
植物中过表达MT 转录因子调控镉胁迫 响应的Meta 分析
2022-02-21 09:33黄亚群
河北农业大学学报 2022年1期
张 威,杜 婕,黄亚群
(河北农业大学 农学院/国家玉米改良中心河北分中心,河北 保定 071000)
已有的研究显示,金属硫蛋白(Metallothionein,MT)能有效的缓解生物重金属离子的毒害[4]。MT蛋白是一种富含半胱氨酸(Cys)低分子量的多肽[5]。其半胱氨酸残基上的巯基对于金属元素有很高的亲和力,尤其是对铜(Cu)、镉(Cd)、锌(Zn)和铅(Pb)等[6]。MT 蛋白一种作用方式是,与重金属离子结合形成复合体,通过溶酶体从细胞质排到细胞外,减轻重金属对生物的不良影响[7];另一种作用方式是,在生物体内维持金属离子的稳态平衡,在金属离子较低时提供金属离子,并将金属离子输送到所需的组织;当金属离子含量提高时,又可以结合超量的金属离子[7-8],或结合非必需金属离子,来替换重金属离子,从而消除其毒性[8]。
MT 蛋白广泛存在于动物、植物和微生物中。根据蛋白质结构不同,分为Ⅰ、Ⅱ和Ⅲ3 种类型。其中,植物MT 蛋白大多数为Ⅱ型,依据半胱氨酸(Cys)的含量和分布,又分为MT1、MT2、MT3、MT4 4种亚型[7]。不同的MT 蛋白结合金属离子能力大小存在着差异[9-10]。有关转MT 基因在镉胁迫和非胁迫条件下的表现存在不一致的看法。一类研究结果显示,在非镉胁迫下,植物过表达MT 个体较其野生型体内的谷胱甘肽和半胱氨酸的含量有显著增加[11],总叶绿素含量增多[6],H2O2含量降低、细胞膜系统的损伤减少[12];在镉胁迫下,植物转MT个体较其野生型对照对不同金属离子的耐受性有显著提高[13-16],受伤害后能更快的得到恢复[7],叶绿素含量减少的幅度要低于野生型个体[6]。另外一类研究结果显示,在重金属镉胁迫条件下,MT 转录因子在叶绿素含量方面发挥负调控作用[17];对转PpMT2基因拟南芥叶片化学染色检测,发现在正常处理条件下H2O2含量要高于其对照野生型,而在非镉胁迫下,转基因拟南芥H2O2含量更低[18]。为了解决单个研究材料、方法以及胁迫处理时间和程度等众多限制因素的不同,导致研究结果表现出不同程度的差异,本研究利用Meta 分析方法,将多个独立研究数据进行整合,定量评价过表达MT 转录因子在调控镉胁迫过程中的作用,以期为进一步培育耐(富集)镉植物品种提供依据。
1 方法
1.1 文献收集
利 用EndNote X7 软 件 从PubMed、Web of science,中国知网、万方 4 个数据库中,以“作物(Crop)/植物(Plant)”、“金属硫蛋白(Metallothionein, MT)”、“重金属胁迫(Heavy metal tolerance)”、“镉胁迫(Cadmium, Cd)”为关键词对发表在2010—2020 年间文献进行检索,共检索到了458 篇。
对上述检索到的文献再按以下标准进行筛选:(1)剔除从不同数据库中检索到的相同研究;(2)至少应包括一个表型评价指标(如, 根长(Root length, RL)、丙二醛含量(Malondialdehyde content, MDA)、 过 氧 化 氢 含 量(H2O2content, H2O2)、鲜 重 (Fresh weight, FW)、干 重(Dry weight, DW)、叶 绿 素 含 量(Chlorophyll content, Chl));(3)剔除综述、会议报告等非研究型文章;(4)每一个研究应包含对照组和处理组,即包含植物野生型和转基因型个体在非胁迫和胁迫下的试验;(5)每一个处理应至少包含3 个重复;(6)若同一篇文献中重金属胁迫程度不同时,将视为多个独立的研究结果。
符合上述标准的文献共16 篇见文献[7、14、16、18 ~30](9 篇英文文献和7 篇中文文献),87 个相对独立的研究数据。在16 篇文献中,有 9篇以烟草(Tabacoo)、6 篇以拟南芥(Arabidopsis)、1 篇以水稻(Oryza sativa)作为受体植物;供体植物有 木 豆(Pigeonpea)、菜 豆(Phaseolusvulgaris),水稻,柽柳(Tamarix androssowii)、巴西橡胶树(Hevea brasiliensis)、芹 菜(Apiumgraveolens)、小 立 碗 藓(Physcomitrella patens)等。
1.2 数据提取
如果同一文献包含多个不同的处理方法,将视为多个独立的研究纳入分析[31]。为了区分涉及同一作者的多个独立研究,采用Author + year +英文字母后缀进行区分。将提取数据按作者及文献发表时间、题目、期刊名称、转录因子类型、供体植物、受体植物等格式整理。
从已纳入的文献中提取对照组和处理组某一性状指标的均值(X)、样本量(n)和标准差(SD) 3 个统计量用于后续的分析。若文献中数据是以表格呈现,则直接提取符合要求的数值;若表中的数是多个重复值,则计算出平均值后再纳入。若文献中数据是以图形呈现,则利用GetData2.26 软件进行提取。若文献中缺少样本容量信息,则按样本容量为1 处理。若研究中以标准误(SE)表示变异的,则利用公式(1)转换为标准差(SD)。
高潮发短信的时候,田卓就坐在那里静静地等候着。发完短信,高潮一抬头,看到田卓在等他,赶紧冲她歉意的一笑。

为了避免不同研究采用的测量单位和尺度上的差异,将在原始文献中镉胁迫或非镉胁迫条件下转基因个体及其野生型的绝对值转换为响应比(R)。在Meta 分析中,效应大小采用响应比的自然对数(lnR)表述,见式(2)。

XA代表转基因植物在重金属胁迫条件下(或MT 过表达转基因)个体的表型值,XB代表非胁迫条件下(或野生型植物)个体的表型值。
1.3 Meta 分析统计方法
采用Q检验对纳入的原始研究进行异质性分析。对异质性较大的研究进一步进行亚组分析,以减少研究间的异质性。Q统计量为标准化效应量的平方和(式(3))。该统计量常用于评价异质性,但在研究数据较少的情况下,该检测的有效性较差。I2为估计的观测效果大小的真异质性与总异质性之比 (公式(4))。当异质性Q检验(卡方检验)的p值大于0.1时,认为同质性假设有效;否则,拒绝同质性假设,即认为研究间存在着异质性。
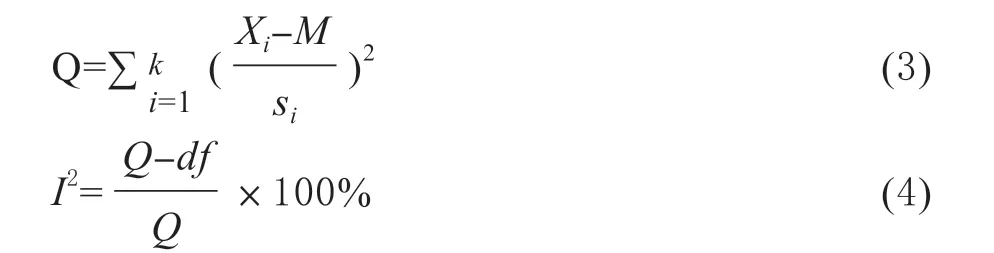
(3)式中,Xi为每个独立研究的效应值,M为所有研究的效应值的均值,Si为每个独立研究的标准差,df为自由度(纳入研究的数量n减1)。式(4)中I2值为0% ~ 100%。若实际计算出I2值为负值,则视为0%。统计学上定义:I2值越大,异质性越大;若I2=0%,则认为各研究之间的差异是因抽样误差所致;若I2>0%,即认为各研究之间存在着异质性,通常将I2> 50%,认定异质性存在。将I2等于25%、50%和75%分位数上的值分别代表异质性的低、中、高程度界限[32]。
用变化率(Change rate)表示转基因个体及野生型个体之间的差异比率,计算公式如式(5)Change rate=(1-e(lnX))×100% (5)
采用R 统计软件3.1 版( 混合效应模型) 的Nlum 软件包计算性状指标变异区间。如果森林图中每个组合效应值的95%置信区间与中心轴0 相交(即森林图中每个独立的水平线与垂直线相交),则认为试验组与对照组的效应值差异未达到显著水平;相反,如果没有交集,则试验组与对照组的效应值有显著差异(P< 0.05)。当小点位于中心轴左侧时,表明MT 转录因子对性状指标起负调控作用,当小点位于中心轴右侧时,则表示正向调控作用。越靠近中轴,MT 转录因子在性状指标中的作用越不显著。
2 结果与分析
2.1 野生型及过表达MT 转基因植物对镉胁迫的响应
植物野生型个体在正常(非镉胁迫)与胁迫条件下,在根长、过氧化氢含量、植物干重3 个性状上存在显著差异(图1a)。其中,镉胁迫对植物根长和干重呈负向影响,变化率分别为60.67%、98.07%;而对野生型个体过氧化氢含量呈现正向影响,变化率为423.40%(图1a)。转基因个体在镉胁迫下,显著降低个体的干重,变化率为97.88%(图1b)。
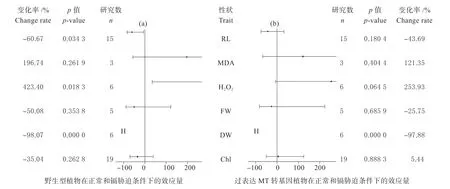
图1 野生型和过表达MT 转基因型植物在不同条件下效应值(lnR)Fig.1 Effect value of wild-type and overexpressed MT transgenic plants under different conditions(lnR)
2.2 同一条件下MT 过表达植物与其野生型的表现
在正常条件下,转基因植物与其野生型在根长、丙二醛含量等6 个性状指标表现均无显著差异(图2a)。
在镉胁迫条件下,转基因植物与其野生型在根长、过氧化氢含量、植物鲜重、叶绿素含量4 个性状指标上表现出显著差异。其中,MT 转录因子对过氧化氢含量发挥负向调控作用,转基因植株中H2O2含量比野生型低29.5%。在根长、植物鲜重、叶绿素含量这3 个指标中,MT 转录因子发挥正向调控作用,其中发挥正向调控作用最大的是叶绿素含量,转基因型植株中叶绿素含量要比其野生型个体高59.12%(图2b)。
2.3 异质性分析
由表1 看出,在正常条件下,无论是转基因植株还是其野生型植株,在干重和叶绿素含量2 个表型性状呈显著性差异。因干重仅纳入3 个研究数据,样本量小,不需再做进一步分析[20]。叶绿素含量纳入了9 个研究信息(图2a),且各研究间存在较大异质性,需要对其进行亚组分析。
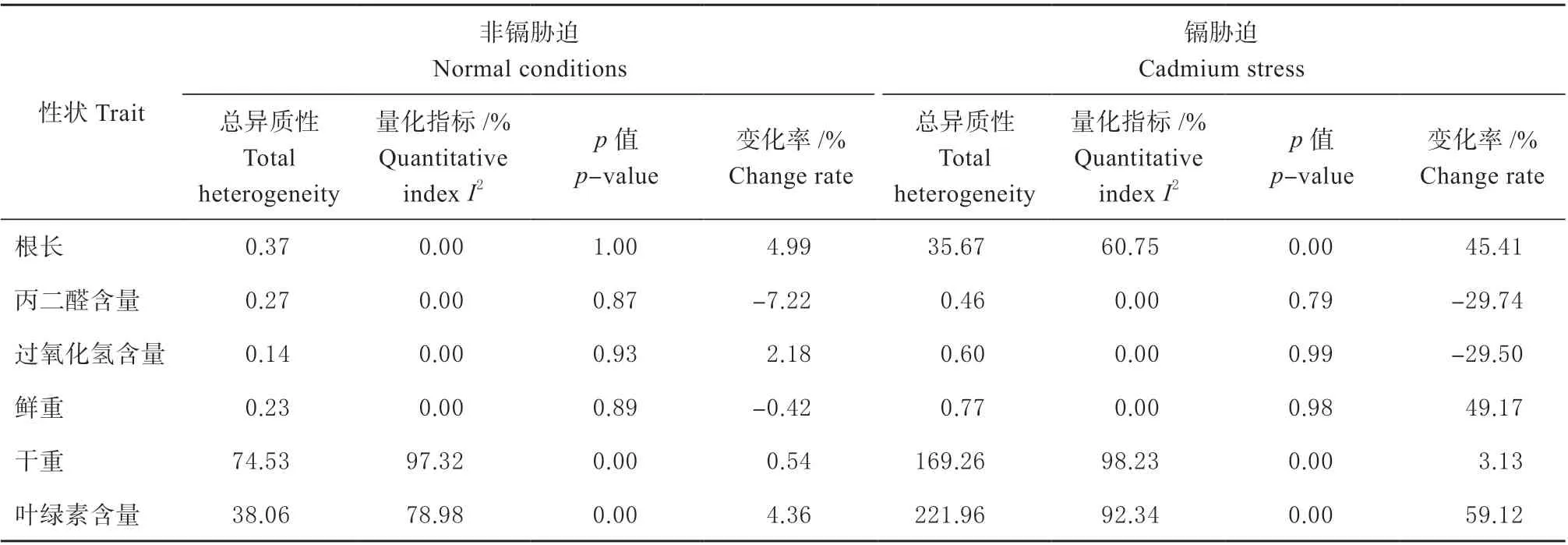
表1 非镉胁和镉胁迫迫条件下6 个表型性状异质性分析Table 1 Heterogeneity analysis of 6 phenotypic traits under normal conditions and cadmium stress
镉胁迫条件下,根长、干重、叶绿素含量3 个表型性状呈显著性差异,因干重只纳入了4 个研究的数据,认为样本个数少,故不再做进一步的亚组间异质性分析。根长和叶绿素含量分别纳入15 个和18 个研究数目(图2b),且2 个性状的I2 均大于50%,存在较大异质性,根长变化率为4.99%,叶绿素含量变化率为59.12%。因此,需要对根长、叶绿素含量做亚组分析。
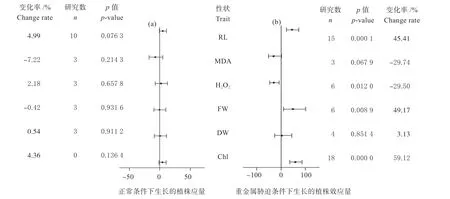
图2 相同处理条件下转基因型植物与野生型加权效应值Fig.2 Weighted effect values of transgenic plants and wild types under the same treatment conditions
2.4 亚组分析
2.4.1 镉胁迫下植物根长的亚组分析 对镉胁迫下根长在培养基质、胁迫时间、胁迫程度、受体植物、受体类型5 个变量进行亚组分析,发现MT 转录因子对根长起正向调控作用。
当培养基质为MS(或1/2 MS)培养基时,转基因植物的根长与其野生型呈显著差异,变化率为89.09%(或55.18%)(图3a)。
胁迫时间在1 ~15 d 内,转基因植物根长与其野生型根长呈极显著差异(P<0.01),变化率为55.32%;当胁迫时间在15 ~30 d 内,转基因植物和野生型植物根长呈现显著差异,变化率为68.74%,随着胁迫时间的延长,MT 转录因子对根长的调控作用逐渐增强(图3b)。胁迫程度较低时,呈现极显著差异(P<0.01),变化率为66.55%;当胁迫程度中等或较高时,虽没有呈现显著性差异,但转基因植物的根长要高于其野生型(图3c)。受体类型为双子叶植物时,均呈现显著性差异,变化率分别为70.17%(图3d)。受体植物类型为烟草、拟南芥时,转基因植物个体与其野生型个体的根长呈显著差异,变化率分别为175.35%、59.89%,当受体植物为烟草时,转基因植物与野生型植物的根长相差最大(图3e)。
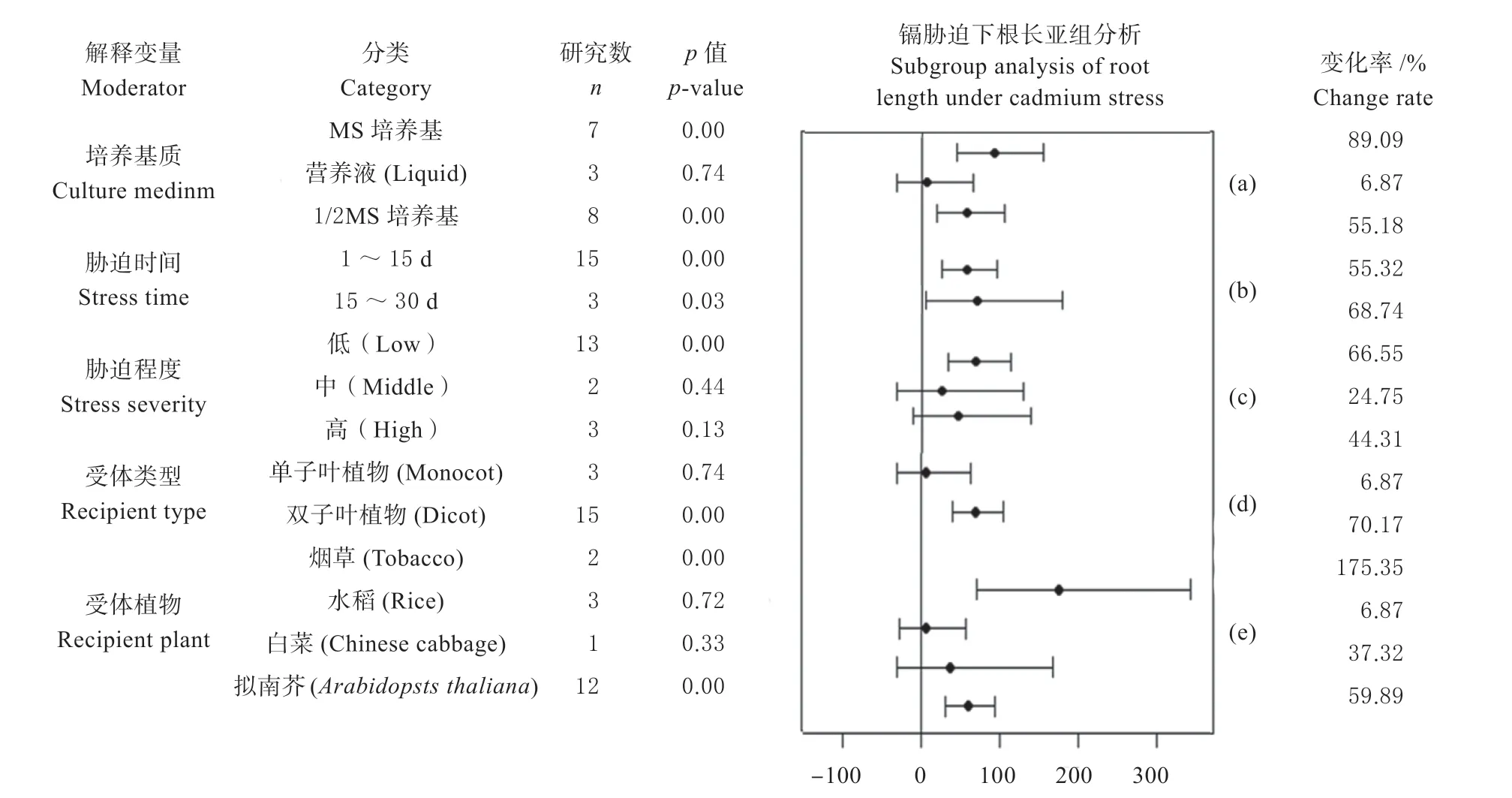
图3 镉胁迫条件下MT 转录因子对植物根长表型指标影响的效应值及其95%的置信区间Fig.3 Effect values and 95% confidence intervals of MT transcription factors on plant root length phenotypic indicators under cadmium stress
2.4.2 不同条件下植物叶绿素含量亚组分析 正常条件下,对受体植物、培养时间、培养基质、供受体类型等6 个变量进行亚组分析,发现只在MS 培养基这1 个解释变量下,植物转基因植株与野生型个体在叶绿素含量上存在极显著差异(P<0.01),转基因植株叶绿素含量比野生型植物叶绿素含量高12.52%(图4-1c)。
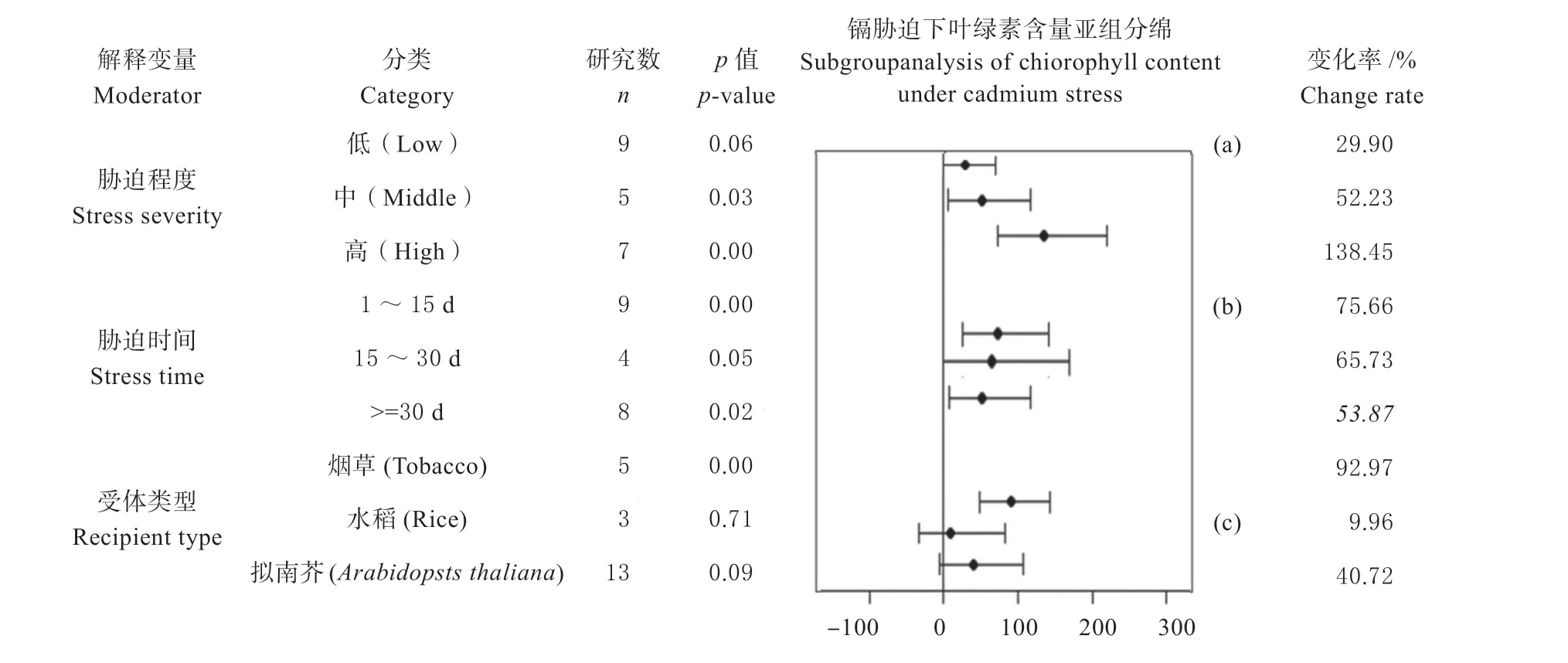
图4 非镉胁迫条件下MT 转录因子对植物叶绿素含量表型指标影响的效应值及其95%的置信区间Fig.4 Effect values and 95% confidence intervals of MT transcription factors on plant chlorophyll content phenotypic indicators under non-cadmium stress
镉胁迫条件下,对胁迫程度、胁迫时间、受体植物类型等变量进行亚组分析,发现叶绿素含量在中度胁迫、高度胁迫、胁迫时间在1 ~5 d、大于等于30 d、受体类型为烟草这5 个指标上呈显著性差异(P<0.05),且均为正向调控,其中高度胁迫下,野生型个体与其转基因植株的叶绿素含量差异最大,变化率为138.45%(图4-2a);且2 种类型个体中的叶绿素含量会随着胁迫程度增强达到极显著差异,变化率由29.90%到138.45%(图4-2a)。从胁迫时间来看,镉胁迫在1 ~15 d,转基因植株与其野生型个体的叶绿素含量呈极显著差异(P<0.01),变化率为75.66%,当胁迫时间到15 ~30 d,转基因植物与野生型植物的叶绿素含量无差异,当胁迫时间大于等于30 d 时,转基因植物与野生型植物的叶绿素含量呈显著差异(P<0.05),变化率为53.87%(图4-2b)。当受体植物为烟草时,转基因植物与野生型植物的叶绿素含量呈极显著差异(P<0.01),变化率为92.97%(图4-2c)。
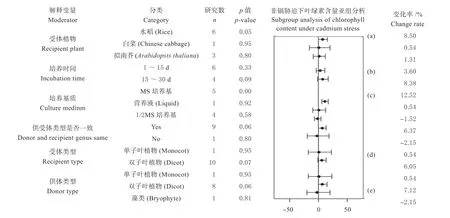
图5 镉胁迫条件下MT 转录因子对植物叶绿素含量表型指标影响的效应值及其95%的置信区间Fig.5 Effect values and 95% confidence intervals of MT transcription factors on chlorophyll content phenotypic indexes of plants under cadmium stress
3 讨论
3.1 镉胁迫对植物生长特征性状的影响
已有研究表明,在镉胁迫下,过氧化物含量明显升高,过氧化过程产生新的自由基,膜的完整性受到破坏[33],植物根长显著缩短,干重显著变轻[34]。植物体中镉的持续积累,会更进一步影响光反应和暗反应的进程以及叶片气孔的闭合[35],降低叶片叶绿素含量,降低电导率[36],抑制碳同化,加速叶片衰老,破坏植物光合作用,最终导致光合效率下降,影响作物的产量与品质[37]。本文Meta 分析结果同样显示,无论是野生型还是转MT 基因植株在镉胁迫下其根长、叶绿素含量和植株干重均显著降低,而过氧化氢含量显著增加。
3.2 MT 基因对植物生长特征性状的影响
崔广娟[38]、王利芬[39]等人通过对豆科、菊科植物进行镉胁迫试验表明植物在镉胁迫下总根长和根表面积随着 Cd 浓度的增加逐渐下降, Meta 分析也获得了相同的研究结果。本研究发现,野生型植物受到镉胁迫后,与其对照组相比根长的长度显著缩短;而转基因植物在胁迫前后的根长没有明显差异。在正常条件下,野生型个体的根长与转基因植株的根长没有表现出显著差异;当受到镉胁迫时,野生型的根长要显著低于转基因植株。本研究亚组分析发现,MT 基因对根长呈正向调控作用,致使转基因植株的根长受镉胁迫影响较小,从而增强了该植物对镉的抗性,保证其正常生长。
H2O2是植物长期处于非生物胁迫的条件下产生的一种有害的生物化学物质。杜佳[6]通过研究柽柳MT3基因在拟南芥中的功能发现,在未经镉胁迫时转基因拟南芥可以降低植物体内H2O2含量,减少细胞膜系统的损伤。Liu[18]研究发现转基因拟南芥叶片在正常处理条件下H2O2含量要高于其对照野生型,而在镉胁迫下与野生型相比,转基因拟南芥具有更好的渗透保护作用和较低的脂质过氧化作用,这与本文研究结果相一致。Meta 分析发现野生型植株H2O2含量在镉胁迫前后有显著增加,而转基因植株H2O2含量增加并不显著;在正常条件下,野生型与转基因植株的H2O2含量无显著差异;而在镉胁迫条件下,野生型植株的H2O2含量与转基因呈显著性差异,且野生型个体内积累的过氧化物含量要高于转基因植株,转基因植株表现出较强的抗H2O2伤害的能力。
3.3 影响MT 过表达调控叶绿素含量效果的因素
已有的报道显示,MT 基因对调控叶绿素含量的作用上有不一致的看法。Kumar[3]、Balasundaram[28]等人认为,MT 具有清除活性氧自由基的功能,镉胁迫下转MT 基因烟草叶绿素含量明显高于野生型对照,过表达MT 转基因型植物的叶绿素含量减少的幅度要低于野生型对照。但是,范仲学等[29]试验发现,在CdCl2浓度为400 μmol/L 时,SorMT2c转基因烟草的叶绿素含量要明显低于野生型;Rono等[30]试验发现在10 μmol/LCd 处理下OsMT1e转基因水稻叶片的叶绿素含量低于野生型。本文通过Meta 分析发现,在镉胁迫程度、胁迫时间、受体类型不一致的情况下,转MT 基因的植株与其野生型对照叶绿素含量存在显著差异。具体表现为:(1)在镉胁迫为高浓度、受体植物为烟草时,叶绿素含量受胁迫时间的影响。当胁迫时间为1 ~15 d,转MT 植物叶绿素含量显著高于野生型对照;当胁迫时间超过30 d,转MT 植物与其野生型对照的叶绿素含量间差异不显著。MT 基因在植物镉胁迫小于15 d 时对叶绿素含量的调控作用更强,随着镉胁迫时间的延长,调控效果减弱,但要比野生型植物叶绿素含量高,在一定程度上起到维持叶绿素含量的作用,从而提高植物光合作用,保证了植物的产量。(2)当胁迫时间大于等于30 d、受体植物为烟草时,转MT 植物与其野生型对照的叶绿素含量在胁迫程度上无明显差异。
本研究还发现,目的基因来源不同,转基因株系中目的基因表达的量不同,重金属运输的机制不同,也就会导致叶绿素含量不同。镉作为非必需的金属元素,在植物体内的运输机制还不十分明确, MT 基因是否会通过镉离子调控或影响其它必需元素(铁、镁等)的吸收、运输,从而影响到叶绿体合成的机制有待进一步探究。
猜你喜欢
小小说月刊·下半月(2021年8期)2021-09-05
少儿科学周刊·少年版(2021年17期)2021-01-17
少儿科学周刊·儿童版(2021年17期)2021-01-17
阅读(科学探秘)(2020年8期)2020-11-06
初中生世界·九年级(2020年9期)2020-09-21
动漫界·幼教365(大班)(2018年3期)2018-05-14
湖南大学学报·自然科学版(2016年12期)2017-05-12
女性天地(2016年10期)2017-04-25
东方教育(2016年9期)2017-01-17
江苏农业科学(2016年4期)2016-06-14
