
大白菜叶片表皮毛特征及重金属镉砷的累积特性
2022-02-21 09:33高培培孙洪欣李明月李瑞鹏刘文菊
河北农业大学学报 2022年1期
高培培,郭 佳,孙洪欣,董 妍,李明月,李瑞鹏,刘文菊
(河北农业大学 资源与环境科学学院/省部共建华北作物改良与调控国家重点实验室/河北省农田生态环境重点实验室/河北省蔬菜产业协同创新中心,河北 保定 071000)
2014 年《全国土壤污染状况调查公报》显示,我国耕地土壤点位超标率19.4%,重金属为主要一类污染物,尤其是污水灌溉区超标点位占26.4%,主要重金属污染物为镉(Cd)和砷(As)。净土方能洁食,农田土壤重金属污染深刻影响着土壤健康和农产品安全,并通过食物链威胁人体健康[1]。重金属镉、砷均被国际癌症研究组织确认为人类一级致癌物,我国南方成人摄入含无机砷稻米的致癌风险中值为20.1 人/万[2],在土壤镉污染严重的稻米产区,高暴露人群仅需4.7 ~8.3 年可达中等Cd 毒害症状[3-4]。稻米是我国南方人群摄入镉砷的主要方式[5],而在北方,尤其是冬春季,主要叶菜类蔬菜大白菜则是人体镉砷摄入的主要方式之一[6],由此产生的农产品健康风险不容忽视。
由于多年污水灌溉、金属冶炼废气排放、含重金属化肥的过量施用,造成我国蔬菜重金属超标的现象时有发生。北京地区部分蔬菜镉超标[7];东北三省的污灌区和矿区均存在蔬菜Cd 含量超标的情况,样品超标率在20%以上[8],最高可达61.0%[9-10]。在华北地区设施蔬菜也出现了不同程度的Cd、Pb 超标问题,尤其是叶菜类蔬菜[11]。蔬菜对人类镉摄入量的贡献率超过70%[12]。大白菜(Brassica rapaspp. pekinensis)作为大众最喜爱的叶菜类蔬菜之一,营养丰富,味道鲜美,有“百菜之王”之美誉,是我国栽培面积最大的露地十字花科蔬菜作物,而京津冀地区大白菜种植面积占全国的1/10 以上[13-14]。 本课题组前期研究表明大白菜中镉砷由根系向叶片的转运系数远高于小麦、玉米、水稻等粮食作物(数据未发表),造成叶片中镉砷重金属的富集,生长在污染土壤中的大白菜并未出现中毒或者抑制生长的现象。已有研究表明,十字花科的印度芥菜叶片表皮毛中的Cd 含量比叶片组织高43 倍,印度芥菜作为重金属超富集植物可把重金属储存在叶片表皮的表皮毛中,以避免出现重金属伤害叶肉细胞的情况,从而达到解毒的作用[15]。那么,大白菜中镉砷的富集是否也与表皮毛有关,则值得进一步探讨。
基于此,本研究以叶片有表皮毛的3 个大白菜基因型(野生型A03,突变体lcc-1,268)、叶面光滑无表皮毛的1 个基因型(CC48)和重金属镉(Cd)、砷(As)复合污染土壤为主要研究对象,通过盆栽试验探究3 个大白菜基因型叶片表皮毛的外观特征,以及采用扫描电镜-能谱的方法表征镉、砷在表皮毛基部的分布状况,最终诠释镉砷在大白菜叶片的累积特性,为培育镉砷低积累的大白菜品种提供数据支持。
1 材料与方法
1.1 试验材料
供试大白菜种质材料:4 个种质材料分别为野生型A03,野生型A03 生物钟突变体lcc-1,自交系268 和CC48,其中野生型A03、突变体lcc-1和268 的叶片着生表皮毛,而CC48 叶片无表皮毛。种子均由河北农业大学园艺学院申书兴教授和赵建军教授团队提供。
供试土壤:土壤样品采自保定某污染区的镉砷复合污染土壤,土壤全Cd 含量为3.89 mg/kg,全As 含量为177 mg/kg(为重度Cd、As 复合污染土壤),土壤质地为中壤土,pH 为8.55,有机质含量17.82 g/kg,有效磷含量42.06 mg/kg,速效钾含量344 mg/kg。将所取土样采回后放于室内自然风干,去除植物根系及石块,全部过5 mm 筛后待装盆备用。
1.2 大白菜幼苗培养
采用琼脂+营养液的联合培养方式进行育苗,首先将琼脂和营养液按1% 的比例混合加热溶解后,装入1.5 mL 离心管中,冷却得到固体营养琼脂,离心管下端剪口,置于装有霍格兰营养液的大白菜水培箱中,箱盖上有小孔可支撑离心管。然后将大白菜种子播种于琼脂表面,下按2 ~3 mm 即可。霍格兰营养液组成:KNO310 mmol/L;CaCl20.7 mmol/L;MgSO40.5 mmol/L;KH2PO41 mmol/L; H3BO350 µmol/L;MnCl212 µmol/L;CuSO41 µmol/L;NaMoO40.3 µmol/L;ZnSO43 µmol/L;FeNa-EDTA 50 µmol/L,pH 调 至5.8。播 种 后 将水培箱放置于植物光照培养箱中,保持16 h/8 h 光照/黑暗循环,温度维持在22 ~25 ℃,光照强度 280 mol/m2s。4 种基因型大白菜各设置4 个重复。培养10 d 后移入培养钵进行土壤盆栽试验。
1.3 土壤培养试验
将所取污染土过5 mm 筛后混入氮磷钾养分,施肥量N 200 mg/kg、P2O5150 mg/kg、K2O 200 mg/kg,分别以尿素、磷酸二氢钾和氯化钾的形式施入,充分混合均匀后将520 g 土壤装于黑色塑料培养钵中,每盆放入1 株大白菜幼苗,共16 盆,4 个基因型的培养钵随机排放,每周移动1 次位置,放于人工气候室中,培养的温度和光照条件同育苗的植物培养箱。培养2 个月后,收获植株地上部的新叶和老叶进行表皮毛的观测和重金属镉砷含量的测定。
1.4 样品采集及测定
植物样品生长2 个月后,每个基因型的单株大白菜分别长有8 ~10 片叶,定义大白菜生长点处的新叶为第1 片叶。因此,分别采集3 个有表皮毛的基因型大白菜的第6 片叶,首先观测表皮毛的生长情况,其次将叶片剪成1 cm×1 cm 的形状后迅速投入装有2.5%戊二醛固定液的5 mL 离心管中,在4 ℃ 条件下寄往山东大学大型仪器分析中心采用带能谱的扫描电镜(SEM-EDX)(Oxford, X-Max 20, UK)观测表皮毛的形貌特征、扫描表皮毛中镉砷的分布状况及其基部重金属镉砷的累积特性;然后,采集4 个基因型大白菜的新叶和老叶(新叶为第1至4 或5 片叶,老叶为剩余的其他叶片),每个基因型取4 个重复,超纯水冲洗后放入鼓风干燥箱(GZX-9240MBE, 上海博讯科技有限公司)中85 ℃ 杀青15 min,之后65 ℃烘至恒重备用。
植物样品的消煮和测定:准确称取粉碎后的大白菜新叶和老叶样品(精确至0.500 ±0.005 g)分别置于50 mL 离心管中,加入5 mL 68% 的HNO3,拧紧瓶盖后回拧半圈,浸泡过夜;次日,将离心管置于石墨消解仪上(SH230,中国海能仪器),120 ℃ 加热消解,1 h 后,将离心管取出,冷却至室温后,加入2 mL 30% H2O2,继续消煮,直至消解完全;1 h 后,打开盖子,赶酸至1 mL,超纯水定容至50 mL,即为消解原液,采用电感耦合等离子体质谱仪(ICP-MS, NexION 350X, 美国珀金埃尔默)测定叶片中Cd 和As 含量,以国家一级标准物质(GBW10047,芹菜)进行准确度和精密度控制,并同步分析空白样品以去除试剂干扰。所有重金属的回收率为75%~90%,均符合质量控制要求[16-17]。
1.5 数据处理与分析
富集系数(BF: Bioaccumulation factor)= 植物地上部重金属含量/土壤中重金属含量;
采用Microsoft Excel 2019 进行数据分析,SPSS 24.0 统计软件进行统计学分析,采用单因素方差分析法(One-way ANOVA)进行显著性检验,采用最小显著差异法(LSD)进行不同基因型大白菜地上部分重金属含量、富集系数以及表皮毛密度的差异显著性比较。
2 结果与分析
2.1 3 个大白菜基因型表皮毛的形貌特征及其镉砷的累积与分布
2.1.1 表皮毛的形貌特征 从图1 可以看出大白菜基因型A03、突变体lcc-1和268 的叶片着生密集的表皮毛。具体形貌特征描述如下:
图1 的a1、a2 为A03 基因型大白菜第6 片叶,叶缘无明显缺刻,叶面平整,叶脉细密,多毛。表皮毛色白呈直立形且在上表皮为匍匐状趋向植物表皮,叶脉及叶柄中则方向垂直于叶表皮,不均匀分布于整个叶片,在上表皮中,叶缘表皮毛明显密于叶片表面,下表皮的主脉表皮毛较侧脉、小脉和叶表面更为密集。通过测量叶上表皮叶缘每1 cm²表皮毛数量,分别为31、36、35 根。
图1 的b1、b2 为突变体lcc-1基因型大白菜第6 片叶,其叶片及表皮毛的形貌特征与野生型A03相似,但是,与A03 不同之处在于叶缘与上表皮均着生密集的表皮毛,下表皮的表皮毛集中分布在叶柄和叶脉。通过测量叶上表皮叶边缘每1 cm²表皮毛数量,分别为24、22、23 根,显著低于A03 的表皮毛数量(P<0.05)。
图1 的c1、c2 为268 基因型大白菜第6 片叶,叶缘及叶面皱褶多皱,叶脉粗密,多毛。表皮毛色白且直立,叶缘表皮毛朝向两侧,上表皮及下表皮的表皮毛方向均垂直于叶表皮,不均匀分布于整个叶片。通过测量叶上表皮叶边缘每1 cm²表皮毛数量,分别为13、14、14 根,显著低于A03 的表皮毛数量(P<0.05)。
图1 的d1、d2 为CC48 基因型大白菜第6 片叶,株行直立,叶片深绿呈平头长形,叶缘皱褶,叶面多皱,叶脉粗密,表面光滑无表皮毛。

图1 3 个基因型大白菜表皮毛外观图片Fig. 1 The images of trichome of three varieties of Chinese cabbage
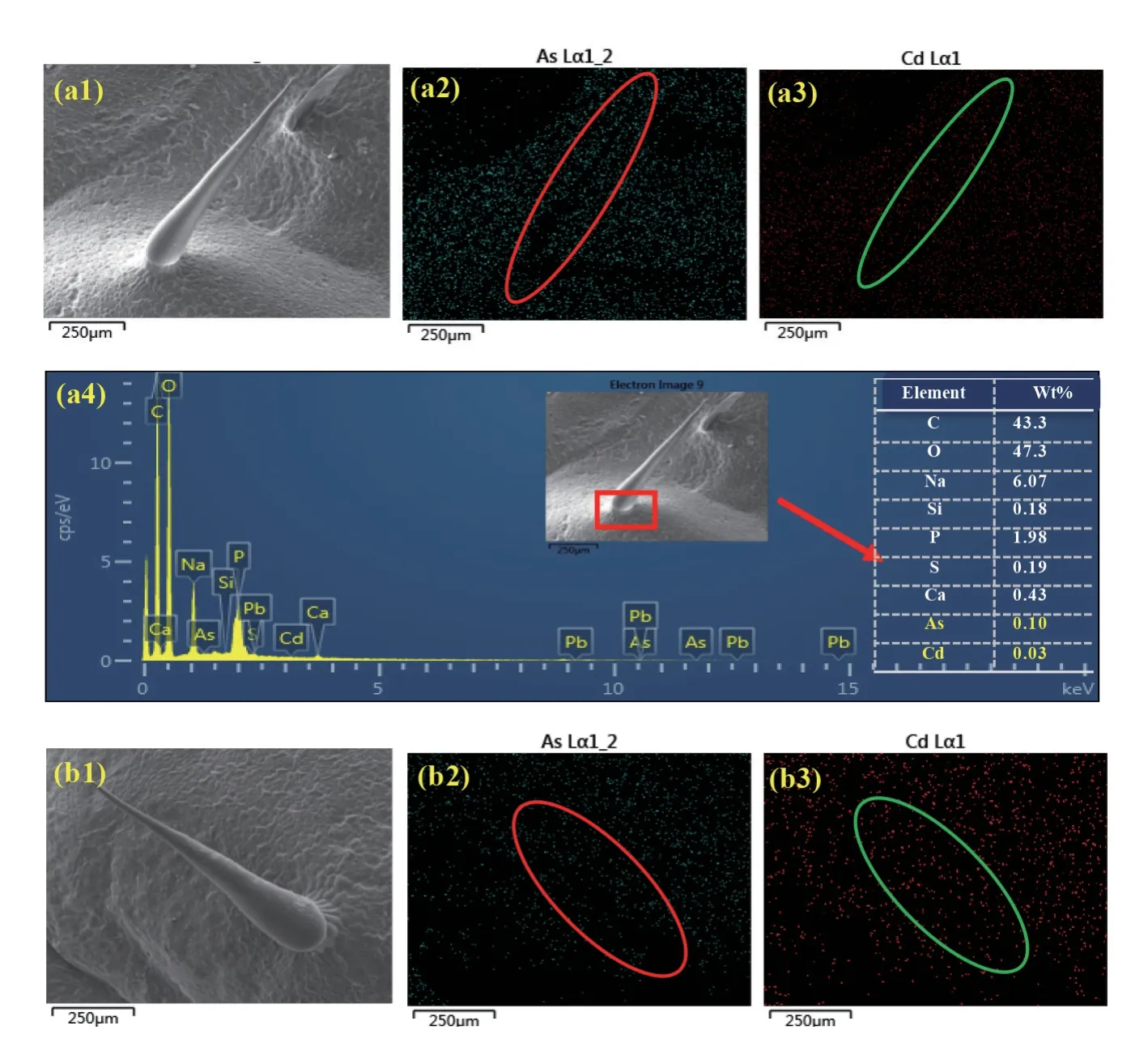
2.1.2 扫描电镜观察表皮毛的表型特征 分别选取3 种基因型大白菜长势良好、形态完整的第6片叶,电镜扫描观察不同基因型大白菜表皮毛的 形态(图2)。
A03 基因型大白菜的表皮毛较为光滑,基部与叶片表面连接不紧密,且连接处似有细胞凸起托住表皮毛。全长864 μm(图2a);突变体lcc-1大白菜叶片表皮毛粗糙,细胞壁坑洼明显,基部与叶片表面连接不紧实,连接处有细胞凸起托住表皮毛,全长870 μm(图2b);268 基因型大白菜的表皮毛外表粗糙,基部与叶片表皮连接紧密且有裂痕,测量后得出表皮毛全长722 μm(图2c)。3 种基因型表皮毛全貌照片大致相同,为圆锥状,无分枝,基部宽,末端渐尖。
2.1.3 扫描电镜能谱表征大白菜表皮毛中重金属镉砷的分布 结合扫描电镜观测的整根表皮毛的全貌特征图,用扫描电镜配备的电子探针能谱扫描表皮毛中镉砷的分布以及表皮毛基部镉和砷的微区定位。
由A03 大白菜叶片表皮毛的Mapping 图可以看出,As 和Cd 在表皮毛中广泛分布(图3 a2、a3),其中图3 的a2 中As 的分布强度较a3 中Cd的分布强。由表皮毛基部能谱扫描结果可以看出,表皮毛基部主要组成元素为C、O、Na,质量百分比分别占43.3%、47.3%、6.07%。此外,在此部位也有一定比例的As 和Cd 存在,其质量百分比分布为0.1%和0.03%,其中As 的峰值主要出现在P、S、Si 和Pb 的峰值附近,说明砷与物理化学性质类似的元素P、Si 可能结合在一起共同存在于表皮毛的基部。Cd 的质量百分比明显小于As,且与元素Ca的峰值较为接近,说明Cd 与Ca 在A03 表皮毛的基部结合在一起(图3 a4)。
图3 的b1 ~b3 为突变体大白菜叶片表皮毛Mapping 图,均表明As、Cd 广泛存在于突变体大白菜的表皮毛中,其中图3 的b3 中Cd 的分布明显强于b2 中As 的分布。对其基部的能谱扫描显示,其主要成分同样为C、O、Na,质量百分比分别为43.16%、45.58%、7.07%,表皮毛基部As 占0.09%,Cd 占0.24%,As 的含量与野生型差别不大,但是Cd 含量远高于野生型(图3 b4)。元素As 和Cd在表皮毛基部的赋存形态与野生型类似。
通过扫描268 基因型大白菜叶片表皮毛的Mapping 图发现(图3 c1 ~c3),砷主要附着在表皮毛细胞壁上,而表皮毛基部Cd 的信号较弱(图3 c1、c2)。表皮毛基部的能谱图显示,基部主要为C、O、Na 元素,质量百分比分布为55.14%、36.74%和4.59%,As 的相对质量百分比为0.03%,并未检测到Cd 的信号(图3 c4),故无法得出准确占比。由图c4 可见,大白菜268 基因型中As 和Cd 在表皮毛基部的赋存形态与野生型和突变体类似,但强度较弱一些。
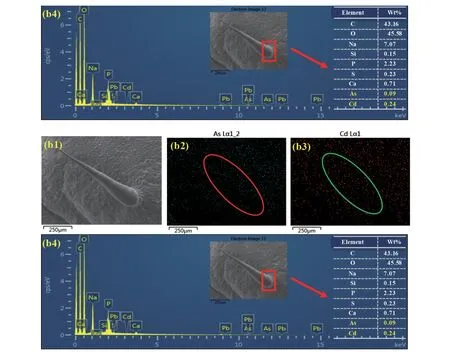
图3 3 种基因型大白菜表皮毛扫描电镜能谱图Fig. 3 The EDX spectrum analysis of trichome of three varieties of Chinese cabbage
2.2 不同基因型大白菜对复合污染土壤中镉、砷的生物富集能力分析
2.2.1 镉 通过比较4 种基因型大白菜新、老叶片中Cd 含量发现(图4),在镉砷复合污染土壤中生长2 个月后,A03 基因型大白菜新叶中Cd 含量最高可达3.92 mg/kg,比突变体lcc-1、268 和CC48 大白菜新叶中Cd 含量显著高0.91、1.13 和1.16 倍。而在老叶中,镉在不同基因型中的含量趋势与新叶相似,A03基因型大白菜老叶中Cd含量最高可达9.42 mg/kg,分别比突变体、268 和CC48 高出1.51、1.50、3.66 倍。对于4 种基因型大白菜(A03、突变体、268 和CC48)而言,老叶中Cd 含量分别为新叶的2.40、1.83、2.05、1.12 倍。从新叶到老叶,着生表皮毛的A03、突变体lcc-1和268 基因型的大白菜叶片Cd 含量的增幅明显高于无表皮毛的CC48,而无表皮毛的CC48 基因型大白菜叶片中Cd 含量几乎没有变化。这说明随着叶龄的增大,表皮毛的存在可以促进镉在叶片中的累积,且主要累积在表皮毛中。
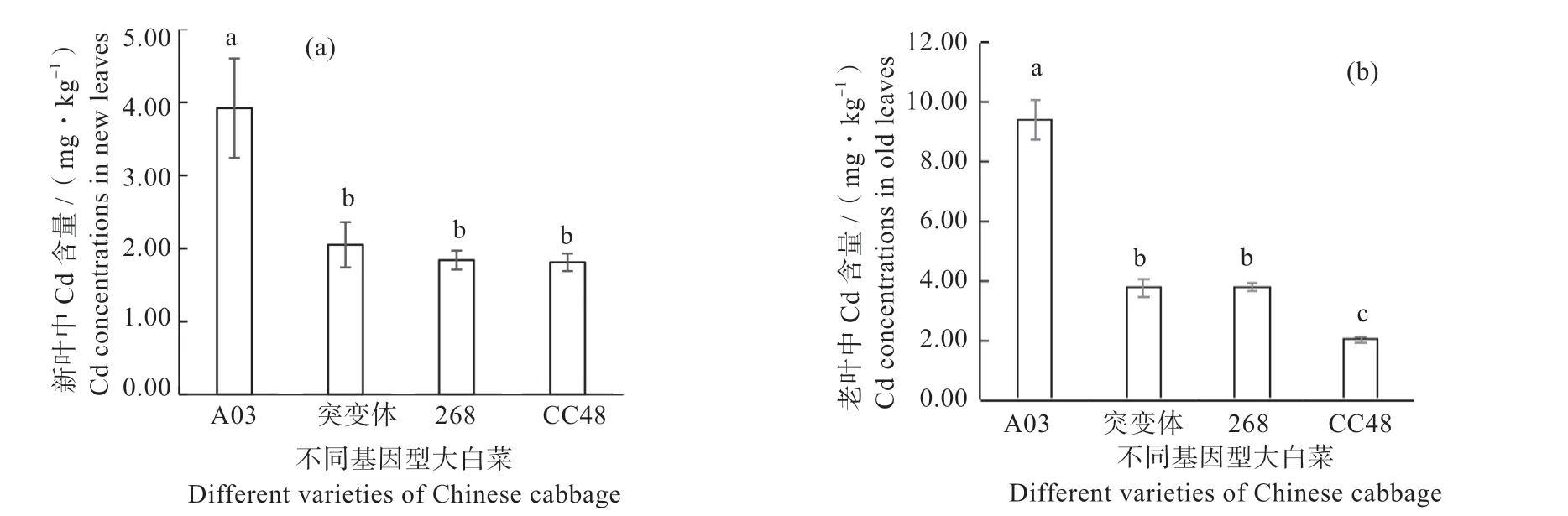
图4 不同基因型大白菜新叶(a)和老叶(b)中Cd 含量Fig. 4 The Cd concentrations in new (a) and old leaves (b) of different varieties of Chinese cabbage
富集系数是大白菜从土壤中吸收重金属的能力的表现,富集系数越大说明其富集重金属的能力越强。由图5 可以看出,在相同镉污染土壤中,不同基因型大白菜的Cd 富集系数存在明显差异,A03新叶对镉的富集系数最高为1.01,分别比突变体、268 和CC48 显著高0.91、1.15、1.20 倍;A03 老叶对镉的富集系数也最高达2.42,同样分别比突变体、268 和CC48 显著高1.31、1.31、3.65 倍;从新叶到老叶,不同基因型大白菜对镉的富集系数都有所提高,A03、突变体、268 和CC48 分别增幅140%、83%、106%、13.0%,长有表皮毛的3 个基因型老叶对镉的富集能力显著高于新叶,而没有表皮毛的CC48 基因型老叶和新叶对镉的富集能力差异不显著。这也说明随着叶龄的增加,有表皮毛的叶片对重金属镉的富集能力明显强于没有表皮毛的大白菜基因型,过多的镉富集在表皮毛中。

图5 4 种基因型大白菜新叶(a)和老叶(b)对Cd 的富集系数Fig. 5 Bioaccumulation factors of Cd in new (a) and old (b) leaves of different varieties of Chinese cabbage
2.2.2 砷 通过比较4 种基因型大白菜新、老叶片中As 含量发现(图6),在镉砷复合污染土壤中生长2个月后,A03 基因型大白菜新叶中As 含量最高可达3.49 mg/kg,比突变体、268 和CC48 大白菜新叶中As 含量显著高2.14、4.06、7.72 倍。对于老叶而言,As 在不同基因型大白菜中的含量趋势与新叶相似,A03 基因型大白菜老叶中As 含量最高可达10.97 mg/kg,分别比突变体、268 和CC48 高出87.2%、149%、778%。这说明表皮毛数量较多的A03 无论新叶还是老叶对As 的富集能力最强。同样,突变体中As 含量也显著高于无表皮毛的CC48。这说明表皮毛对大白菜叶片富集砷有一定的贡献。此外,对于4种基因型而言,老叶中As 含量分别为新叶的3.14、5.28、6.38、3.13 倍。由此可见,无论是着生表皮毛的A03、突变体和268 基因型,还是无表皮毛的基因型CC48,大白菜老叶中As 含量明显高于新叶。这说明随着叶龄的增大,As 在叶片中的累积逐渐增多,这种累积不仅表现在表皮毛中,也表现在叶肉细胞中,这一点与表皮毛对镉的富集不同。
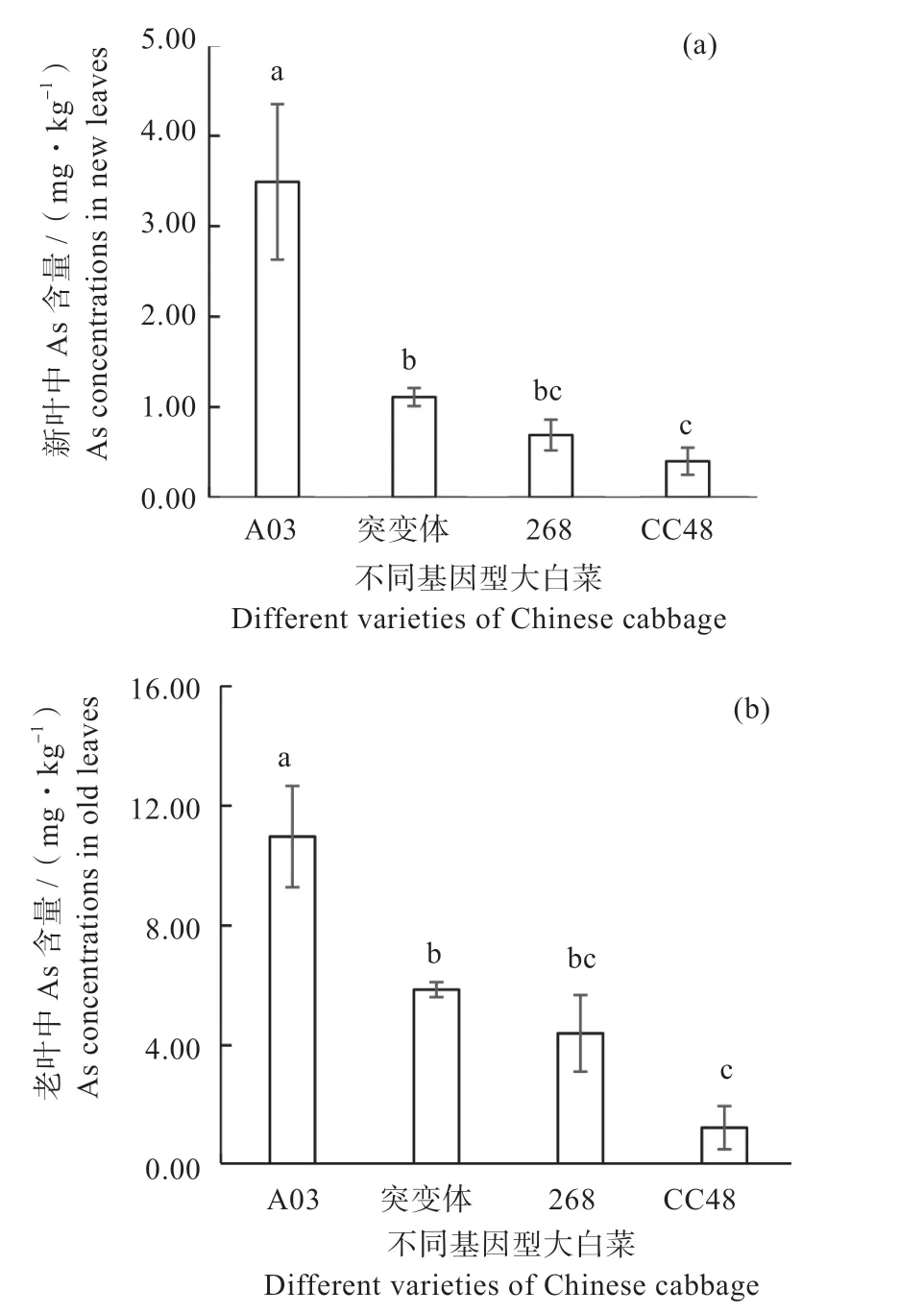
图6 不同基因型大白菜新叶(a)和老叶(b)中As 含量Fig. 6 The As concentrations in new (a) and old leaves (b) of different varieties of Chinese cabbage
由图7 可以看出,在相同砷污染土壤中,不同基因型大白菜的As 富集系数存在明显差异,A03 新叶对砷的富集系数最高为0.02,分别比突变体、268和CC48 显著高2.33、4.00、9.00 倍;A03 老叶对砷的富集系数也是最高为0.06,同样比突变体、268和CC48 显著高0.88、1.48、7.86 倍。与没有表皮毛的CC48 相比,表皮毛数量较多的A03 和突变体大白菜叶片对As 的富集能力显著增强,说明表皮毛对大白菜叶片中As 的富集有贡献。然而,从新叶到老叶,长有表皮毛的3 种基因型大白菜对砷的富集系数,以及没有表皮毛的CC48 基因型大白菜对砷的富集能力均显著提高。这说明随着叶龄的增加,大白菜叶片对重金属砷的富集能力明显增强,这种富集能力的增强与叶龄或者在污染土壤中的生长时间长短有关,其中表皮毛在大白菜叶片富集As 的过程中也起到了一定的辅助作用。

图7 不同基因型大白菜新叶(a)和老叶(b)对As 的富集系数Fig.7 Bioaccumulation factors of As in new (a) and old (b) leaves of different varieties of Chinese cabbage
3 讨论与结论
毛状体是叶原基发育过程中首先从表皮细胞分化而来的细胞[18],起源于植物外表皮细胞组织,是许多植物表面的附属物,如水稻、烟草、番茄、马铃薯、大白菜和棉花(棉纤维)等。表皮毛参与各种生物功能,如缓冲作用,增加植物表皮层的厚度,在表皮层外建立抵抗外来入侵的屏障[19]。表皮毛由单个或多个细胞构成,有分枝或没有分枝;有腺体或没有腺体。植物表皮毛的大小和密度并非一成不变,会根据生长发育阶段和所处位置的不同而改变[20-21]。大白菜表皮毛属于单细胞非腺体,没有分泌的功能,但是否对转运至此的重金属具有区隔化作用?本研究发现,叶片上着生较多表皮毛的大白菜基因型A03 和突变体,无论老叶还是新叶对污染土壤中镉砷的富集能力均显著高于无表皮毛的CC48,虽然前者叶片中富集了较多的镉砷,但是生长并没有受到抑制,这说明表皮毛对吸收转运至大白菜叶片的镉砷具有区隔作用,从而起到解毒的作用。由于表皮毛体细胞内富含大量的线粒体,因此可以说表皮毛是一个代谢活性很强的细胞,本身有强大的生命力和适应能力[22]。此外,研究发现,在组织水平上,重金属主要存在于表皮细胞、亚表皮细胞和表皮毛中;细胞水平上,重金属主要存在于质外体和液泡中,证明重金属在植物的叶片中的确存在区室化分布[18]。这些证据都证实了本研究的结果。
土壤重金属污染是隐蔽、长期且不可逆的,其治理或者修复是一个世界性难题。对于镉砷复合污染严重的农田土壤,不可否认,采用种植重金属超富集植物的植物提取方法来修复污染的土壤是一项既经济、美化环境又有利于土壤生态恢复的技术。然而,我国人多地少是不争的事实,若大面积种植利用价值少和经济效益低的超富集植物来修复污染土壤,不利于我国农业经济的发展和进步。此外,河北省乃至整个中国农田重金属污染程度主要为轻污染,占总超标点位的85.1%[2,23]。因此,对于轻中度镉砷污染且农作物可以正常生长的农田,更应该采用植物固定或者利用植物本身的特性,探索其解毒的作用机制,从而实现轻度污染土壤的安全利用。本研究采用不同的大白菜基因型来探究其在镉砷污染土壤中的生长状况、叶片表皮毛的形貌特征及其对重金属镉砷的富集能力,一方面可以将具有表皮毛的大白菜种植在镉砷污染较重的菜田土壤中,因为表皮毛的区隔化的解毒作用,使大白菜从土壤中吸取大量的镉砷污染物,使土壤中镉砷含量水平逐年降低,达到修复土壤的目的,另一方面,可以将没有表皮毛且镉砷含量很低的基因型种植在污染程度较轻的土壤中,使大白菜可食部位镉砷含量符合我国农产品食用安全标准,实现镉砷轻度污染土壤的安全利用。
综上所述,相较于无表皮毛的大白菜,有表皮毛的大白菜老叶对Cd、As 具有更高的累积特征;野生型A03 的表皮毛数量显著多于突变体,其叶片中Cd、As 含量也显著大于突变体,表明表皮毛与重金属镉、砷的解毒机制有关。表皮毛对Cd、As的区隔化作用是大白菜解Cd、As 毒的作用机制。
猜你喜欢
作物学报(2022年2期)2022-11-06
作物学报(2022年8期)2022-05-29
作物学报(2022年6期)2022-04-08
金山(2020年9期)2020-11-10
散文选刊·下半月(2020年6期)2020-07-14
故事作文·低年级(2018年2期)2018-02-22
东坡赤壁诗词(2017年3期)2017-07-05
语文世界(初中版)(2017年1期)2017-03-06
大江南北(2014年3期)2014-11-23
