
Variation in water supply leads to different responses of tree growth to warming
2022-03-08 02:18PengfeiZhengDndnWngGuodongJiXinxioYuZiqingLiuYusongWngYongeZhng
Forest Ecosystems 2022年1期
Pengfei Zheng ,Dndn Wng ,Guodong Ji,* ,Xinxio Yu,** ,Ziqing Liu ,Yusong Wng ,Yonge Zhng
a Key Laboratory of State Forestry Administration on Soil and Water Conservation,Beijing Forestry University,Beijing,100083,China
b State Key Laboratory of Simulation and Regulation of Water Cycle in River Basin,China Institute of Water Resources and Hydropower Research,Beijing,100038,China
c Co-Innovation Center for Sustainable Forestry in Southern China,Nanjing Forestry University,Nanjing,210037,China
Keywords:Climate change Drought stress Tree rings Stable isotope Snowmelt Temperate forests
ABSTRACT Background: Global climate change,characterized by changes in precipitation,prolonged growing seasons,and warming-induced water deficits,is putting increased pressure on forest ecosystems globally.Understanding the impact of climate change on drought-prone forests is a key objective in assessing forest responses to climate change.Methods:In this study,we assessed tree growth trends and changes in physiological activity under climate change based on measurements of tree ring and stable isotopes.Additionally,structural equation models were used to identify the climate drivers influencing tree growth for the period 1957-2016.Results: We found that the mean basal area increment decreased first and then increased,while the water use efficiency showed a steady increase.The effects of climate warming on tree growth switched from negative to positive in the period 1957-2016.Adequate water supply,especially snowmelt water available in the early critical period,combined with an earlier arrival of the growing season,allowed to be the key to the reversal of the effects of warming on temperature forests.The analysis of structural equation models (SEM) also demonstrated that the growth response of Pinus tabuliformis to the observed temperature increase was closely related to the increase in water availability.Conclusions:Our study indicates that warming is not the direct cause of forest decline,but does indeed exacerbate droughts,which generally cause forest declines.Water availability at the beginning of the growing season might be critical in the adaptation to rising temperatures in Asia.Temperate forests may be better able to withstand rising temperatures if they have sufficient water,with boosted growth even possible during periods of rising temperatures,thus forming stronger carbon sinks.
1.Introduction
Global climate change is putting increasing pressure on forest ecosystems worldwide(Adams et al.,2009;Williams et al.,2010;Devi et al.,2020).Forest productivity depends on various interacting climatic and non-climatic factors,with the main climatic factors including solar radiation,available water,temperature,atmospheric CO2concentration,and nitrogen deposition,among others (Ciais et al.,2005;Allen et al.,2010;Elliott et al.,2015).Among all climatic factors,water deficits are particularly exacerbated by increased warming,and they are one of the most proximal climate threats to forest ecosystems(Poulter et al.,2013;Xu et al.,2020;Zhang et al.,2021).Climate warming can impact forest ecosystems in several ways:extending growing seasons(Piao et al.,2007;Poulter et al.,2013),stimulating photosynthesis (Tumajer et al.,2017),promoting warming-induced water deficits (Restaino et al.,2016),and accelerating snowmelt(Cooper et al.,2020;Zhang et al.,2021).
Most studies have expected an overall negative impact of climate change on forest ecosystems,including declines in forest productivity(Berner et al.,2013;Pellizzari et al.,2016),a reversal of the carbon budget,and even mass dieback events(McDowell et al.,2008,2011;Klos et al.,2009).Climate warming has accelerated widespread growth declines and forest mortality in North America (Peng et al.,2011;D'Orangeville et al.,2016),Europe (Jucker et al.,2017;Tumajer et al.,2017;Albert et al.,2018),and Asia(Poulter et al.,2013;Schaphoff et al.,2016;Zhang et al.,2019).The observed decline in productivity has been inferred to be mainly related to high temperatures and drought stress(McDowell,2011;Dulamsuren et al.,2013).Diverse studies,ranging from those based on greenhouse experiments (Adams et al.,2009),forest-climate change models (Williams et al.,2010,2013),and region-scale forest health monitoring(Littell et al.,2008;Williams et al.,2013),have suggested that warming is causing an increase in the demand for atmospheric water in semi-arid forests.Increased atmospheric water demand may lead to tree hydraulic failure and carbon starvation(Steinkamp and Hickler,2015).
List of abbreviations
SPEI standardized precipitation evapotranspiration index
BAI basal area increment
iWUE intrinsic water use efficiency
VPD saturated vapor pressure difference
RH relative humidity
ETevapotranspiration
However,some studies have found that warming can also have local positive effects in certain regions.For example,in some humid areas of North America,abundant precipitation might help forest ecosystems to better cope with future warming (Cooper et al.,2020;Li et al.,2021).Longer growing seasons associated with warming have also increased forest productivity in some low temperature areas (Clark et al.,2014;Schaphoff et al.,2016).In addition,increased precipitation in winter associated with warming may delay and reduce drought caused by climate warming in northern Eurasia(Ye et al.,2008;Christensen et al.,2021).Hence,it remains unclear whether accelerated warming will increase or decrease radial growth of dominant trees in many regions(Littell et al.,2008;Repo et al.,2021).The nonlinear relationship among tree growth,warming,and warming-induced water deficits increases the uncertainty of predictions (Liu et al.,2013;Shestakova et al.,2017;Zhang et al.,2019).
Parts of inland Asia have warmed relatively rapidly over recent decades,and increasingly frequent and severe drought events have also been observed in central and eastern Asia (Spinoni et al.,2014;Stephenson et al.,2018).Some region-scale studies have monitored the decline in tree growth associated with warming of Inner Asia(Allen et al.,2010;Poulter et al.,2013;Zhang et al.,2019).However,limited studies have been conducted to reveal how warming affects temperate forests in North China (NC) (Sun et al.,2018;Zhang et al.,2021).The potential impact of warming on forests is complex and likely to require consideration of both local and broad-scale factors.
The study of stable carbon isotopes in tree rings has been widely used in research on forest degradation,global environmental change,and tree physiology(Jucker et al.,2017;Sun et al.,2018).In the present study,we assessed tree growth trends and changes in physiological activity under climate change based on patterns in tree rings and stable isotopes.We sought to determine how drought and climate warming have affected tree growth and to identify the climate drivers that control tree growth.Based on that reported for other forests,we assumed that climate warming causes a relative decrease in tree growth owing to drought stress (Allen et al.,2010;Liu et al.,2013).We posit that the potential positive impact of warming alleviates low temperature stress and thereby extends the growing season.Finally,we hypothesize that water supply in all forms,such as snowmelt and growing season precipitation,is a key constraint to tree growth.Additionally,we have the following specific goals:(1) to study the characteristics of regional climate change (temperature,precipitation,and standardized precipitation evapotranspiration index (SPEI)) and the drought stress caused by increased warming;(2) to assess tree growth trends (basal area increment,BAI) and how warming and drought stress affect tree intrinsic water use efficiency(iWUE);(3)to assess the water supply constraint to forest productivity in the context of future climate change.
2.Materials and methods
2.1.Description of sampling points
Our study area is located in the Jundu Mountains northwest of Beijing,which is within a temperate forest zone and has a warm temperate continental monsoon climate(Fig.1).The sampling site is located in the largest naturalPinus tabuliformisforest in North China,with mature trees ranging from 80 to 360 years of age.We selected the nearest meteorological stations to describe the climatic conditions,in Yanqing(115.58°E,40.27°N,487.9 m a.s.l.) and Huailai (115.3°E,40.25°N,570.9 m a.s.l.),which are 13 and 25 km away from our study area,respectively.According to meteorological data,the average annual temperature is about 9.6°C,the average annual precipitation is about 391.9 mm,and the annual frost-free period is 175 days.The main tree species found at the sampling points arePinus tabuliformis,Betula platyphylla,andJuglans mandshurica.We selectedPinus tabuliformisas the study tree species because its tree rings are clearer and wider than those of the other dominant tree species.
2.2.Tree ring collection and processing
Studies assessing tree ring δ13C isotopes show that four trees and four cores collected from a single sampling site can accurately represent the absolute δ13C content and temporal trend of a sampling site(Lu et al.,2019).In this study,10 sample plots (20 m × 20 m) were selected to uniformly cover our study area,and a total of 40 mature trees (average age,94 years) in good growth condition were selected.A total of 80 tree cores with a diameter of 5.15 mm were drilled at a height of 1.3 m in May 2017.In order to prevent carbon source pollution,all samples were stored in glass tubes.Owing to improper operation of the collection process or peculiarities of the trees themselves,some tree cores were of poor quality,and the rings were thus difficult to identify.We selected 38 samples with clear tree rings and fewer missing rings for cross-dating and δ13C analysis.
After the standard treatment of the tree core samples,including natural drying,fixing,grinding,etc.,we used the LINTAB 5 measuring system(Rinntech,Heidelberg,Germany)to measure the tree-ring width(measuring accuracy,0.001 mm).Additionally,all samples were visually cross-dated to avoid issues with missing or false rings.We used the COFECHA procedure for cross-dating the tree core samples and the RCSigFree procedure for inferring the chronology to mitigate the problem of trend distortion (Melvin and Briffa,2008).Finally,we separated the standard tree ring samples according to the tree ring chronology,and samples from the same sample site were combined into a single sample for each year for subsequent carbon isotope analysis.
We homogenized the resulting samples using a ball mill (Retsch,Haan,Germany) and then extracted the α-cellulose according to the method described by Ferrio and Voltas(2005)for stable carbon isotopic analysis.We placed 0.4-0.6 mg of α-cellulose samples per year in tin capsules,and then,an elemental analyzer (Flash EA 1112,Thermo Finnigan,Germany) and a stable isotope ratio mass spectrometer (DELTAplusXP,Thermo Finnigan,Germany) were used to measure the δ13C value with a systematic error of less than 0.2‰.The isotope ratios(13C/12C),indicated with the δ symbol,are presented relative to the Vienna Pee Dee Belemnite standard(for carbon).In order to focus on the effects of climate change on tree growth,we corrected the effect of tree-ring isotopes on changes in atmospheric δ13C values based on the method described by McCarroll and Loader (2004).The atmospheric background δ13C in the correction process was derived from the data published by Shestakova et al.(2017).
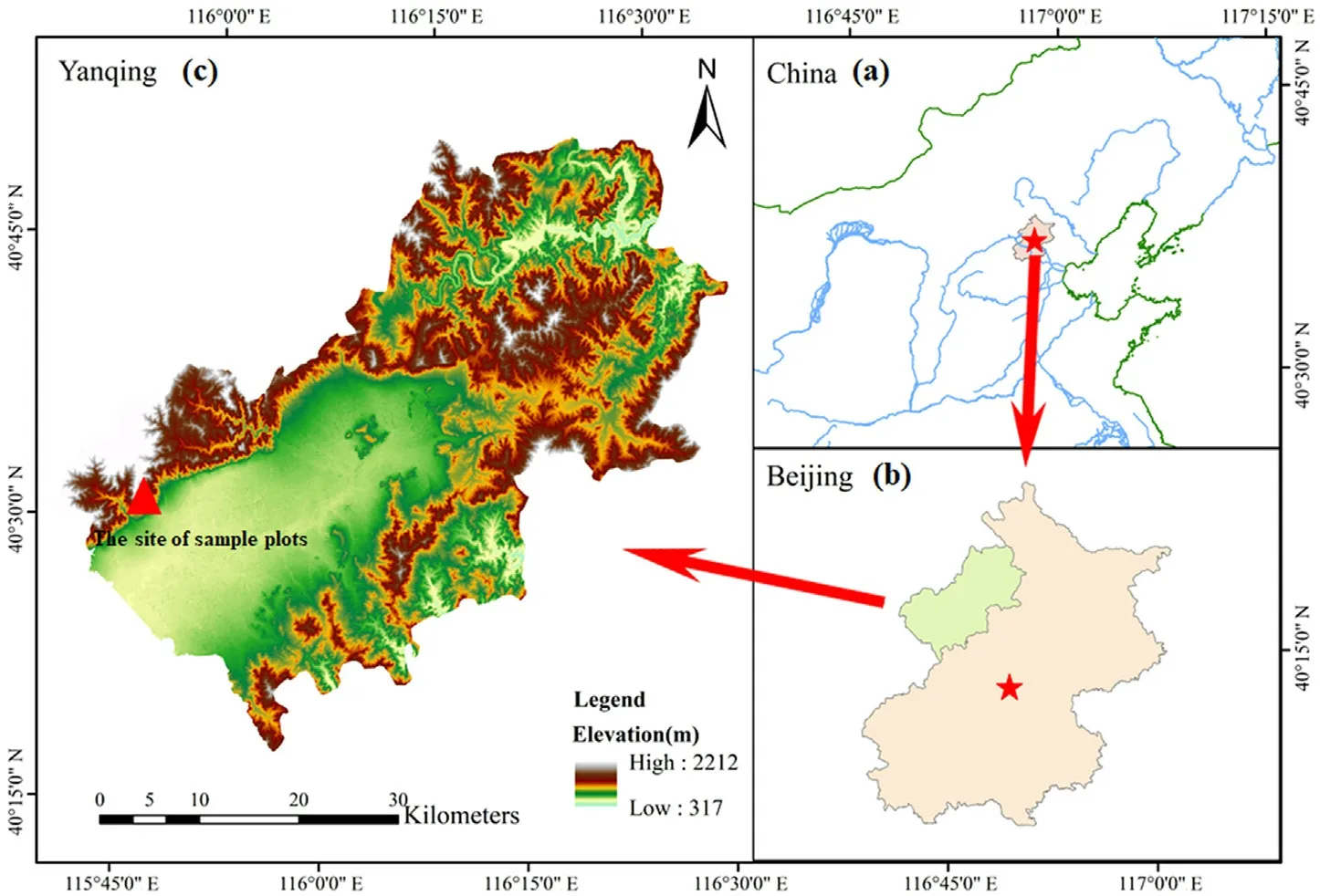
Fig.1.Location map and sampling site of our study area.
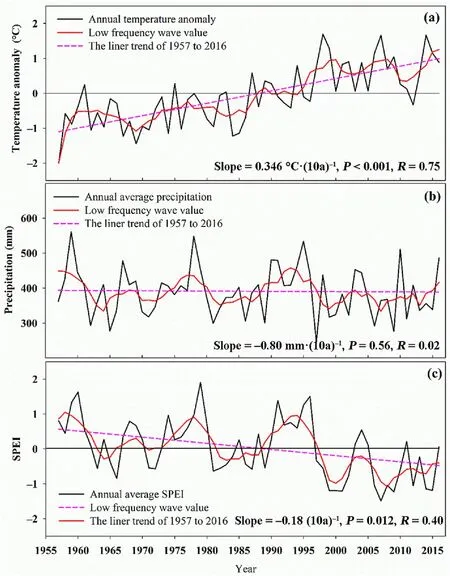
Fig.2.Change in air temperature anomalies (a),annual precipitation (b),and standardized precipitation evapotranspiration index (c) in our study area in 1957-2016.* and • represent the significance test results of P <0.05,and P <0.1,respectively.
2.3.Data acquisition and calculation
Data on monthly temperature,precipitation,atmospheric pressure,and relative humidity (water vapor pressure,relative humidity,etc.)were obtained from the China Meteorological Data Sharing Service System (http://data.cma.cn/).We selected the Yanqing and Huailai meteorological stations for data representative and used the Kendall method and double mass curve(DMC)analysis to test the homogeneity of the meteorological data.Thus,the meteorological data for these sites were determined to be reliable and without aberrations and thus to well represent climate change occurring in the local area.Temperature anomalies and annual precipitation were used to represent climate changes in our study area (Fig.2),where Temperature Anomalies represent the difference between the annual average Temperature and the multi-year average Temperature.We estimated snowmelt data based on potential snowmelt and winter snowfall.The potential snowmelt water data were estimated from temperature and winter precipitation data,according to a simple formula(Zhang et al.,2019):

where,Mis the potential snowmelt,in mm•day-1;Cmis the degree-day coefficient,in mm•degree-day-1;Tais the average daily air temperature(°C);Tbis the base temperature(°C).In the calculations used,Cmwas usually set at 2.74,andTbwas set at 0°C.
We set up soil temperature and soil volumetric water content measurement probes (Decagon 5 TE) at different soil depths (0-20,20-40,40-60 cm) in the sample plot,together with an EM50 data collector,to monitor long-term soil temperature and soil volumetric water content since 2019.
To quantify the severity of the drought,we used station climate data to estimate standardized precipitation evapotranspiration indices(SPEIs)and saturated vapor pressure difference (VPD) (Vicente-Serrano et al.,2010;Li et al.,2021).We used monthly temperature,monthly rainfall,and weather station latitudes to calculate the SPEI change from 1957 to 2016 in R,with regional SPEI data(downloaded from https://spei.csic.es/spei_database) used to validate the SPEI calculations.We used air temperature(Ta)and relative humidity(RH)to estimate changes in VPD within the study area from 1957 to 2016 and used the measured water vapor pressure data from meteorological stations for verification by applying the following equation:
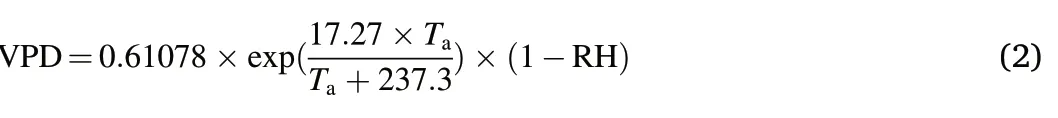
We used daily temperature data to calculate the start and end of the growing season.The beginning of the growing season is defined as a period of five consecutive days with daily temperatures above 5°C,based on the typical value for the onset of wood generation(Rossi et al.,2008;Maxwell et al.,2020),while the end of the growing season is determined using a daily temperature threshold below 0°C.In order to describe the absolute radial growth trend of trees,we converted tree-ring width measurements into basal area increment (BAI) values (Biondi and Qeadan,2008;Mina et al.,2016).We calculated BAI based on a series of tree ring width sequences for intersecting dates:

where,Ris the radius of the tree andtis the year in which the tree rings were formed.Finally,we calculated the mean BAI chronology for each location (Fig.3b).The trend in BAI across two consecutive periods(1957-1987 and 1988-2016) was independently assessed by linear regression,and the switch date was determined by using a Kalman filter.
We estimated intrinsic water use efficiency (iWUE) using the quantitative relationship between δ13C and iWUE determined (Zadworny et al.,2019):

where,δ13C is the stable carbon isotope value of tree-ring cellulose;δ13Catmis the stable carbon isotope value in atmospheric CO2;Δ is the13C discriminant value referring to the difference in isotope levels during photosynthesis between the tree leaf and air;Carepresents the concentration of atmospheric CO2;arepresents the stomatal fractionation coefficient in the diffusion process,which is about 4.4‰;andbrepresents the fractionation coefficient in the carboxylation process of Rubisco and PEP carboxylase,which is about 27‰.Additionally,the coefficient 1.6 represents the ratio of the diffusivity of water vapor to CO2in the air.TheCavalues are from NOAA's Earth System Research Laboratory (http://www.esrl.noaa.gov/).
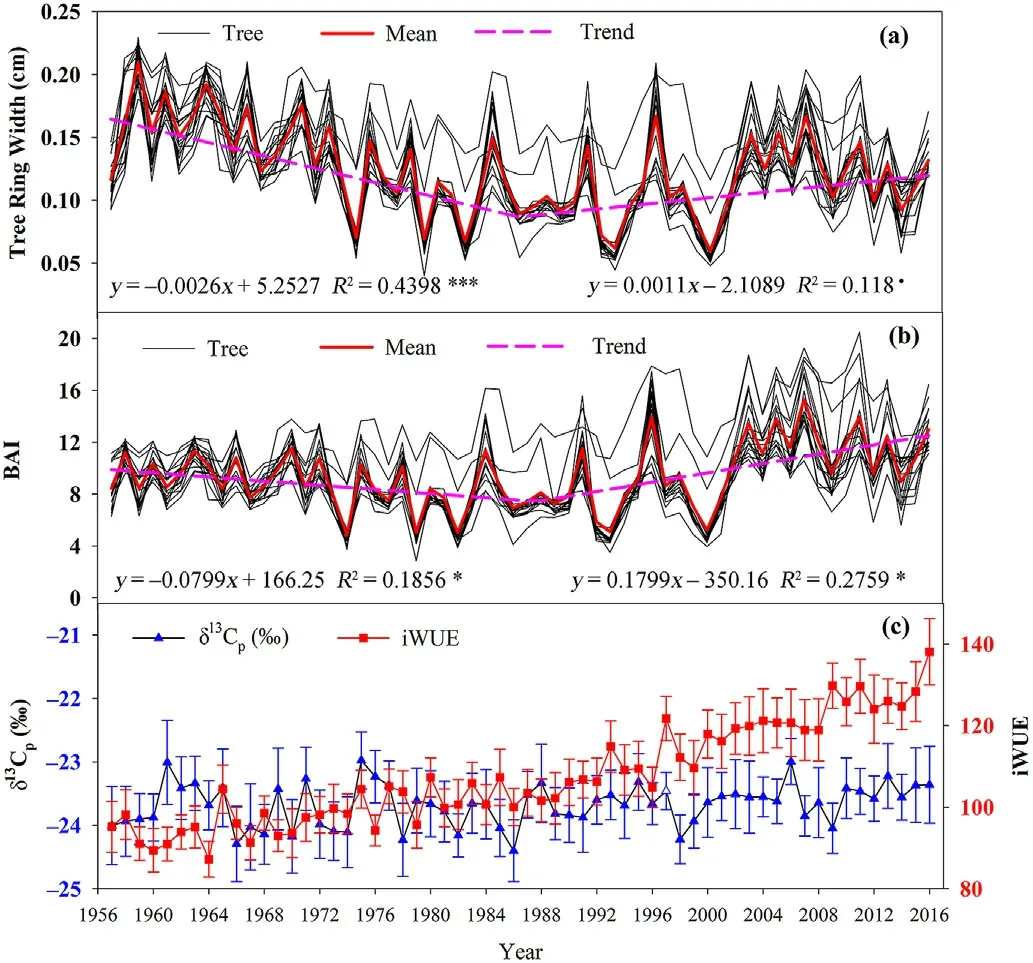
Fig.3.Trends in tree ring width (a),growth of tree sectional area (b),δ13C content of tree rings,and intrinsic water use efficiency (c) during 1957-2016.
We estimated transpiration in our study area from 1957 to 2016 based on annual carbon sequestration and tree iWUE.The annual carbon sequestration estimate was adopted from the biomass model and allometric growth model forPinus tabuliformisin our study area established by Yang et al.(2021).
2.4.Statistical analysis
We determined the time-dependent relationships between tree growth and climate by using Pearson correlation analysis and Kalman filters.We used a sliding window correlation analysis to assess the change in the correlation coefficient between climate factors and site chronology,and we also calculated the correlation between the temperature of each month during the growing season(March to November)and the growth of trees.The climate is generally considered to be the average of meteorological conditions,e.g.,temperature and rainfall,over a 30-year period,as this length of time is considered sufficient to understand the trend in climate change.Accordingly,for our sliding window analyses,we used a fixed window of 30 years,starting with 1957-1986 and ending with 1987-2016,and repeating iterations in oneyear increments(Biondi and Waikul,2008).We also calculated the correlations between tree growth and seasonal (spring,summer,autumn)mean climatic variables,as this is more representative of climatic conditions than data from any single month.
In order to explore the potential interaction between the water cycle and temperature,we calculated the correlations of temperature with precipitation,evapotranspiration,saturated water pressure difference,and water use efficiency (Littell et al.,2008;Poulter et al.,2013;Repo et al.,2021).We also examined how the association between precipitation and growth temperature changed,by using an 11-year time window to assess their consistency over the study period.
Structural equation models(SEM)can be used to assess the effects of multiple climate variables on tree growth and reveal the relative importance of various climate variables on radial growth by inferring covariance among the variables(Grace et al.,2010;Elliott et al.,2015).We used the SPSS Amos V25 (IBM Corp.,Armonk,NY,USA) to build structural equation models for two time periods,1957-1986 and 1987-2016,because the correlation between temperature and water cycle shifted between these two periods.We also examined the relative importance of variables that may affect tree growth using the average BAI of tree rings in our study area,as well as temperature,which was divided into three variables (March-April mean temperature TEM3-4,May-July mean temperature TEM5-7,and August-October mean temperature TEM8-10).We tested models with different variables,recorded their comparative fit index (CFI),normed fit index (NFI),χ2,Akaike information criterion (AIC),and root mean square error approximation(RMSEA) values,and used a final model with optimized values (NFI>0.9,CFI>0.9,P>0.05,and minimized χ2,AIC and RMSEA).
3.Results
3.1.Changes in climate and tree growth in our study area
The region has experienced a period of rapid warming since 1957,and the rate of warming has been 0.35°C per 10 years (Fig.2).The average temperature from 1987 to 2016 was more than 1°C higher than the average temperature from 1957 to 1986.Despite the precipitation not exhibiting a long-term trend,the SPEI showed an obvious downward trend,likely owing to climate warming(Fig.2).The SPEI changed from positive to negative in the period 1957-2016,and the rate of change in SPEI has been 0.18 per 10 years,i.e.,the climate in our study area has faced a trend of warming and drying.

Fig.4.Relationship between tree growth and temperature in different periods organized by month(a and b)as well as season and year(c and d).The vertical line represents the mean Pearson correlation coefficient for each 30-year period,with the different colors representing the different 30-year periods from 1957 to 2016.(For interpretation of the references to color in this figure legend,the reader is referred to the Web version of this article.)
Unlike the constant trends of climate change,the growth of tree ring width and BAI in our study region first decreased and then increased from 1957 to 2016.The inflection point was determined to occur in roughly 1988 by using a Kalman filter approach(Fig.3).The average BAI increased at a rate of 1.799 cm2•(10 a)-1after 1988 (R=0.43,P<0.01),while it decreased at a rate of 0.799 cm2•(10 a)-1before 1988(R=0.66,P<0.01).In parallel with changes in mean temperature,the iWUE ofPinus tabuliformisshowed a significant increasing trend(Fig.3c),increasing by 6.68 μmol•mol-1•(10 a)-1.In particular,the increasing trend was obviously stronger after 1987,reaching 10.29 μmol•mol-1•(10 a)-1.The average iWUE in the period 1987-2016 was 19.5%higher than the average iWUE in the period 1957 to 1986.
3.2.Impact of climate warming on tree growth
The relationship between tree growth and temperature in our study area changed from negative to positive in the period 1957-2016(Fig.4).Correlation coefficients were negative during 1957-1986(mean,-0.34;median,-0.35;standard deviation SD,0.1),while they changed to positive values (mean,0.11;median,0.11;standard deviation,0.04) in the period 1987-2016.Correlation coefficients between tree ring growth and seasonal temperature also changed greatly after 1987,showing a positive influence of temperatures in spring,summer,and autumn(Fig.4c and d).
Warmer temperatures may lead to an earlier arrival of the growing season,with the start of the growing season being 1.5 d•(10 a)-1earlier since 1957 (Fig.5b).The end of the growing season is also delayed(Fig.5a),but this change was not significant.Compared with the growing season start date during the first 5 years of our study period,the start date of the growing season during the last 5 years was 15 days earlier.Thus,the start date of the growing season has advanced from late April up to early April.We found no significant correlation between growing season length and tree growth(P>0.05).
We found that the previously observed negative effect of temperature on tree growth in March and April has been reversed since 2000(Fig.4),which may be related to the earlier growing season being caused by warming.The beginning of the growing season shifted earlier to late March in 2000,and the soil moisture content has been significantly increased by the supply of snowmelt water(Fig.5c).

Fig.5.Change in the end(a)and start(b)of the growing season and change in snowmelt amount in March and April (c) from 1956 to 2016.
3.3.Effects of water supply on tree growth in the growing season
Historical meteorological records show that snowmelt usually begins in late March,with precipitation mainly occurring in the form of snowfall in early March,while snowpack generally disappears in early April(Fig.5c).The time series of snow cover in our region shows that March is the main time in which snowmelt occurs,and there is little snowpack after April.Soil moisture records also show a clear rise in soil moisture from March to April,with the soil temperature also changing from negative to positive during the same period(Fig.6).
We also observed a significant increasing trend in saturated vapor pressure difference (VPD) and evapotranspiration (ET) in the period 1957 to 2016(Fig.7),with VPD increasing by 0.18 hPa•(10 a)-1and ET by 17.68 mm•(10 a)-1.We also found a weakly negative correlation between precipitation and temperature overall,with an average correlation coefficient of-0.1.Additionally,the negative correlation between precipitation and temperature was slightly strengthened in the warming period(Fig.8a).All these factors reflected that the climate in our study area was gradually becoming arid,and the water stress on trees was thus gradually becoming intensified.
There was a positive correlation between temperature and VPD(Fig.8b),and the correlation increased during the warming period(P<0.01).The correlation between temperature and ET was also positive(Fig.8c).However,during the warming period,this correlation weakened (P<0.01) and gradually became non-significant.This indicated that the effect of temperature on ET became weaker as the climate warms,and thus,water supply could gradually become the main factor influencing ET.There was also a positive correlation between temperature and iWUE(Fig.8d),but the positive correlation became a negative correlation after 2000.
3.4.Impacts of climate change on tree growth over time
The climate in our study area exhibited a gradual warming and drying trend (Figs.2 and 7).Water stress on tree growth gradually intensified(Fig.3c),and warming may be the main underlying factor.Thus,between May and July,the negative effects of warming on tree growth were not mitigated,and water supply may be the main factor determining the impact of warming on tree growth.

Fig.6.Changes in soil temperature and moisture across different soil layers from 2019 to 2020.
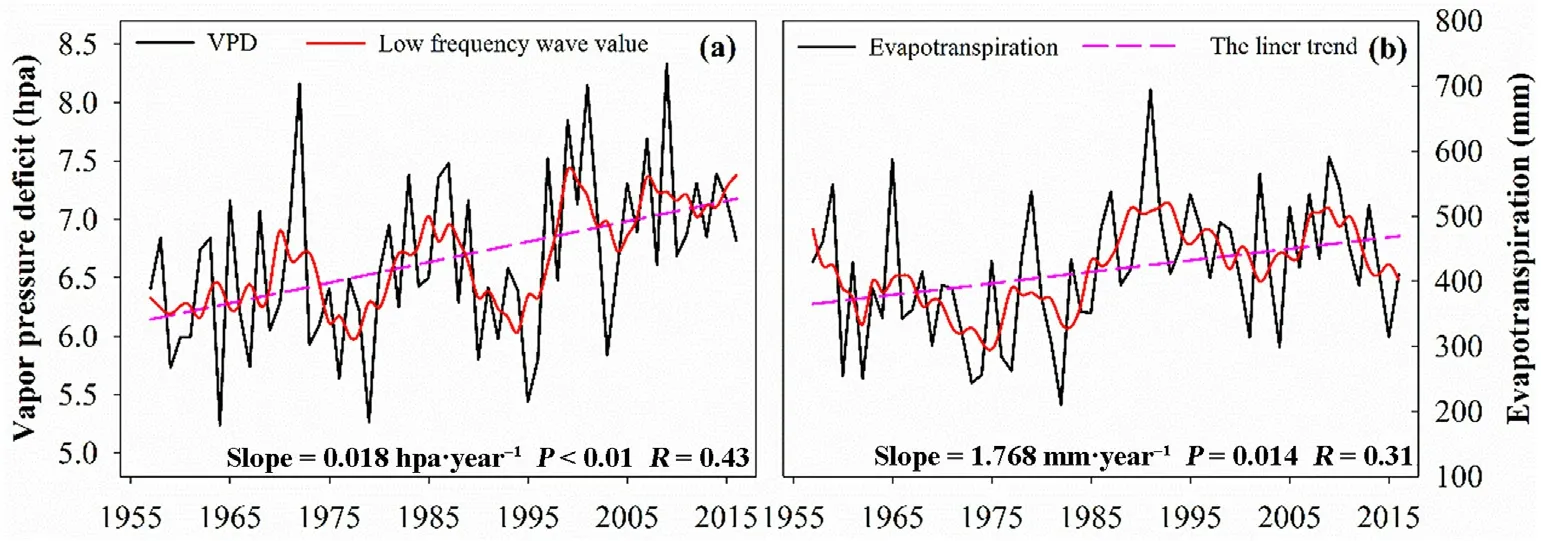
Fig.7.The trend in saturated vapor pressure difference (a) and evapotranspiration (b) in our study area from 1956 to 2016.
To test this hypothesis,we established structural equation models(SEMs) with standard path coefficients for 1957-1986 and 1987-2016(Fig.9),respectively.We found that during 1957-1986,the correlation between warming and summer precipitation was low (R=-0.10,P=0.166),while during 1987-2016,there was a very significant negative correlation between temperature and summer precipitation(R=-0.36,P<0.001)(Fig.9).SEM analysis also indicated the importance of snowmelt water for tree growth during 1987-2016,as the correlation between avera ge temperature in March and Aprilandtree growth changed between the two periods owing to the influence of snowmelt water.
4.Discussion
4.1.Effects of climate warming on forest ecosystems
In this paper,we show that the annual and seasonal correlations between temperature and tree growth changed from negative to positive as the climate warmed over the years.Our findings contrast with those reported that increasing temperatures are often caused by a shift from negative to positive temperature-tree growth relationships (Ciais et al.,2005;Peng et al.,2011).In these studies,temperature rises increase the photosynthetic carbon sequestration of trees,and the extension of growing seasons has also been linked to temperature rises in the early stage of climate warming(D'Orangeville et al.,2016).Collectively,these factors (extended growing seasons and stimulated photosynthesis)strengthen positive relationships between rising temperatures and tree growth in the early stages of warming (Piao et al.,2007;Zhang et al.,2019).However,continued temperature increases exacerbate water deficits (which increases evapotranspiration),leading to drought stress during tree growth (van Mantgem et al.,2009;Restaino et al.,2016;Gradel et al.,2017),ultimately inducing trees to eventually close their stomata and stop growing altogether(McDowell et al.,2008,2011;Choat et al.,2012).
Many studies have suggested that global warming is prone to induce physiological drought (Ciais et al.,2005;Dulamsuren et al.,2013;D'Orangeville et al.,2016).The increased tree iWUE observed in our study suggests that there was an increase in demand for moisture corresponding with warming.Moreover,the reversal of the relationship between tree growth and temperature from positive to negative occurred mainly in the early and late growing season,while the negative effect of temperature on tree growth was always present in the middle of the growing season.Hence,our results are indeed consistent with other studies on the physiological response of trees to warming(Steinkamp and Hickler,2015;Shestakova et al.,2017).

Fig.8.Trend in Pearson correlation coefficients between temperature and water-related factors (i.e.,precipitation,VPD,ET,and iWUE).The blue and red lines indicate trends in the linear regression fit,where the solid lines indicate significant correlations and the dashed lines indicate non-significant correlations.The purple horizontal line represents the average Pearson's correlation coefficient over the following three periods of time:1962-1978,1979-1995,and 1996-2012.(For interpretation of the references to color in this figure legend,the reader is referred to the Web version of this article.)
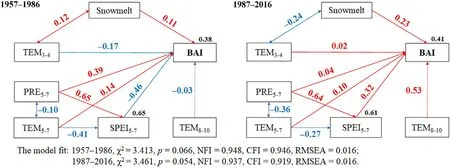
Fig.9.SEM structural equation model for climate variables and radial growth of trees during two different periods (1956-1986 and 1987-2016) with standard path coefficients.Model variables include the snowmelt amount,average temperature from March to April (TEM3-4),average temperature from May to July (TEM5-7),precipitation from May to July(PRE5-7),standardized precipitation evapotranspiration index from May to July (SPEI5-7),average temperature from August to October(TEM8-10),and basal area increment of trees (BAI).
While the growing season has been advancing progressively over the entire study period,there was limited direct influence of extended growing season on tree growth during the period of this study.It is understood that warming intensifies the water deficit of the atmosphere and often intensifies climate aridity in some area(Mcdowell et al.,2011;Dulamsuren et al.,2013;Liu et al.,2013).In our study,a negative correlation between temperature and precipitation was also confirmed,indicating that long-term warming trends promoted the drying of the climate to some extent.In areas where the water supply is limited,insufficient water supply tends to result in hydraulic failure of trees (McDowell,2011;Mcdowell et al.,2011).Drought stress caused by warming has been suggested to be the main driving force behind general increases in tree mortality in arid regions (Berner et al.,2013;Liu et al.,2013;Jucker et al.,2017).
4.2.The importance of snowmelt water for tree growth
The growth ofPinus tabuliformisbegins in early April(Rossi et al.,2008;Seo et al.,2011).However,the precipitation in our study area is very low in April,averaging only 15.5 mm.We also found that before 1986,the precipitation accumulation was very low in winter and mainly concentrated in summer.The average winter precipitation accumulation during 1957-1986 was only 9.1 mm.Combined with the early melting of snow,the water supply to support the growth of wood in the early part of the growing season may be limited(Tognetti et al.,2019;Repo et al.,2021).In contrast,during 1987-2016,the average precipitation accumulation in winter was 26.7 mm,and the warming temperature was associated with an increase in the proportion of precipitation falling in winter.However,some studies have found that warmer temperatures do not lead to earlier snowmelt,increased precipitation in winter leads to increased snow cover change,and deeper snowpack can require greater energy input to overcome cold content and liquid water holding capacity and initiate snowmelt(Musselman et al.,2017;Martin et al.,2018).The interaction between the lengthening of the growing season and the presence of snow in April makes snowmelt an important source of water supporting the growth of trees in the early growing season (Vellend et al.,2017;Reinmann et al.,2019;Zhang et al.,2019;Li et al.,2021).
In some temperate forests,snow melt rings often appear around tree trunks,and a similar phenomenon has been observed in our study area(Vellend et al.,2017).Snowmelt has become an important water resource for trees in the early growing season of forests (Littell et al.,2008;Martin et al.,2018;Repo et al.,2021).In the early stage of tree growth,the physiological activities of trees begin to increase their demand for water,and trees are then particularly vulnerable to water stress.Higher soil water supply caused by snowmelt water can offset the negative influences of VPD on turgor pressure(Zhang et al.,2019;Cooper et al.,2020),which is crucial to counterbalance the drought stress caused by warming and promote a positive relationship between temperature in the early growing season and tree growth.Water is also an important source of carbohydrates for photosynthesis in plants,where carbohydrates produced and stored early in the growing season can then be used to overcome nutrient deficiencies,cold,or drought(Palacio et al.,2014;Dietrich and Kahmen,2019).
4.3.Water supply is a key factor in tree growth responses to warming
Our results demonstrate that water supply is a key factor shaping tree growth responses to warming.Contrary to the shift in the temperature-tree growth relationship in March and April,we found that the temperature-tree growth correlation from May to July did not change over the course of the historical data.This is linked to the strengthened increasing VPD and decreasing rainfall.Shestakova et al.(2017) also found a similar response to warming in tree growth under different rainfall conditions.They found that the growth of the Siberian cold-dry forest increased in areas with rainfall greater than 160 mm in May and decreased in drier areas of the study range.Similarly,Liu et al.(2013)found that in northeastern China,the BAI increment of trees decreased in areas with an average annual precipitation accumulation of 200-400 mm,while tree growth increased in areas with an average annual precipitation accumulation of 400-700 mm.Zhang et al.(2019)also found thatPinus sylvestrispopulations subject to heavy rainfall (201-265 mm)from May to July responded more positively to temperature increases than the drier parts of the study area (145-160 mm absolute precipitation accumulation from May to July).Another analysis of long-term climatological data showed that climate warming in China caused an increase in actual annual evapotranspiration from 1960 to 2002 in arid regions,whereas this was not found in subhumid regions (Gao et al.,2007).All these suggests that water supply is a key factor in tree growth responses to warming,and if water availability is sufficient,future warming is likely to promote plant growth and forest expansion in such regions(Rotenberg and Yakir,2010;Berner et al.,2013).
In our study,we have identified interaction among climate warming,shifting growing seasons,water supply in all forms,and the growth response of trees in a temperate forest system.Owing to the earlier growing season,snowmelt water in the beginning of the growing season became an important water source supporting tree growth.These results contrast with those from other temperate forest regions,where climate warming has often led to widespread forest degradation (van Mantgem et al.,2009;Restaino et al.,2016).The supply of snowmelt water and the earlier arrival of the growing season conditions in our study region jointly led to a shift from negative to positive temperature-tree growth relationships.
Our study suggests that warming is not directly the cause of forest decline,but water deficits were exacerbated by increasing temperature.Global warming increases the sensitivity of semi-arid forests to drought,which is manifested by decreased forest growth and tree mortality caused by warming (Williams et al.,2010,2013;Peng et al.,2011).Continued warming led to shorter periods of drought (18.7 weeks) and longer durations in inland Asia,and it was sufficient to promote widespread forest declines and tree mortality(Adams et al.,2009).Increasing precipitation can decrease water stress to some extent,but on seasonal and longer time scales,increasing evaporation demand caused by warming may exceed precipitation input,leading to water stress in forest ecosystems (Adams et al.,2009;Williams et al.,2013).The intensification of drought caused by continued warming will not exclusively lead to declines in forest growth and mass dieback events in semi-arid regions,and water availability at the beginning of the growing season might be critical in the adaptation to rising temperatures in Asia(Farooqi et al.,2021).
5.Conclusion
In this study,we used tree-ring chronology and stable isotopes to assess the response of tree growth to climate change.We found that the effects of climate warming on tree growth transitioned from negative to positive from 1957 to 2016.Adequate water supply during the growing season,especially snowmelt water available in the early part of the growing season,combined with an earlier arrival of the growing season,appeared to be the key to the reversal of the climate sensitivity of trees in our study area.Water supply in all forms has led to different responses of tree growth to warming throughout the growing seasons.Our study suggests that warming is not the direct cause of forest decline,but does indeed exacerbate droughts,which generally causes forest decline.SEM analysis also demonstrated that the growth response ofPinus tabuliformisto the observed temperature increase was closely related to the increase in water availability.Additionally,the influence of drought on tree growth decreased owing to a compensating effect of a strengthening relationship between precipitation and radial growth.Thus,temperate forests may be better able to withstand rising temperatures if they have sufficient water,with boosted growth even possible during periods of rising temperatures.However,in semi-arid regions where water supplies are limited,continued warming could lead to reduced forest growth or even mass dieback events,and water availability at the beginning of the growing season might be critical in the response of forests to rising temperatures in Asia.
Funding
This research was supported by the National Natural Science Foundation of China(Grant No.41877152),the Fundamental Research Funds for the Central Universities(2019ZY35) and the Beijing Municipal Education Commission (CEFF_PXM2019_014207_000099).
Availability of data and materials
Available on request.
Authors' contributions
ZP,JG and YX planned and designed the research.ZP performed experiments and conducted fieldwork with the help of WY.ZP analyzed the data and wrote the manuscript under the guidance of JG and ZY.WD led the compilation of data compilation and significantly contributed to the analysis of data and its interpretation and rewrote the final version of manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not Applicable.
Competing interests
The authors declare that they have no competing interests.
Author details
1Key Laboratory of State Forestry Administration on Soil and Water Conservation,Beijing Forestry University,Beijing 100083,China.2State Key Laboratory of Simulation and Regulation of Water Cycle in River Basin,China Institute of Water Resources and Hydropower Research,Beijing 100038,China.3Co-Innovation Center for Sustainable Forestry in Southern China,Nanjing Forestry University,Nanjing,210037,China.
Acknowledgements
We thank Erdie Zi and Weiwei Lu for their help with fieldwork.

- Forest Ecosystems的其它文章
- Two-level optimization approach to tree-level forest planning
- No treeline shift despite climate change over the last 70 years
- Active forest management accelerates carbon storage in plantation forests in Lishui,southern China
- Patterns and driving factors of leaf C,N,and P stoichiometry in two forest types with different stand ages in a mid-subtropical zone
- Forest height mapping using inventory and multi-source satellite data over Hunan Province in southern China
- Conifer establishment after the eruption of the Paricutin volcano in central Mexico