
Patterns and driving factors of leaf C,N,and P stoichiometry in two forest types with different stand ages in a mid-subtropical zone
2022-03-08 02:18YunniChangQuanlinZhongHongYangChaobinXuWeipingHuaBaoyinLi
Forest Ecosystems 2022年1期
Yunni Chang ,Quanlin Zhong,c,* ,Hong Yang ,Chaobin Xu ,Weiping Hua ,Baoyin Li,c,**
a Fujian Provincial Key Laboratory for Plant Eco-physiology,Fujian Normal University,Fuzhou,350007,China
b College of Geographical Sciences,Fujian Normal University,Fuzhou,350007,China
c State Key Laboratory for Subtropical Mountain Ecology of the Ministry of Science and Technology and Fujian Province,Fujian Normal University,Fuzhou,350007,China
d College of Environmental Science and Engineering,Fujian Normal University,Fuzhou,350007,China
e Department of Geography and Environmental Science,University of Reading,Reading,UK
f College of Ecological and Resources Engineering,Wuyi University,Wuyishan,354300,China
g College of Forestry,Fujian Agriculture and Forestry University,Fuzhou,350002,China
Keywords:Leaf stoichiometry C/N/P ratios Environmental factors Stand age groups Chinese fir forest Natural evergreen broadleaved forest
ABSTRACT Background: Carbon (C),nitrogen (N),and phosphorus (P)stoichiometry is a key indicator of nutrient utilization in plants,and C/N/P ratios are related to the life histories and adaptation strategies of tree species.However,no consensus has been reached on how leaf stoichiometric characteristics are affected by forest type and stand ages.The relationships between leaf stoichiometry and geographical,meteorological,and soil factors also remain poorly understood.Methods:Leaf and soil were sampled from forest stands of different age groups(young,middle-aged,near-mature,and mature)in two forest types(Chinese fir(Cunninghamia lanceolata)forests and evergreen broadleaved forests).The relationships between leaf C,N,and P stoichiometric parameters and geographical,meteorological,and soil factors were analysed by using redundancy analysis (RDA) and stepwise linear regression analysis.Results: Leaf C concentrations peaked in the near-mature stands with increasing age irrespective of forest type.Leaf N and P concentrations fluctuated with a rising trend in Chinese fir forests,while decreased first and increased later from young to mature phases in natural evergreen broadleaved forests.Chinese fir forests were primarily limited by N and P,while natural evergreen broadleaved forests were more susceptible to P limitation.Leaf C,N,and P stoichiometric characteristics in Chinese fir forests were mainly affected by the soil total P concentration (SP),longitude (LNG),growing season precipitation (GSP) and mean temperature in July (JUT).The leaf C concentration was mainly affected by GSP and JUT;leaf N and P concentrations were both positively correlated with LNG;and leaf P was positively correlated with SP.In evergreen broadleaved forests,however,leaf stoichiometric parameters displayed significant correlations with latitude (LAT) and mean annual precipitation(MAP).Conclusions: Leaf stoichiometry differed among forest stands of different age groups and forest types.Leaf C,N,and P stoichiometry was primarily explained by the combinations of SP,LNG,GSP and JUT in Chinese fir forests.LAT and MAP were the main controlling factors affecting the variations in the leaf C,N,and P status in natural evergreen broadleaved forests,which supports the temperature-plant physiological hypothesis.These findings improve the understanding of the distribution patterns and driving mechanisms of leaf stoichiometry linked with stand age and forest type.
1.Introduction
Ecological stoichiometry addresses the mass balance of multiple nutrient elements at scales from organs to ecosystems.Element ratios have been widely used to analyse ecosystem function and nutrient limitations,providing new perspectives to understand energy flow and nutrient cycling in ecosystems(Elser et al.,2000;Wu et al.,2012;Li et al.,2018a;Bai et al.,2020;Zhu et al.,2020).Carbon(C),nitrogen(N),and phosphorus (P) are the three main elements which play vital roles in plant functions such as affecting plant growth and adaptations in terrestrial habitats (Cao et al.,2017).C provides a structural basis for plants,and N and P are essential nutrient elements and important limiting factors for primary production(Zhang et al.,2019a).Leaves are the primary photosynthetic organs and are sensitive to environmental changes(Liu et al.,2017;Guo et al.,2018).Leaf C,N,and P statuses are associated with many critical aspects of plant growth,reproduction,and ecosystem functions and can be used as indicators of how plants use nutrients and respond to environmental changes(Qin et al.,2018).Leaf stoichiometric characteristics are considered to represent both the heritable characteristics of plants and their acclimations to specific environmental conditions.Therefore,research on leaf stoichiometric characteristics can advance the understanding of biogeochemical cycling and the structure and functions of ecosystems(Bai et al.,2020;Zhu et al.,2020).
Leaf stoichiometry varies with forest types and growth stages(Wang et al.,2016a).As plants grow,the element ratios of leaves usually change but the response patterns could be divergent(Wang et al.,2016a;Li et al.,2018b).For example,in some plantations,no significant trend was observed for leaf C and N concentrations and C/N ratios along a chronosequence,whereas the leaf C/P and N/P ratios were markedly lower in young forests than that in middle-aged and mature forests due to a decrease in leaf P concentrations(Cao et al.,2017;Zhang et al.,2018b).Li et al.(2018)suggested that the concentrations of C,N,and P in black locust (Robinia pseudoacacia) leaves increased with an increase in stand age (Li et al.,2018a).Data collected from secondary forests around the world showed that plant C/N ratios increased significantly with an increase in stand age (Yang and Luo,2011).In an evergreen alpine shrub(Rhododendron agglutinatum)forest in the Miyaluo Natural Reserve,West China,younger leaves had higher concentrations of nutrients,and C/N and C/P ratios increased with stand age,whereas juvenile leaves had the lowest N/P ratio (Wang et al.,2016a).The C/N/P stoichiometry in tree leaves lacked consistent variation with age in Chinese pine (Pinus tabulaeformis) forests (Wang et al.,2018).These age-related changes in leaf nutrient concentrations may be a consequence of different adaptive strategies adopted by different plant species (Tegischer et al.,2002).Since plant age may explain the inconsistent results regarding leaf C,N,and P statuses in different climatic regions,the effects of stand age on leaf C,N,and P concentrations and ratios in various forest types in the same climatic region need more exploration.
Plant life forms have been found to influence leaf C,N,and P concentrations and ratios.Conifers have significantly lower leaf N(or higher leaf C) concentrations than that of broadleaved trees,but there is no significant difference in leaf P concentrations between the two types of plants in Beijing and its surroundings areas in China(Han et al.,2009).Wu et al.(2010)suggested that in the Pearl River Delta,South China,leaf C and N concentrations were higher in coniferous forests than that in evergreen broadleaved forests,and the opposite pattern was found for leaf P concentrations.However,the leaf N and P concentrations and N/P ratios were higher in evergreen broadleaved forests than that in coniferous forests across 654 terrestrial plant species along a north-south transect in eastern China (Ren et al.,2007).These inconsistent results demonstrated that the variation in leaf stoichiometry between the two forest types is still an unaddressed issue.
Abiotic factors (such as geographical,meteorological,and soil factors) may affect leaf nutrient concentrations (Sardans et al.,2011;Guo et al.,2018;Song et al.,2018).Regional differences in these factors could result in differences in the stoichiometric characteristics of plants.The temperature-plant physiological hypothesis proposes that at low temperatures,plant leaves contain high concentrations of N and P to offset the reduced rate of biochemical reactions caused by the low temperature-induced decrease in the efficiency of N-rich enzymes and P-rich RNA(Reich et al.,2004).The global studies found that leaf N and P concentrations of 1,280 species increased from the tropics to cooler and drier mid-latitudes(Reich et al.,2004;Liu et al.,2021).The leaf N and P concentrations of 753 Chinese terrestrial plant species increased with increasing latitude (LAT) and decreasing mean annual temperature(MAT),whereas the N/P ratios had no obvious changes (Han et al.,2005).The leaf N/P ratios increased with increasing LAT and decreasing MAT on the Loess Plateau,north China,whereas leaf N and P contents were independent of LAT(Zheng et al.,2007;Liu et al.,2021).The leaf C,N,and P stoichiometry ofMachilus pauhoiunderstorey seedlings was affected by both longitude(LNG)and MAT(Yu et al.,2018a).The leaf N concentrations inQuercusspecies across China increased linearly with LNG,while quadratic relationships were found between leaf P concentrations and leaf N/P ratios and LNG(Wu et al.,2012).These inconsistent results demonstrated the reginal variations in stoichiometric patterns.
Soil properties directly affect plant growth.The biogeochemical hypothesis emphasizes the crucial role of soil nutrient availability in determining leaf nutrient contents (Hu et al.,2021).Numerous studies have demonstrated that the stoichiometric characteristics of plants are affected by soil characteristics(Fitter et al.,2009;Cao et al.,2016).Fan et al.(2015)demonstrated that the C/N/P ratios in soil directly reflected soil fertility,regulated vegetation patterns,and indicated the plant nutritional status.Liu et al.(2021)pointed out that soil properties made larger contribution than climatic factors to the N/P stoichiometric variations in Qinghai spruce across the Qilian Mountains,West China.However,most studies have focused on the impact of a single environmental factor on leaf C,N,and P concentrations and ratios,whereas only a few studies have focused on the combined effects of multiple factors(Yu et al.,2018a;Liu et al.,2021).
Plant age and forest type can cause variations in leaf C,N,and P stoichiometry in different regions;however,to our knowledge,the effects of stand age on leaf C,N,and P stoichiometry in various forest types in the same climatic region are still unclear.Additionally,the combined effects of multiple factors have been little explored.Therefore,considering forest stands as a basic unit to study the relationship between leaf stoichiometric characteristics and abiotic factors (geographical,meteorological,and soil factors)can be an innovative method to investigate the variation between different forest stands composed of trees of different functional types and ages.Subtropical forests in China are the only oasis belts in the same latitude region in the world,playing crucial roles in biodiversity conservation,climate regulation,and global carbon cycle(Zhang et al.,2015,2018b;Lai et al.,2016).Chinese fir [Cunninghamia lanceolata(Lamb.)Hook.]is one of the most important native and evergreen coniferous timber species in subtropical China(Liu et al.,2020b).Chinese fir plantations account for 19% of the total forest area (8.93 million hectares) and 25% of all plantation stocks (625 million m3) in China (Zhang et al.,2020).It has been widely applied as an economic tree species and represents the most commonly planted evergreen coniferous species in China,due to its excellent timber quality and high yield(Zhang et al.,2018a).Evergreen broadleaved forests are the typical and widespread zonal vegetation type in this region (Yan et al.,2008).Evergreen broadleaved forests mainly consist of Fagaceae,Lauraceae,Theaceae,Elaeocarpaceae,and other families (Ding et al.,2015).Therefore,Chinese fir and evergreen broadleaved forests were selected for this study.
Here,we surveyed Chinese fir forests and evergreen broadleaved forests along age sequences in Southeast China.We sampled the leaves of the dominant species in each forest stand and analysed the variations in leaf C,N,and P stoichiometry and abiotic factors.The following two hypotheses were tested in this study:(1)there are distinct differences in leaf C,N,and P stoichiometry among stand developmental stages of different forest types(Cao et al.,2017);and(2)the relative contributions of geographical,meteorological and soil factors to the changes in leaf C,N,and P stoichiometry differ among different forest ecosystems (Liu et al.,2021).We aimed to(1)evaluate the differences in leaf C,N,and P stoichiometric characteristics in Chinese fir forests and evergreen broadleaved forests in different age groups,and(2)determine the main environmental factors affecting leaf C,N,and P stoichiometric characteristics in the two forest types with different stand ages.These results will be helpful to understand the relationships between leaf C,N,and P stoichiometry and abiotic factors,providing new insights into forest adaptation mechanisms to environmental change and forest type conversion.The findings can also provide theoretical guidance for carbon neutrality actions.
2.Materials and methods
2.1.Study area
The study area (117°12′-119°5′E,25°26′-27°48′N) is located in Fujian Province,Southeast China,which is dominated by mountains and hill with altitude from 97 to 756 m.It is also in the middle subtropical monsoon climate zone,with an average temperature of 18.86°C,an annual frost-free period of 255-336 days,and an annual precipitation of 1,697 mm.Based on Chinese soil taxonomy,the soil is classified as lateritic soil.Chinese fir forests and evergreen broadleaved forests are the two main forest types in the research area.
2.2.Field sampling and laboratory analyses
According to the Second Type Inventory of Forest Resource of Fujian Province conducted in 2008,we selected representative sites for Chinese fir forests and evergreen broadleaved forests in the research area.The sites were surveyed from August to October 2011 using the quadrat sampling method.Based on the average age of the dominant species according to the China National Forest Inventory Technical Regulations(State Forestry Bureau of China),the forests were classified into four age groups:young forests (YF),middle-aged forests (abbreviated as HF to avoid confusion with mature forests),near-mature forests (NF),and mature forests (MF).Chinese fir forests were divided into the following age groups:YF,≤10 years;HF,11-20 years;NF,21-25 years;and MF,26-35 years.Evergreen broadleaved forests were divided into the four age groups as follows:YF,≤40 years;HF,41-60 years;NF,61-80 years;and MF,81-120 years.Research sites were selected by considering forest type,forest age structure,proportion of forest age and forest management.A total of 17 sites were selected in this study(Table 1).Regarding site preparation techniques,we used hole site preparation by digging holes of 40 cm×40 cm×30 cm(length×width×depth)for Chinese fir plantations.The evergreen broadleaved forest consisted of tree species,includingCastanopsis carlesii,C.eyrei,C.fargesii,Quercus glauca,andSchima superba;as well as shrubs,includingCamellia oleifera,Loropetalum chinensis,Itea oblonga,Syzygium buxifolium,andRapanea neriifolia;and herbs,includingDicranopteris dichotoma,Phragmites australis,Carex tristachya,andWoodwardia japonica.The selected sites were located far from settlements,and the evergreen broadleaved forests were free from human interference.
Three plots,50 m×20 m(length×width),were selected at each site;the distance among the plots was greater than 100 m.Each plot was divided into 10 m×10 m(length×width)quadrats,and the diameter at breast height (DBH ≥2 cm) and heights of all trees in each plot were measured(Table 1).The mean DBH and mean height were calculated at each site in the two forest types.Based on their dominance in the research area,the following species were selected:Castanopsis carlesii,C.eyrei,C.fargesii,Quercus glauca,Schima superba,Neolitsea sericea,Ilex litesaefolia,Alniphyllum fortunei,Elaeocarpus sylvestris,Camellia oleifera,Altingia gracilipes,Adinandra millettii,Cinnamomum japonicum,Elaeocarpus decipiens,Michelia maudiae,Sapium discolor,Daphniphyllum oldhami,C.kawakamii,C.fissa,andIlex latifolia.Three healthy trees with the DBH and height close to the mean DBH and mean height were selected to represent standard trees for each of the selected tree species.Within each sampling site,leaves of the selected standard trees of the dominant species were collected from the upper,middle,and lower parts of the canopy and four cardinal compass directions using a pole pruner.After measuring the leaf fresh mass,all samples were oven-dried at 70°C for 48 h to a constant mass and weighed to obtain the dry mass.
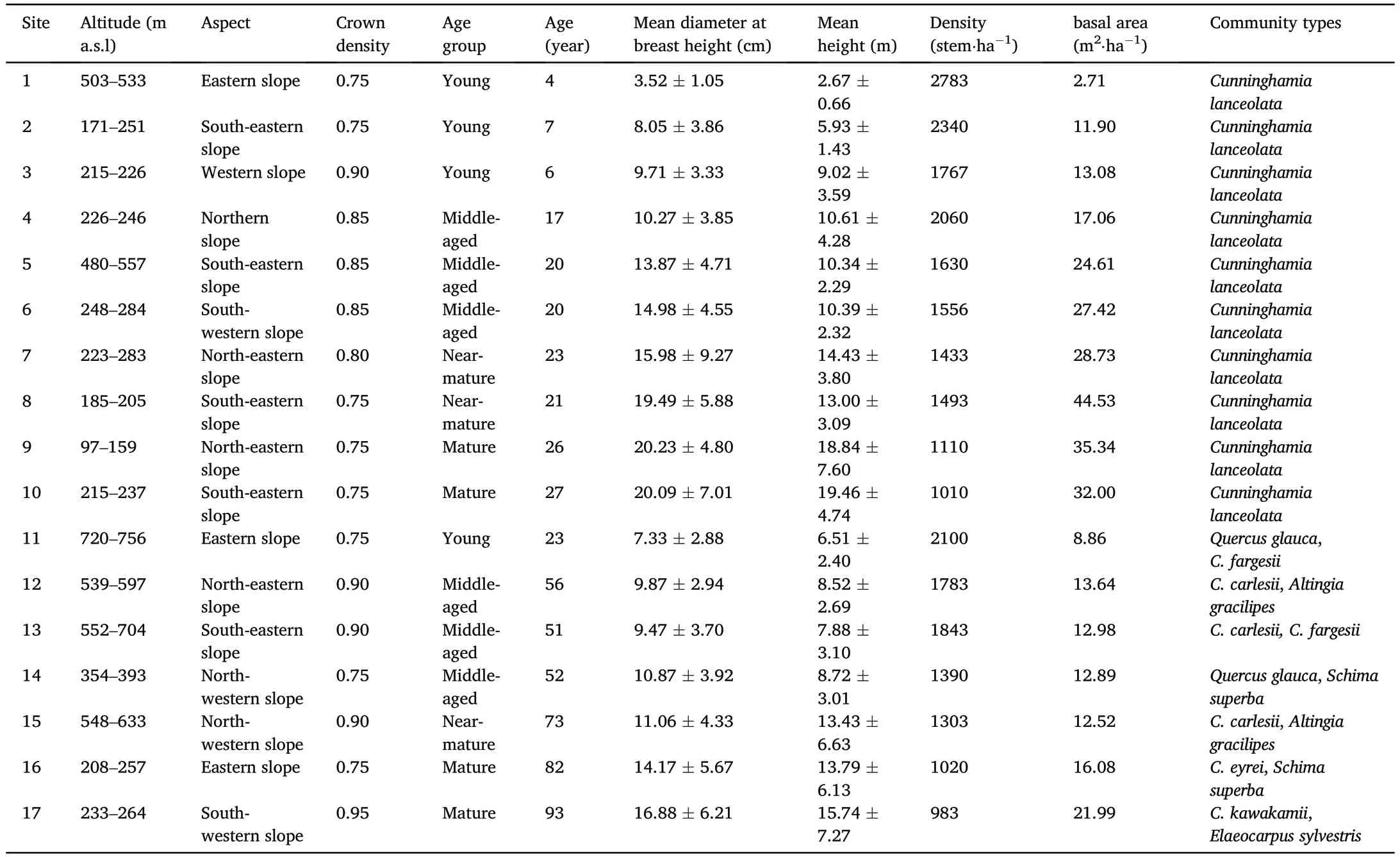
Table 1 Stand characteristics of Chinese fir(Cunninghamia lanceolata) forests and evergreen broadleaved forests (data are mean ± standard deviation).
Three soil cores were collected from the upper 20-cm soil layer along a diagonal direction across each plot using an auger(5-cm diameter)and mixed to form one soil sample per plot.After roots and stones were removed,the soil samples were air-dried and sieved through a 2-mm mesh sieve.
The dried leaves and air-dried soil samples were ground to fine powder to determine the C,N,and P concentrations.The total C and N concentrations of the leaf and soil samples were analysed using an Elemental Analyser (Elementar Vario EL III;Elementar,Langenselbod,Germany).The total P concentrations of leaves and soil samples were analysed using the molybdenum-antimony anti-colorimetric method(Chang et al.,2013).All chemical analyses were conducted in triplicate.The concentrations of C,N,and P in the samples were expressed as mass concentrations.
2.3.Geographical and meteorological data acquisition
Geographical factors (LNG and LAT),meteorological factors (MAT;MAP;mean temperature in July,JUT;mean temperature in January,JAT;growing season precipitation,GSP;and relative humidity,RH),and soil factors(soil total C content,SC;soil total N content,SN;and soil total P content,SP) were also analysed in this study.LAT and LNG were measuredin situusing a handheld GPS (GPSMAP639sc,Maibang,Nanjing,China).The meteorological data from 1981 to 2010 were obtained from the National Weather Service Meteorological Data Center(data.cma.cn/) (Table 2).Soil samples were collected and analysed as described in the field sampling and laboratory analyses section above.

Table 2 Geographical,meteorological,and soil factors of Chinese fir (Cunninghamia lanceolata)forests and evergreen broadleaved forests.
2.4.Statistical analysis
Normality tests indicated that all data met the assumptions of normality and homogeneity.Analysis of variance (ANOVA) and least significant difference (LSD) multiple comparisons were conducted to identify significant differences in the leaf C,N,and P stoichiometric characteristics among the forest types and stand ages.Significance was set atP<0.05.ANOVA was performed using OriginPro software(version 2020,Origin Laboratories,Massachusetts,USA).To further assess the effects of abiotic factors on leaf C,N,and P stoichiometry,redundancy analysis(RDA)and stepwise linear regression analysis were performed to evaluate the relationships between the leaf C,N,and P stoichiometry and geographical,meteorological,and soil factors.RDA was performed to choose key factors affecting leaf C,N,and P stoichiometry.Stepwise linear regression analyses were performed to select the best regression model with the lowest Akaike information criterion(AIC)value.RDA and stepwise linear regression analyses were conducted in R (R Core Team,2019).
3.Results
3.1.Leaf C,N,and P concentrations
The mean leaf concentrations of C,N,and P in Chinese fir forests were 461.24,16.56,and 0.99 mg•g-1,respectively,while the mean leaf concentrations of C,N,and P in evergreen broadleaved forests were 460.31,16.51,and 0.71 mg•g-1,respectively.There were no significant differences in leaf C and N concentrations between the two types,whereas leaf P concentrations in Chinese fir forests were 39% higher than that in evergreen broadleaved forests.
Leaf C concentrations differed markedly among the four age groups in the two forest types.In Chinese fir forests,leaf C concentrations decreased first with stand age,then increased,and decreased later again.The highest values were observed in YF stands and NF stands,whereas the lowest values appeared in MF stands.In evergreen broadleaved forests,leaf C first increased and then decreased with stand age;the peak occurred in NF stands,whereas the lowest value appeared in YF stands.Leaf C concentrations in Chinese fir forests were higher than those in evergreen broad-leaved forests in YF stands,whereas leaf Cconcentrations in evergreen broad-leaved forests were higher than those in Chinese fir forests in MF stands(Fig.1a).
Leaf N and P concentrations differed markedly among Chinese fir forests of different stand ages.The highest leaf N and P concentrations were observed in MF stands,whereas the lowest leaf N and P concentrations appeared in NF stands.Leaf N and P concentrations in MF stands were markedly higher than those of the other three age groups.However,in the evergreen broadleaved forests,the highest leaf N and P concentrations were found in YF stands,whereas the lowest leaf N and P concentrations were observed in NF stands.In addition,MF evergreen broadleaved forests had markedly lower leaf N and P concentrations than MF Chinese fir forests.In YF stands,leaf N concentrations in Chinese fir forests were markedly higher than those in evergreen broadleaved forests(Fig.1b and c).
3.2.Leaf stoichiometric ratios
The mean leaf C/N,C/P,and N/P ratios were 30.88,492.94,and 16.61,respectively,in Chinese fir forests,while the average leaf C/N,C/P,and N/P ratios were 29.54,775.69,and 25.54,respectively,in evergreen broadleaved forests.The C/P and N/P ratios in evergreen broadleaved forests were generally higher than those in Chinese fir forests,while there were no significant differences in C/N ratios between the two forest types in any of the age groups.
Leaf C/N ratios differed markedly among stands of different ages.In Chinese fir forests,the C/N ratios were higher in NF stands (40.33),followed by YF stands(37.89),HF stands(25.49),and MF(19.00)stands.Leaf C/N ratios in evergreen broadleaved forests were markedly higher in NF stands than in the other three age groups.The two forest types differed significantly in their C/N ratios in YF and MF stands(Fig.1d).

Fig.1.Stoichiometric characteristics of leaf carbon(C),nitrogen(N),and phosphorus(P)in different forest types(Chinese fir(Cunninghamia lanceolata)and evergreen broadleaved forests)and age groups.Different lowercase letters indicate significant differences(P<0.05)among different age groups for the same forest type,while different uppercase letters indicate significant differences between two different forest types for the same age group.YF:young forest,HF:middle-aged forest,NF:near-mature forest,and MF:mature forest.
Leaf C/P and N/P ratios differed significantly among the four age groups in both Chinese fir and evergreen broadleaved forests.The leaf C/P ratios in NF stands were higher in the two forests.In Chinese fir forests,the highest leaf N/P ratio was observed in HF stands;in evergreen broadleaved forests,the highest leaf N/P ratio was found in MF stands.The two forest types differed markedly in leaf C/P and N/P ratios in HF,NF,and MF stands(Fig.1e and f).
3.3.Relationships between leaf C,N,and P stoichiometry and environmental factors
In Chinese fir forests,the first axis length of the detrended correspondence analysis (DCA) was 0.2 (<3);therefore,RDA was conducted to identify the main factors that affecting leaf C,N,and P stoichiometric characteristics.The first RDA axis accounted for 86.75% of the total variation,with the strong loadings of LNG,SP,JUT and GSP (Fig.2a).The second RDA axis accounted for 9.09% of the total variation.Therefore,LNG,SP,JUT and GSP were identified as the dominant factors influencing leaf C,N,and P stoichiometry in Chinese fir forests.
In evergreen broad leaved forests,the first RDA axis accounted for 98.73%of the total variation,with strong loadings of LAT,SP,and MAP(Fig.2b),and the second RDA axis accounted for only 0.68%.Thus,LAT,SP,and MAP were identified as the main factors affecting leaf C,N,and P stoichiometry in evergreen broadleaved forests.
Stepwise linear regression analyses between the abiotic factors and leaf C,N,and P stoichiometry were conducted.We selected the best regression models with the lowest AIC (Table 3).In Chinese fir forests,leaf C concentrations were negatively correlated with JUT and GSP(R2=0.28,P=0.004).Leaf N concentrations were positively correlated with LNG (R2=0.22,P=0.005).Leaf P concentrations were positively correlated with LNG and SP(R2=0.50,P<0.001).Leaf C/N ratios were significantly negatively correlated with JUT(R2=0.11,P=0.039).Leaf C/P ratios were negatively correlated with LNG and SP (R2=0.44,P<0.001).Leaf N/P ratios were significantly negatively correlated with SP and positively correlated with JUT and GSP(R2=0.39,P=0.001).
In evergreen broadleaved forests,leaf C concentrations were significantly negatively correlated with LAT and positively correlated with SP(R2=0.76,P<0.001).Leaf N and P concentrations were significantly positively correlated with LAT (R2=0.75,P<0.001;R2=0.95,P<0.001),and leaf P concentrations were also negatively correlated with MAP.The leaf C/N,C/P,and N/P ratios were negatively correlated with LAT (R2=0.74,P<0.001;R2=0.93,P<0.001;andR2=0.96,P<0.001,respectively),and the leaf C/P and N/P ratios were also positively correlated with MAP(Table 3).The standardized regression coefficients of LAT were larger than those of MAP,indicating that LAT had the largest influence on leaf P,C/P and N/P.

Table 3 Regression models for leaf C,N,and P stoichiometry of Chinese fir (Cunninghamia lanceolata) forest and evergreen broadleaved forest and environmental factors.
4.Discussion
4.1.Leaf C,N,and P stoichiometric characteristics in forest stands among different age groups
The stoichiometric ratios of C,N,and P are pivotal indicators of nutrient utilization and allocation in plants and are related to the life histories and adaptation strategies of plants (Zhu et al.,2019).The distribution ratios of C,N,and P within a plant vary with stand development(Wang et al.,2016a).In this study,leaf C,N,and P concentrations were significantly affected by stand age,indicating the differences in ecological adaptability of plants at various stages of development(Zhang et al.,2019b).
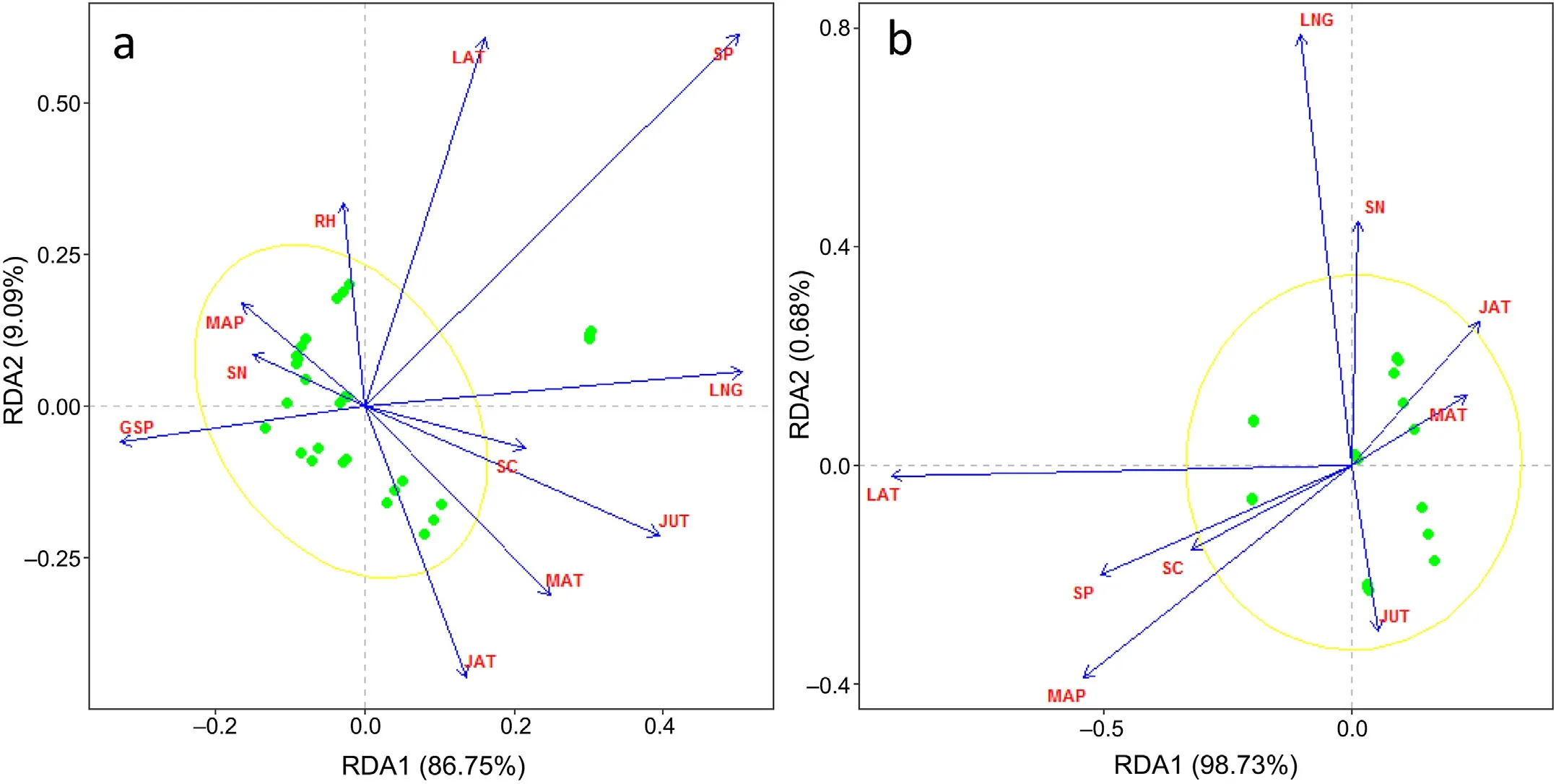
Fig.2.Redundancy analysis of leaf carbon(C),nitrogen(N),and phosphorus(P)stoichiometric characteristics and environmental factors in Chinese fir(Cunninghamia lanceolata) forests (a) and evergreen broadleaved forests (b).LAT,latitude;LNG,longitude;MAT,mean annual temperature;MAP,mean annual precipitation;JUT,mean temperature in July;JAT,mean temperature in January;RH,relative humidity;GSP,growing season precipitation;SC,soil total C in the 0-20 cm soil layer;SN,soil total N in the 0-20 cm soil layer;SP,soil total P in the 0-20 cm soil layer.Yellow ellipses indicate confidence ellipses.(For interpretation of the references to color in this figure legend,the reader is referred to the Web version of this article.)
Plant species with different life forms often display variable stoichiometry in an ecosystem,indicating the contrasting C,N,and P utilization strategies.In stands ranging from young to mature,leaf C concentrations in NF stands were higher regardless of forest type.This indicates that the tree leaves in NF stands had higher organic compound concentrations and a higher C storage capacity,compared with stands of other ages.A possible explanation for this is the variation in photosynthetic characteristics.Plant age affects photosynthetic performance(Bielczynski et al.,2017);because of higher photosynthetic rates,carbon storage in stands at the NF stage may be much higher.With increasing age,the net photosynthetic rate ofPicea schrenkianafirst increased and later decreased(Zang et al.,2009).
Leaf N and P concentrations fluctuated with the increasing stand age in Chinese fir forests(Fig.2).Leaf N and P concentrations in MF Chinese fir stands were significantly higher than those in the other three age groups,indicating that trees accumulate nutrients as they develop and grow.A possible reason is that soil P concentrations first decreased and then increased along the gradient of stand age(Cao et al.,2019).Similar findings were observed in black locust plantations of different ages in the Loess Plateau (Ma et al.,2017),north China,probably due to root absorbing and transporting nutrients from the soil and storing them in the stems and leaves with the growth of plants.
In natural evergreen broadleaved forests,leaf N and P concentrations first decreased and later increased from young to mature stages,and they were highest in YF stands,which is similar to previous findings (Gao et al.,2007).Most plant nutrients are limited by their availability in soil(Townsend et al.,2007;Bui et al.,2013),and nutrient concentrations in plant tissues are related to their concentrations in the soil.Plant litter makes large contribution to nutrient cycling between plants and soils(Ågren,2004).Soil N mainly originates from litter return and atmospheric N deposition.YF stands were characterized by a low canopy density,sufficient light,and high temperatures.These characteristics might lead to higher microorganism activity in the soil,which in turn leads to enhanced decomposition rates of litter and accelerated return of litter N to the soil.The increase in soil available N is beneficial for plants to increase their N concentrations.In MF stands,however,individuals were larger,and the litter yield was higher than those of younger stands.Therefore,the amount of N returned to the soil was higher,resulting in increases in soil N.Soil P originates from rock weathering and litter return,and the patterns of soil P were similar to those found for soil N.Soil P concentrations first declined and later increased.Because of the close relationship between soil and plant nutritional status,changes in soil nutrient concentrations may affect plant nutrient concentrations.The differences between the two forest types in this study may be caused by differences in tree species and nutrient resorption efficiency or by the influence of species richness,microbial activities,and litter decomposition rates (Wang et al.,2018).
Leaf C/N and C/P ratios,which are important physiological indices,are related to plant growth rate and plant carbon assimilation capacity(Ågren,2004;Frost et al.,2005;Wang et al.,2018).Leaf C/N and C/P ratios in NF stands were higher than in stands of other ages in the two types of forests (Fig.1),indicating a high carbon assimilation capacity.Leaf N/P ratios can be used as an index to diagnose N saturation and as thresholds to determine nutrient limitations(Güsewell et al.,2003;Elser et al.,2007;Wang et al.,2018).Leaf N/P ratios varied significantly across age groups in the two forest types.Chinese firs were limited by different nutrients at various stages.With regard to the age sequence,the age-related increase in leaf N/P ratios indicated that natural evergreen broadleaved forests became progressively limited by P.These results are similar to those reported for black locusts in previous studies(Fan et al.,2015;Cao et al.,2017).
4.2.Factors influencing leaf C,N,and P stoichiometry
Environmental changes can affect the stoichiometric characteristics of plants.Previous studies reported that leaf C,N,and P stoichiometry was affected by temperature (Reich et al.,2004),precipitation (Townsend et al.,2007),soil nutrients (Cai et al.,2019),and plant functional groups(Zhang et al.,2012).In Chinese fir forests,leaf C was significantly negatively correlated with JUT,which may be due to the declined photosynthetic rates with enzyme deactivation and denaturation at high temperatures in summer(Moore et al.,2021).Another possible reason is that carbon assimilation at high temperature was limited by stomatal and mesophyll conductance (Zhu et al.,2021).Leaf N and P concentrations increased with increasing LNG.Similarly,one study revealed the different N deposition fluxes along the east-west gradient (Zheng et al.,2014).Our results could be caused by the fact that a large amount of deposited atmospheric reactive N was absorbed by plants,thus leaf N concentration increased(Wang et al.,2016b;Xiao et al.,2016;Liu et al.,2019).Nitrogen deposition promotes plant absorption of soil P by increasing the activity of root phosphatase or by stimulating a plant to use the remaining N in the plant to produce more phosphatase(Yu et al.,2018a).The biogeochemical hypothesis emphasizes that soil nutrient availability is the dominant factor of leaf element contents (Chen et al.,2013;Liu et al.,2020a).Most plant nutrients are determined by their availability in soil and the retranslocation of nutrients between the soil and plants (Fan et al.,2015).Previous studies indicated that leaf stoichiometry was determined primarily by the soil parent material and the degree of soil weathering (Fan et al.,2015;Liu et al.,2021).For this reason,soil P was the main predictor of leaf P concentrations in this study.Similar to our results,some findings demonstrated that the relationships between plant nutrients and soil nutrients.For example,plant P and N/P are largely shaped by changes in soil P (Liu et al.,2019).Nutrient concentrations in soil and plants were tightly linked inEucalyptusplantations,and P limitation increased with stand age (Fan et al.,2015).Hedin(2004)reported that the dynamics of plant C,N,and P were primarily affected by the soil P supply,especially in tropical forests.Furthermore,after decades of N deposition,subtropical forests can be N-saturated;thus,these ecosystems may shift from being N-limited to P-limited,exhibiting decreasing P contents and increasing N/P ratios(Yu et al.,2018b).
Latitudinal gradients in leaf C,N,and P stoichiometric characteristics indeed exist in natural evergreen broadleaved forests identified in dataset.Similar to previous findings(Li et al.,2015;Hu et al.,2021),this study found that leaf C concentrations decreased with increasing LAT.When LAT decreases,the temperature increases,precipitation decreases,and plants may increase the proportion of C-rich tissues(such as lignin)to adapt to environmental change(Li et al.,2015).In this study,we found that as LAT increased,leaf N and P concentrations increased,and the N/P ratio decreased.This finding supports the temperature-plant physiological hypothesis (Reich et al.,2004),which proposes that leaf N and P concentrations increase from the tropics to cooler and drier mid-latitudes because of temperature-related plant physiological stoichiometry and biogeographical gradients in soil substrate age.The reduced rates of biochemical reactions are caused by the diminished efficiency of N-rich enzymes and P-rich RNA at low temperatures (Reich et al.,2004).Our results are in agreement with the trends identified in some previous studies (Reich et al.,2004;Ren et al.,2007).In addition,leaf P concentrations declined when MAP increased in our study,which is similar to early findings (Chen et al.,2013;Tian et al.,2019).With increase in precipitation,leaching losses of nutrients from leaves and soil increase gradually,which directly decreases leaf P concentrations (Wu et al.,2010).Because of changes in leaf C,N,and P concentrations,leaf C/N,C/P and N/P ratios decreased obviously with increasing latitude,and C/P and N/P ratios increased noticeably with increasing MAP.
Leaf nutrient concentrations are also controlled by many other interrelated factors,including light,annual accumulated temperature>10°C,topography,stand density and increases in biomass production(Tang et al.,2013;Read et al.,2014;Guo et al.,2018).Studies evaluating more factors will help improve our understanding of the complex relationships between leaf stoichiometry and environmental factors.
5.Conclusions
This study explored leaf stoichiometry in various age groups in two forest types in subtropical areas.The results indicated that the leaf C,N,and P stoichiometric characteristics differed among forest stands of different ages in the two forest types.These differences may be caused by changes in litter return and soil nutrients.In Chinese fir forests,the tree growths were more limited by P and N;in natural evergreen broadleaved forests,P limitation increased with stand age.The main driving factors of leaf C,N,and P stoichiometric characteristics in the two forest types were different,which is probably related to the compositions of functional groups in the two forest types.Leaf C concentrations were significantly correlated with GSP and JUT.Leaf N and P concentrations were mainly driven by LNG and SP in Chinese fir forests,which is in consistent with the prediction of the biogeochemical hypothesis.Furthermore,leaf stoichiometric characteristics in natural evergreen broadleaved forests were primarily affected by the combination of LAT and MAP.The normalized regression coefficient indicated larger effect of LAT.Therefore,the changes in leaf stoichiometry support the temperature-plant physiological hypothesis.This study is helpful to improve our understanding of nutrient patterns and the dominant factors affecting leaf stoichiometric patterns in forests with different age groups in subtropical areas.
Authors’contributions
YNC,QLZ,CBX and BYL designed the study,and YNC,QLZ and CBX participated in field work and data collection.YNC,CBX and WPH participated in statistical analysis.YNC,QLZ and CBX prepared the manuscript.QLZ,HY,and BYL contributed to revise the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China(No.31971643),the Industry-University Cooperation Project of Fujian Science and Technology Department (Nos.2020N5008,2019N5009),the General program of Natural Science Foundation of Fujian Province of China (No.2018J01737),Special Funding Project of Fujian Provincial Department of Finance(SC-299)and Minjiang Scholar Programme.
Availability of data and materials
Not applicable.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We thank Bo Hu,Zhi Zhang,Ling Fei,and Leilei Zhang for assistance with the field and laboratory work.We are very grateful to Yong Zheng,Hongbin Xie and Xiaoying Gong(Fujian Normal University)for linguistic revision.
Abbreviations
YF young forest
HF middle-aged forest
NF near-mature forest
MF mature forest
LAT latitude
LON longitude
MAT mean annual temperature
MAP mean annual precipitation
JUT mean July temperature
JAT mean January temperature
RH relative humidity
GSP growing season precipitation
SC soil total C content
SN soil total N content
SP soil total P content

- Forest Ecosystems的其它文章
- Two-level optimization approach to tree-level forest planning
- No treeline shift despite climate change over the last 70 years
- Variation in water supply leads to different responses of tree growth to warming
- Active forest management accelerates carbon storage in plantation forests in Lishui,southern China
- Forest height mapping using inventory and multi-source satellite data over Hunan Province in southern China
- Conifer establishment after the eruption of the Paricutin volcano in central Mexico