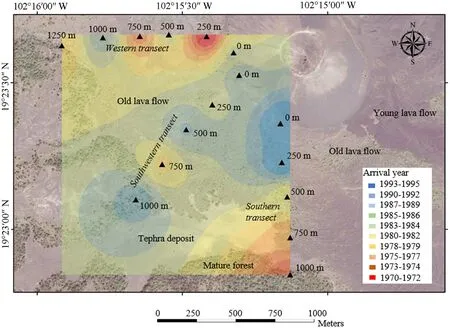
Fig.4.Year of the establishment of conifer trees after the Paricutín eruption between 1943 and 1952.
The elevation of the plots ranged from 2,554 to 2,672 m,starting from the edge of mature forest fragments (Fig.2a),passing through petrified magma and sandy lands(Fig.2b)until the highest point with the presence of conifers at the base of the volcanic cone (Fig.2c).Seven plots had a zenith position,while four of them were North-facing,three were Eastfacing,and two were South-facing.The slope of the plots ranged from 0°(flat surface)to 45°(steep slopes).Nine plots had a rocky substrate,four had a sandy substrate;the rest had a mixed substrate.The density of coniferous trees varied between 6 and 28 trees•(100 m-2),with an average density of16.7trees•(100m-2)(Table1).Noevidenceofpastfires,ortrees affected by pests was detected.No broadleaf tree species were recorded.
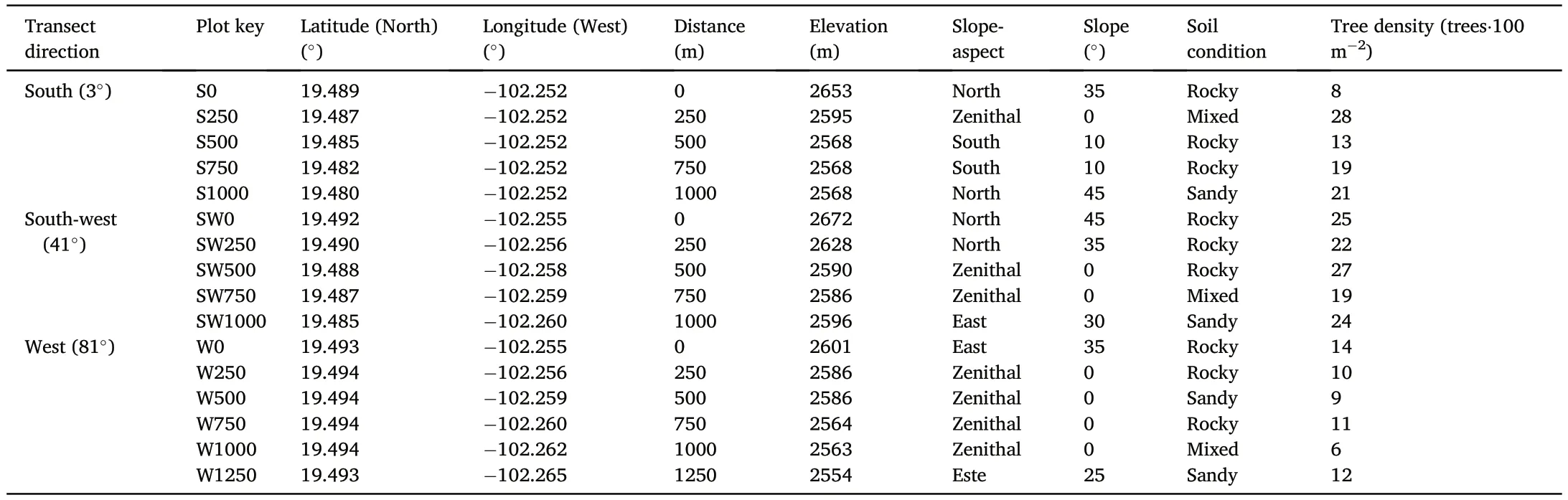
Table 1 Biophysical characteristics of the sampling plots around the Paricutín volcano,Mexico.The parenthesis indicates the azimuth of each transect.
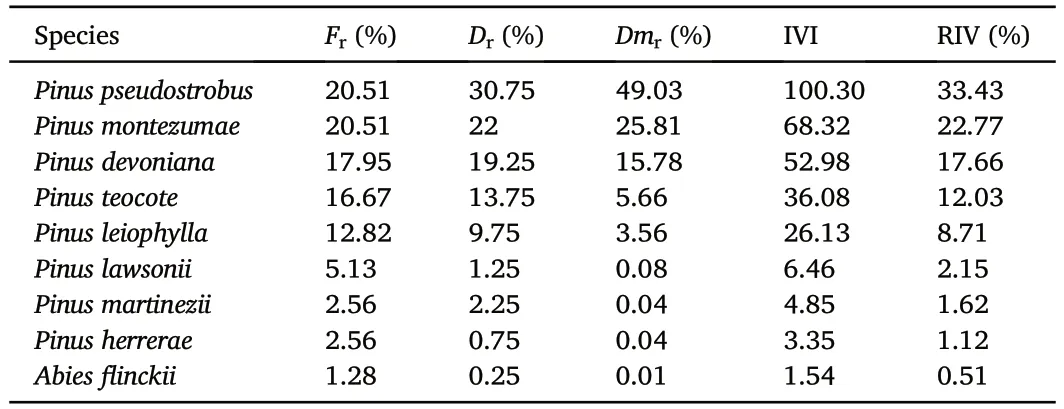
Table 2 Importance values of the coniferous species present around the Paricutín volcano,Mexico.
In each sampling plot,the closest 25 conifer trees with a basal diameter ≥of 5 cm were selected.The species were identified and their basal diameter was measured (cm).Then,with a Pressler increment borer,1 to 2 radial cores were extracted per tree,depending on whether the pith was reached in the first or second attempt.Following Speer(2010),radial cores were collected from the stem base,as close as possible to the ground,to obtain the tree age (Fig.2d).In total,radial cores from 400 individuals were sampled.
2.3.Growth ring cross-dating
In the laboratory,the increment cores were dried,glued to wooden mounts,and gradually polished with coarse to fine-grit sandpaper.The age of establishment was obtained by cross-dating the growth rings under a microscope (Stokes and Smiley,1996).Skeleton plots were used to compare the sequence of narrow rings with a regional master chronology obtained from an area 10 km south of the Paricutín volcano of a population of trees fromAbies religiosa(Kunth) Schltdl.et Cham (sacred fir)(Cerano-Paredes et al.,2014).This procedure allowed the identification of micro-rings,false rings,and missing rings(Speer,2010).
2.4.Composition of forest community
The importance value index (IVI) for all the sampled species was estimated,which indicates the importance of a species within a stand,obtained from the sum of relative frequency(1),relative density(2),and relative dominance (3) of the species according to the basal area (4)(Curtis and McIntosh,1951).The IVI(5)of each species was calculated as follows:

where
Fr=relative frequency
m=number of units where a species is found
MT=total number of units sampled
Dr=relative density
n=total number of individuals sampled
NT=total number of individuals sampled from all species
Dmr=relative dominance
AB=total basal area
ABT=total basal area of all species sampled
d=diameter at the base of trees

Fig.5.Relationship between conifer age and (a) distance to the volcano,(b) elevation,(c) slope,(d) slope aspect,(e) soil condition,(f) species,and (g) basal diameter.Relationship between distance to the volcano and (h) species,and (i) tree density.The key species of panel (f) are:dev= Pinus devoniana,her= Pinus herrerae,law=Pinus lawsonii,lei=Pinus leiophylla,mar=Pinus martinezii,mon=Pinus montezumae,pse=Pinus pseudostrobus,teo=Pinus teocote.Abies flinckii was not graphed since only one tree was sampled.
Once the importance value of each species was obtained,it was standardized to know its relative importance value (RIV),according to the following formula:

where
RIV=relative importance value
VIT=sum of the importance value index of all species
2.5.Age structure
The years of tree establishment were grouped and charted according to the sampling plot and the distance to the volcano.Pulses of regeneration were identified when four or more trees established in the same year,followed and preceded by 5-year periods without recruitment.A continuous establishment pattern was defined when there were no periods without recruitment.
The influence of distance to the volcano,elevation,transect,slope,substrate,plot tree density,tree basal diameter,and the species of the aged trees was evaluated,as well as the relationship between the distance to the volcano,the number of species and the tree density.First,the normality of distribution data was assessed through Shapiro-Wilk tests,in which tree age(W=0.94274,p>0.05),elevation(W=0.89602,p>0.05),and tree density (W=0.93614,p>0.05) followed a normal distribution,hence these variables were analyzed through linear regressions(lmfunction;stats package),analysis of variance(aovfunction;stats package) and analysis of covariance (aovfunction),with a confidence level of 95%.Since slope (W=0.79246,p<0.05) and basal diameter (W=0.90984,p<0.05) were not distributed normally,we used the non-parametricmagfunction (mcvg package) to analyze their relationship with tree age.These tests were run in the R program version 3.4.3(R-project Core Team,2017).
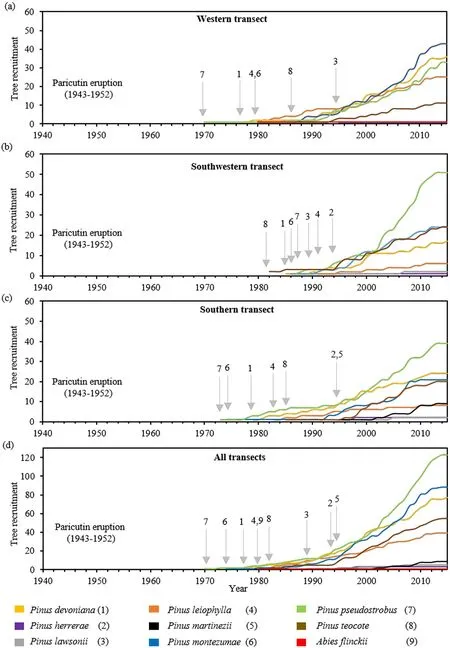
Fig.6.Conifer abundance after the eruption of Paricutín volcano in the(a)western,(b)south-western,(c)southern,and(d)all transects.The arrows indicate the year of establishment for the first time.
An arrival map of the trees was generated according to the tree with the oldest establishment year for each plot,and the colonization year was interpolated for the sampled area using the inverse distance weighting technique.Also,an accumulation curve of the individuals established by species was performed,which allowed the identification of the oldest year of establishment of each species.The order of arrival of the species was evaluated using a sequence randomness test with the runs.test function (snpar package) (R-project Core R Core Team,2017).
2.6.Relationship between establishment and climatic factors
The relationship between tree establishment and inter-annual climatic conditions was assessed through a superposed epoch analysis(SEA),in which the climatic conditions that occurred five years before,two years after,and during the year of establishment of 5 or more trees were compared(Brewer et al.,2016).A ring-width index(RWI)was used as a proxy of regional precipitation,obtained from a climatic chronology elaborated from trees ofAbies religiosanear the Paricutín for the period 1970-2014 (Cerano-Paredes et al.,2014).Also,the years of the establishment were compared with the El Ni~no Southern Oscillation Index(ENSO) during the period 1970-2012 (Cook,2000;NOOA,2017),and with a local reconstruction of the Palmer Drought Severity Index(PDSI)that covers the period 1970-2008(Stahle et al.,2012).
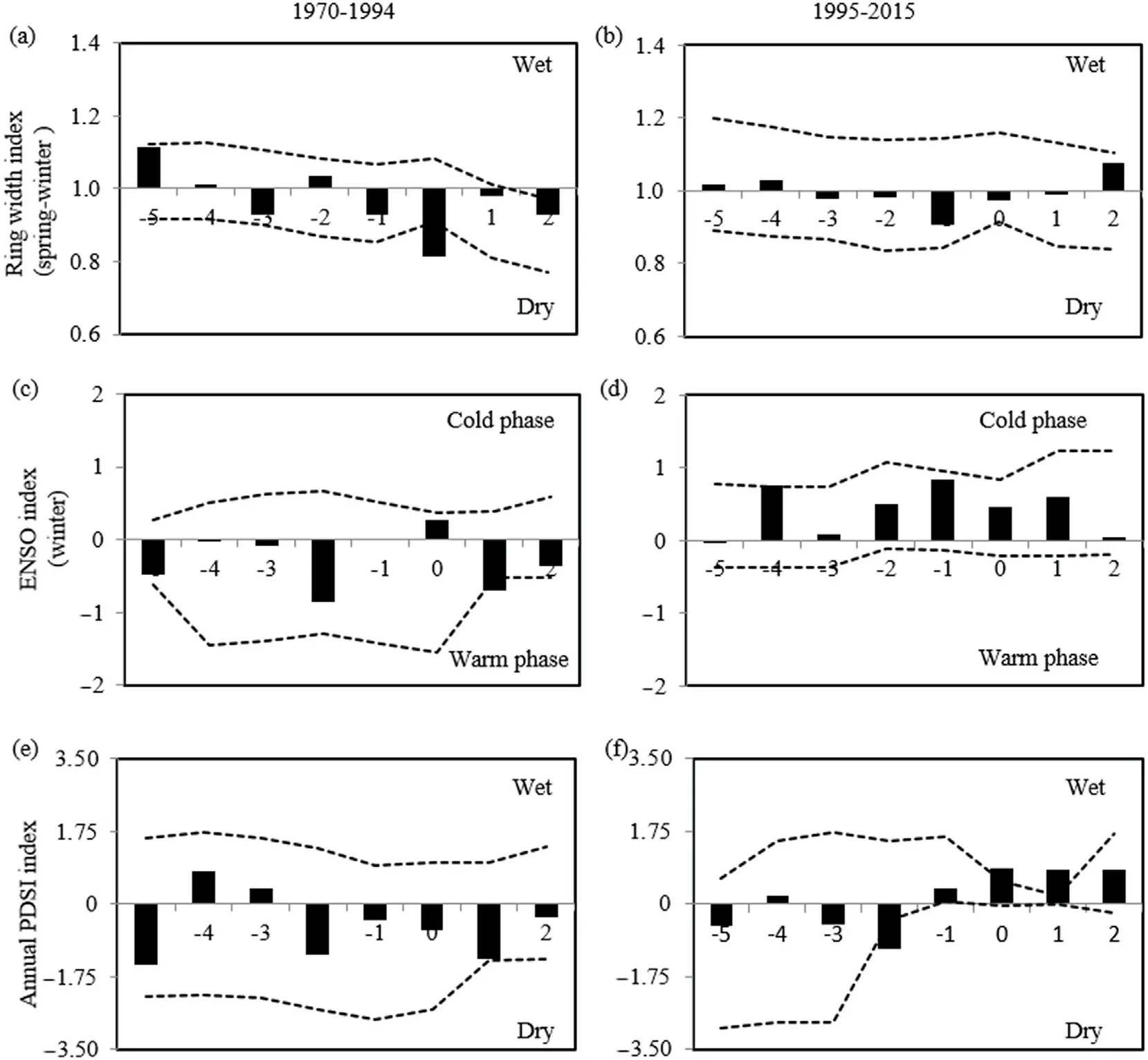
Fig.7.Superposed epoch analysis between the years of establishment and the climatic conditions for the period 1970 to 2015 in the Paricutín volcano.The upper part shows the relationship with the ring width index as a proxy for precipitation in the periods(a)1970-1994 and(b)1995-2015.The middle shows the El Ni~no Southern Oscillation index (c and d),while the Palmer Drought Severity Index (e and f) is in the lower part.The dotted lines represent the confidence intervals of the 1000 iterations with a confidence level of 95%.
The SEA was divided into two periods(1970-1994 and 1995-2015)because after 1995 the number of established individuals increased compared with the first period.A bootstrapping technique was used with 1000 iterations and 95% confidence intervals,in the software FHAES version 2.0.2(Brewer et al.,2016).The years of the establishment were compared also graphically,to identify whether an increase in tree recruitment was associated with wet years.
3.Results
3.1.Forest community composition
Eight pine species were collected:Pinus devonianaLindley(Michoacán pine),Pinus herrerae(Herrera pine),Pinus lawsoniiSchltdl.et Cham.(Lawson's pine),Pinus martinezii,Pinus montezumaeLambert(Montezuma pine),Pinus oocarpaSchiede ex.Schtldl.(Mexican yellow pine),Pinus pseudostrobusLindley (false white pine) andPinus teocoteSchiede ex.Schtldl.(Aztec pine).Only one fir species was found:Abiesflinckii.Data on frequency,density and mean basal area of each species can be found in Table S1.The species with the highest relative importance were,in descending order,Pinus pseudostrobus,Pinus montezumae,Pinus devoniana,andPinus teocote(Table 2).Moreover,Pinus pseudostrobusandPinus montezumaehad the highest relative dominance according to the basal area,whilePinus devonianaandPinus teocotehad a high relative density.Pinus martinezii,Pinus herrerae,andAbiesflinckiiwere the species with the lowest relative importance value.
3.2.Age structure
The establishment of the conifer trees began in 1970,whereas the youngest sampled tree established approximately in 2015,which was found at the volcano base.The general pattern of the tree establishment has been continuous since there were only 4 plots with peaks of tree establishment in the same year,in which abundant tree establishment occurred in 2001(Fig.3a),2003(Figs.3e),and 2012(Fig.3j and Fig.3n).In the rest of the plots,we observed that once the first individual was established,the following trees were established continuously,but not always in consecutive years.For example,in the plots located 750 and 500 m from the volcano in the western transect(Fig.3g and Fig.3j),we observed that there were 4 and 6 years between one established tree and the next one.
A clear spatial pattern was not found,suggesting that the oldest trees were found in the plots farthest from the volcano,but there was a large variation in the year of the arrival among the sampling plots(Fig.4).The oldest tree was established in 1970 at the 250-m plot in the western transect.In this transect,the most recent arrival year occurred at the 1000-m plot,whereas,at the volcano base,the establishment occurred in 1985.In the southwestern transect,this pattern was not very clear either,and,even in the plot farthest from the volcano(1,000 m),the colonization did not occur until 1990.In the southern transect,the plots closest to the volcano had more recent years of establishment.The relationship between the age of establishment and the distance to the volcano,however,was not significant (p>0.05),nor was it affected by the identity of the transect(p>0.05)(Fig.5a).
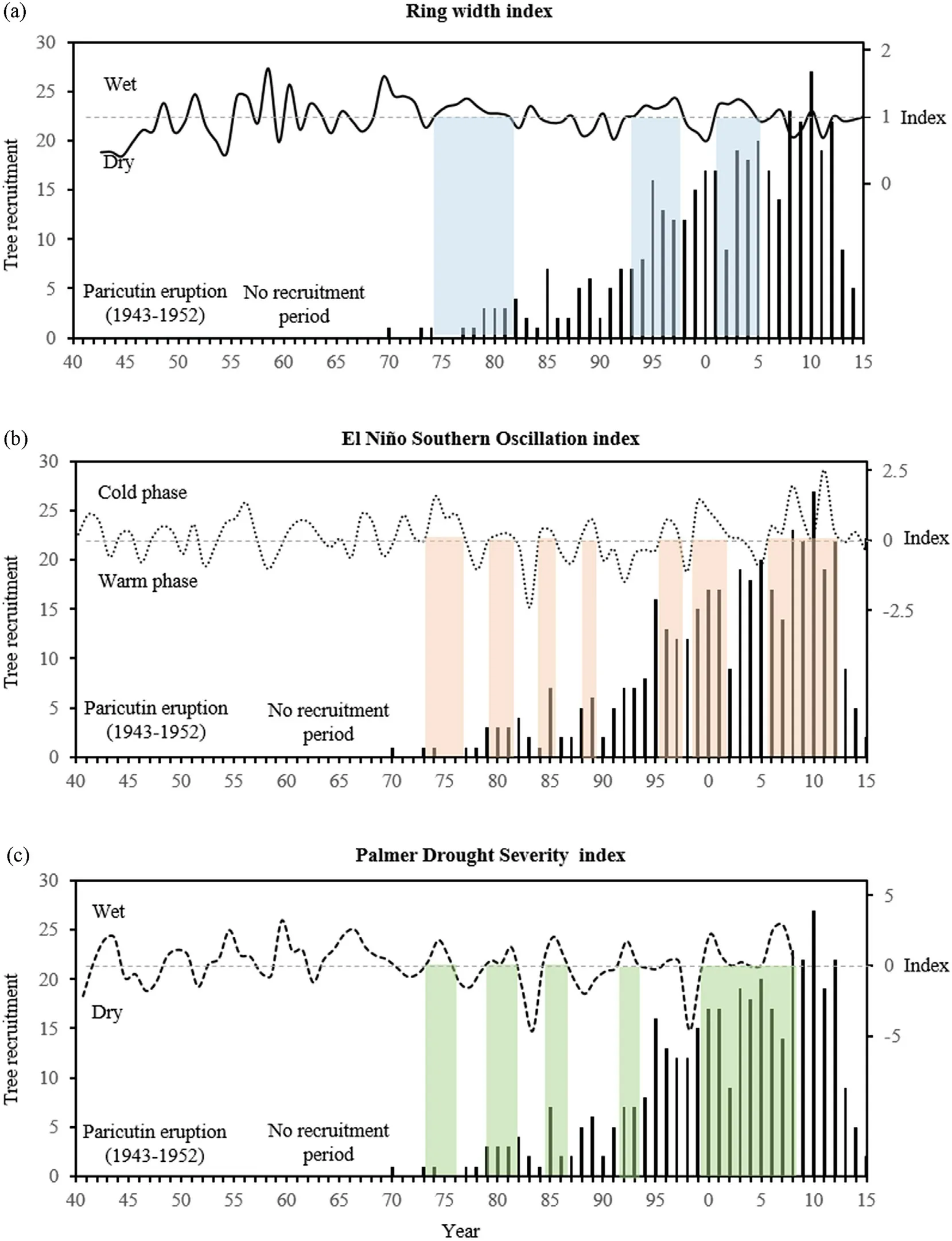
Fig.8.Relationship between(a)the temporal variation of the ring width index,as a proxy for vegetation,(b)El Ni~no Southern Oscillation index,and(c)the Palmer drought severity index over the establishment years of the trees at the Paricutín volcano.Bars indicate recruited trees,dotted lines represent climate indices,and vertical colored stripes show the wet periods of the three indices from 1970 to 2015.
Elevation did not influence the age of tree establishment (p>0.05)and was not influenced by the transect(p>0.05)(Fig.5b).However,as the slope of the terrain increased,the tree age decreased(df=399,t=-4.07,p<0.05),although with a low determination coefficient (r2=0.04)(Fig.5c).The age of establishment also differed significantly with respect to slope aspect (df=3,F=94.6,p<0.05) (Fig.5d),soil condition(df=2;F=22.73,p<0.05)(Fig.5e)and the conifer species(df=7,F=2.08,p<0.05) (Fig.5f).Age was also positively correlated with basal diameter(r2=0.34,df=399,t=14.43,p<0.05),then trees with a large diameter represent older individuals (Fig.5g).The number of species increased significantly as the sampling plots approached the volcano base(df=4,F=9.42,p<0.05)with a correlation coefficient ofr2=0.62(Fig.5h).Tree density decreased with the greater distance from the volcano (df=399,t=-1.97,p<0.05) but with a very low correlation(r2=0.007)(Fig.5i).More details on these analyses can be found in Table S2.
The most dominant species were the first to establish after the Paricutín eruption:Pinus pseudostrobusin 1970,Pinus montezumaein 1974,Pinus devonianain 1977,andPinus teocotein 1980.This pattern was very similar within the first decade in the western (Fig.6a) and southern(Fig.6c) transects,whereas tree establishment in the southwestern transect began until 1980 (Fig.6b).Pine species with less relative abundance(Pinus herrerae,Pinus lawsonii,andPinus martinezii)were also the last to become established between 1989 and 1995 (Fig.6d).The order of arrival of tree species,however,was completely random (p>0.05)in the three transects(Fig.6a-6c)and after grouping the data from all transects(Fig.6d).
The individual accumulation curves indicated that between 1970 and 1995,there was a period of slow recruitment for the populations of all tree species,except forAbiesflinckii,which arrived in the community by 1980 in the southwestern transect and its population has not grown since then.Starting in 1995,the populations of the dominant species increased steadily at an annual rate of 12.9%,until reaching an apparent period of relative stabilization in 2010(Fig.6).
3.3.Establishment and climate variation
The conifer recruitment was significantly associated with dry conditions during the establishment year in the period 1970-1994 (Fig.7a),but not in the period 1995-2015 (Fig.7b).There was no significant association with the occurrence of the warm or cold phase of the ENSO before or during the year of establishment of the conifers in the period 1970-1994 (Fig.7c),but in the period 1995-2015,there were wet conditions,although not significant,related to the cold phase of the ENSO (Fig.7d).The climatic conditions related to the PDSI did not influence the tree establishment in the period 1970-1994 (Fig.7e),however,during 1995-2015,tree establishment was related to wet conditions(Fig.7f).
The graphical comparison of the three evaluated indices and tree establishment confirmed that,as indicated by the SEA,during the first period the establishment of new trees occurred during dry years,while in the second period,the new individuals were established both in wet and dry years(Fig.8a).During 1995-2015,tree establishment was relatively influenced by the cold phase of the ENSO,when conditions were more humid(Fig.8b).Finally,it was observed that during a prolonged phase of humid conditions between 2000 and 2008,according to the PDSI,there was continuous and abundant recruitment in the community(Fig.8c).
4.Discussion
The conifer species have shown a great ability to establish in volcanic substrates,even a few years after the Paricutín eruption.AlthoughPinus montezumaeseedlings had been detected 8 years after the eruption(Eggler,1963),the oldest living tree in our sample(Pinus pseudostrobus)was established 10 years later (1970),which means that the suitable conditions for seed germination and conifer establishment was reached until that time,which allowed seedlings to survive and grow(Dale et al.,2005).Currently,the conifer populations are growing,probably becoming a young forest or reaching a mid-seral phase,where recruited saplings are derived from colonizer trees(Powell,2012).
The beginning of colonization was slightly longer than in other volcanic substrates.For example,Pinus hartwegiiLindley(Mexican mountain pine)took 10 years to establish in lahars of the Popocatépetl volcano in Mexico (Franco-Ramos et al.,2019),Pseudotsuga menziesiiFranco (Douglas-fir) arrived after 14 years in avalanche deposits near Mount St.Helens,in the United States (Pierson,2007),whereasLarix cajanderiMayr (Dahurian larch)appeared after 6 years in lahars at the Shiveluch volcano,Russia (Salaorni et al.,2017).It is important to mention that conifer arrival would be expected to take longer in lava flows than in lahars because igneous rocks are very porous,retain little water,and are difficult for roots to penetrate(Vilmundardóttir et al.,2018).
The rapid colonization of conifers in deposits of tephra and volcanic lavas of Paricutín can be explained by the presence of“safe sites”,where microclimatic and edaphic conditions gradually allow tree establishment(Cutler et al.,2008).The number of safe sites increases due to the incorporation of ash and tephra in the lava flows during the eruptive phase(Del Moral and Grishin,1999),as well as the entry of soil particles from sites with preserved vegetation through hydric or wind processes,which together form a suitable substrate for the growth and development of tree roots(Deligne et al.,2013).Once the first plants are established,biological weathering and the mechanical breaking of the rocks by the roots improve the edaphic conditions(Moldaway and Sparrow,2006).
In the case of Paricutín,the lava flows located towards the south and southwest of the cone were covered by ash and tephra during the eruptive period,while subsequent lava flows were covered by other layers of lava(Inbar et al.,1994).It may explain why primary succession has been concentrated in the oldest lava flows,while the rest of the area covered by younger lava remains devoid of arboreal vegetation(Lindig-Cisneros et al.,2006),which is similar to that reported for the El Jorullo volcano,where the oldest flows covered by tephra also show more vegetation(Deligne et al.,2013).Moreover,in the areas covered by tephra,erosion has played an important role in reducing the depth of this material(Inbar et al.,1994) since a depth greater than 30 cm of tephra significantly limits the establishment of conifers(Gómez-Romero et al.,2006).
In volcanic substrates,there is a strong relationship between tree age and distance from the preserved forests (Del Moral et al.,2005).However,the nearest trees to the Paricutín cone are not necessarily the youngest,which may be associated with the wind-dispersion of conifer seeds,increasing their probability to reach distances up to 120 m from the source of propagules (McCaughey et al.,1986).In the case of the species found at the Paricutín volcano,all of them release winged seeds,dispersed through the wind (Farjon et al.,1997).Besides,the arrival of conifers was not directional,but random,which is common in sites with primary succession(Garibotti et al.,2011).Nonetheless,the four species that colonized first currently have greater abundance and basal area,which is consistent with the trend in which the first arriving species tend to dominate the succession(Powell,2000).Also,these species have fast growth rates (Klepac,2001),of whichPinus pseudostrobusandPinus montezumaeare the dominant species in the nearby forest fragments(Fregoso et al.,2003).
The conifer community of the Paricutín volcano experienced an expansion process between 1995 and 2010.In this mid-seral phase,the seed bank might have begun to be fundamentally local,whereas the external sources of propagules decreased in importance(Del Moral et al.,2005).Additionally,completely stochastic processes,such as the dispersal of seeds from external sources,cease to operate,while deterministic processes,such as competition and facilitation,are more relevant (Cutler et al.,2008).Currently,only 70 years after the Paricutín eruptive period,there are already well-developed conifer stands,which is faster than in other volcanic sites recorded outside of the tropics,for example,600 years in Mount Hekla,Iceland (Vilmundardóttir et al.,2018),and 200 years in the Cascade Range,United States(Deligne et al.,2013).
The climate variability had a low influence on the establishment of conifers in the Paricutín substrates since trees were recruited in wet and dry years.This pattern has also been observed in other Mexican conifer forests (Sáenz-Ceja and Pérez-Salicrup,2020),which contrasts with the strong correlation between wet years and conifer recruitment in the semi-arid northern Mexico(Fulé and Covington,1997).On the one hand,rainfall regimes in tropical montane forests,where the Paricutín is located,may not be as contrasting as extra-tropical regions,so differences in tree recruitment during wet and dry years might not so clear (Astudillo-Sánchez et al.,2017).On the other hand,amelioration of substrates properties by a litter layer and biological weathering could have made suitable seed germination,even in dry years (Crisafulli et al.,2005).
In terms of ecological restoration,the fast establishment of conifers around the Paricutín suggests that the regeneration of tropical Mexican conifers can occur rapidly after high-magnitude disturbances,even in primary succession scenarios.Indeed,pine establishment began only two years after large fires in mixed pine-oak forests (Juárez-Martínez and Rodríguez-Trejo,2003) and seven years after pyroclastic flows at the Popocatépetl volcano (Franco-Ramos et al.,2019).At later seral stages,the basal area can be similar to that of mature pines only 28 years after high-severity fires(Quintero-Gradilla et al.,2019).It suggests that forest succession could be left in place without human intervention,as long as the sources of propagules are close to the affected areas and the soil conditions allow the establishment of seedlings (Ángeles-Cervantes and López-Mata,2009).
The Paricutín volcano represents a unique site for characterizing the primary succession after a high-magnitude disturbance.It is important to mention that disturbances such as fires or insect infestations have remained absent,but when they occur they could interrupt,reverse,and alter the successional trajectories(Riff et al.,2003).The establishment of conifers also skipped different stages of succession (the previous colonization by shrubs and later by broadleaf tree species),as has been recorded in some glaciers(Garibotti et al.,2011),which underscores the great adaptive capacity of conifers that inhabit the Paricutín region,within a region that host a high species diversity,including those of the genusPinus(Medina,2003).More research is needed on the successional dynamics after these disturbances,on the most recent established saplings after 2015,and the assessment of the influence of topographic and climatic variables,as well as the implications for ecological restoration and forest management.
5.Conclusions
The conifers around the Paricutin volcano have shown a great ability for colonizing substrates after 70 years of ending its eruptive period.The arrival of conifers began only 18 years after the last eruption in 1952,a process that has been random,unrelated to the distance from preserved forests,and not influenced by climate variability.The analysis of age structure was useful to identify a continuous establishment pattern,with a progressive increase of the population since 1995,followed by a population stabilization process from 2010.It is important to document the colonization of tree species in the more recent lava flows,where tree regeneration is low,and to include the most recent established saplings.It is also necessary to evaluate the ability of tropical coniferous species to regenerate in sites subjected to high-magnitude disturbances,information that could be useful to guide ecological restoration strategies.This study is a pioneer in applying dendrochronology to lava flows from recent volcanic activity,so it is recommended to replicate this type of study in both recent and ancient volcanic substrates,where successional patterns with annual resolution could be characterized.
Funding
The first author thanks the support of the Graduate Program on Biological Sciences at Universidad Nacional Autónoma de México(UNAM),and the National Council of Science and Technology (CONACyT,Mexico) for the scholarship received.We also thank the UNAM PAPIIT project IN209716 and CONACYT project 251694 for funding which allowed this research.The supporting sources had no involvement in the preparation of the data or manuscript,or the decision to submit for publication.
Availability of data and materials
The dataset used and/or analyzed during the current study is available from the corresponding author on reasonable request.
Author's contributions
JESC and BLSC conceived the study,with input from DRPS and JTSR.JESC and JTRS established the plots and collected the data.JESC and DRPS analyzed the data.All authors contributed to writing the manuscript and read and approved the final manuscript.
Ethics approval
Not applicable.
Consent for publication
Not applicable.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgments
The first author is grateful for the support of the Graduate Program in Biological Sciences at the Universidad Nacional Autónoma de Mexico,and the Mexican Council of Science and Technology (Conacyt) for the scholarship received to carry out graduate studies.The second author thanks the Undergraduate Program in Geosciences at the Escuela Nacional de Estudios Superiores(ENES),UNAM Campus Morelia.All the authors thank the authorities of the Caltzontzin Indigenous Community for the facilities granted for performing this study,and especially Jerónimo Ángel,Ignacio Cabrera,Luis Lara,and Ramón Guerrero for their support during the field sampling,as well as the Ecojardín at the Instituto de Investigaciones en Ecosistemas y Sustentabilidad for the space provided for the processing of dendrochronological samples.Finally,we thank Mary-Ann Hall for the editorial revision of the manuscript.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.fecs.2022.100007.

- Forest Ecosystems的其它文章
- Two-level optimization approach to tree-level forest planning
- No treeline shift despite climate change over the last 70 years
- Variation in water supply leads to different responses of tree growth to warming
- Active forest management accelerates carbon storage in plantation forests in Lishui,southern China
- Patterns and driving factors of leaf C,N,and P stoichiometry in two forest types with different stand ages in a mid-subtropical zone
- Forest height mapping using inventory and multi-source satellite data over Hunan Province in southern China