
miR-125靶向SMAD4抑制脂多糖诱导的人结肠上皮细胞向肿瘤细胞转化
2022-02-19 10:20张聪敏郭金培马二民
基础医学与临床 2022年2期
曾 笛,张聪敏,郭金培,张 楠,马二民
(1.郑州市第七人民医院 普外科,河南 郑州 450000; 2.河南中医药大学第一附属医院 普外科, 河南 郑州 450000)
肠道菌群通过分泌脂多糖(lipopolysaccharide,LPS)介导的异常炎性反应可能对结直肠癌的发生发展具有重要作用[1-2]。正常生理状态下,肠道炎性可促进组织损伤修复[3]。但肠道菌群功能失调时,菌群释放过量LPS,导致肠道炎性反应亢进,常伴随结肠上皮细胞恶性细胞转化[4]。microRNA(miRNA)可与靶基因的3′-UTR区域特异性结合而影响靶基因的降解或翻译蛋白产物表达[5-6]。TGF-β信号通路被发现与多种癌症的发生和发展相关,且SMAD4在其介导的肿瘤细胞功能转换过程有重要意义。其中miR-125被认为是SMAD4结合的关键miRNA之一,但在LPS诱导的结肠上皮炎性微环境中的作用尚未报道。因此,本研究拟利用LPS对正常结肠上皮细胞进行低浓度长期炎性刺激,探索炎性微环境在正常结肠上皮细胞向肿瘤细胞转化中的作用。
1 材料与方法
1.1 材料与试剂
LPS(上海爱必信生物科技有限公司);人结肠上皮细胞系HCoEpiCs(中国科学院上海细胞库);DMEM培养基、胎牛血清(Thermo Fisher公司);CCK-8试剂盒(南京福麦斯生物科技有限公司);miR-125 mimic和inhibitor套装(广州锐博生物科技有限公司);SMAD4-3′-UTR野生型和突变型质粒(上海吉凯基因公司);Lipofectamine3000(Invitrogen公司)氯仿、异丙醇和无水乙醇(上海沪试试剂有限公司);Trizol、mRNA反转和定量试剂盒(TaKaRa公司);miRNA提取、定量试剂盒(北京天根生物科技有限公司);萤火虫/海肾底物发光试剂盒(Promega公司);4%多聚甲醛(南京迈博生物有限公司);吉姆萨染色液(广州泛思生物科技有限公司);琼脂(北京索莱宝生物科技有限公司);引物(上海生工生物工程有限公司)见表1。

表1 引物序列Table 1 The sequence of primer pairs
1.2 方法
1.2.1 细胞培养及分组处理:在37 ℃、5% CO2的细胞培养箱中,用含10%胎牛血清的DMEM培养基培养HCoEpiCs结肠上皮细胞。当观察到细胞在培养瓶底面汇合达到80%时,用0.25%胰蛋白酶(含EDTA)消化细胞2 min,加入含10% FBS的完全培养基终止消化,离心后弃上清,重悬并加入到含新鲜培养基的培养皿中。培养24 h后,以CCK-8法实验确定的最佳LPS刺激浓度为后续诱导上皮细胞恶性转化的刺激浓度。用培养基将LPS稀释到最佳刺激终浓度培养HCoEpiCs细胞并传代,连续重复此细胞刺激和传代操作至40代。
以8×104个/mL的浓度铺细胞到6孔板内,每个孔加2 mL的完全培养基,放到培养箱内24 h。将miR-125 mimic、inhibitor、mimic-NC和inhibitor-NC(50 nmol/L)用125 μL无双抗的DMEM混匀,将5 μL Lipofectamine3000用125 μL无血清和双抗的DMEM混匀并静置5 min。上述两组液体充分混匀后,室温下放置15 min后加入6孔板。在培养箱中培养48 h后,进行后续实验。
1.2.2 CCK-8法检测细胞活性:收集对数增值期的细胞,将5×103个细胞接种于96孔板,每孔培养基体积为200 μL,培养24 h后观察细胞是否均匀贴壁。分为0(空白对照组)、5、10和15 μg/mL刺激组,分别加入不同浓度含LPS的完全培养基,继续培养24 h后每孔加入10 μL CCK8试剂,在细胞培养箱中孵育2 h后用酶标仪检测450 nm处吸光度值。
1.2.3 RT-qPCR检测HCoEpiCs结肠上皮细胞mRNA:向6孔板中加入1 mL的RNAiso Plus反应5 min并转移到1.5 mL无酶离心管中。加200 μL氯仿。4 ℃,12 000×g离心15 min,吸取上清到新无酶离心管内。加500 μL异丙醇,室温放置10 min。4 ℃,12 000×g离心10 min,弃上清。加75%乙醇1 mL,4 ℃、7 500×g离心5 min。离心管倒置10 min后,加无酶水使离心管内沉淀溶解,在核酸蛋白分析仪上测定RNA浓度。用PrimeScriptTMRT试剂盒进行反转录。用TB Green试剂盒进行荧光定量。反应体系为:mix 10 μL,正向引物0.4 μL,反向引物0.4 μL,RoxⅡ 0.4 μL及6.8 μL的无酶水。反应条件:95 ℃预变性5 min,95 ℃变性5 s,60 ℃退火延伸30 s,共进行45个循环。以GAPDH或U6为内参,所得数值采用2-ΔΔCt计算相对表达量。
1.2.4 平板克隆和软琼脂集落检测HCoEpiCs结肠上皮细胞向肿瘤细胞转化:取LPS刺激后1代、30代和40代对数增值期的细胞,经胰蛋白酶消化后将细胞重悬在10%完全培养基中。以每孔500个细胞的密度分别均匀接种在6孔板中。在细胞培养箱中培养2周。当孔中出现肉眼可见的克隆细胞团时,弃去上清液,用PBS清洗3次。每孔加1 mL的4%多聚甲醛固定细胞15 min。小心吸去固定液,加200 μL的吉姆萨染色液染20 min,用流水缓慢洗去染色液,用肉眼直接计数克隆,计算克隆形成率。
配制1.2%和0.7%的琼脂,经高压灭菌后,放入42 ℃水浴锅中,将1.2%琼脂与2×培养基混合,每孔1.5 mL铺入6孔板作为下层,37 ℃孵育30 min使之凝固;将单细胞悬液与1 mL的0.7%琼脂充分混匀,以每孔5 000个细胞的密度加入6孔板作为上层胶。凝固后于上层胶上加入200 μL完全培养基,每隔3 d更换一次培养基。10 d后在显微镜下观察克隆大小,并拍照。
1.3 统计学分析
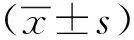
2 结果
2.1 不同浓度LPS对HCoEpiCs细胞活性的影响
与对照组相比,当LPS刺激浓度为10 μg/mL时细胞活力开始受到明显抑制,其抑制率随着药物浓度的递增而增加(图1,P<0.05),选取该浓度为后续细胞刺激浓度。

*P<0.05 compared with control group图1 不同浓度LPS对HCoEpiCs结肠上皮细胞活力的影响Fig 1 Viability of HCoEpiCs cells treated with different concentrations of LPS
2.2 LPS对HCoEpiCs细胞向肿瘤细胞转化的影响
细胞周期蛋白相关基因cyclinD1在刺激40代(P40)后表达量出现明显增加(P<0.05),而癌基因c-myc和抗凋亡基因Bcl-2均在刺激30代后(P30)表达量出现明显增加(图2A) (P<0.05)。平板克隆和软琼脂克隆实验发现LPS刺激后30代未出现明显的细胞克隆,而刺激后40代细胞形成明显的克隆(图2B,C)。

A.expression of cyclinD1,c-myc, and Bcl-2 in the continuously LPS-treated HCoEpiCs cells; B.colony formation of LPS-treated HCoEpiCs cells; C.soft agar colony formation assay of HCoEpiCs cells treated with LPS; *P<0.05 compared with P1; P1,P10,P20,P30,and P40 mean HCoEpiCs cells continually treated with LPS at the 1th,10th,20th,30th,and the 40th passage图2 LPS连续刺激对HCoEpiCs结肠上皮细胞向肿瘤细胞转化的影响Fig 2 Results of LPS-induced HCoEpiCs cells transformed to cancer cells
2.3 LPS诱导HCoEpiCs细胞中miR-125和SMAD4的表达情况
与P1代相比,P40代细胞中的miR-125表达出现明显的增加(P<0.05),而SMAD4表达出现明显的降低(P<0.05)(图3)。

A.relative expression of miR-125 in the 1th and 40th passage of LPS-induced HCoEpiCs cells; B.the level of SMAD4 in the 1th and 40th passage of LPS-treated HCoEpiCs cells; *P<0.05 compared with P1 group图3 HCoEpiCs结肠上皮细胞中miR-125和SMAD4表达情况Fig 3 Relative expression of miR-125 and SMAD4 in HCoEpiCs cells
2.4 干预miR-125对HCoEpiCs细胞SMAD4表达的影响
过表达miR-125后SMAD4和癌基因c-myc表达出现明显的降低(P<0.05),而抑制miR-125表达后SMAD4和c-myc均出现明显的增高(P<0.05)(图4)。

A.relative level of SMAD4 in the 40th passage of LPS-treated HCoEpiCs cells who transfected miR-125 mimic or inhibitor; B.expression of c-myc in the 40th passage of LPS-treated HCoEpiCs cells who transfected miR-125 mimic or inhibitor; *P<0.05 compared with mimic-NC group,#P<0.05 compared with inhibitor-NC group图4 干预miR-125对HCoEpiCs细胞相关mRNA表达的影响Fig 4 Effect on HCoEpiCs cells relative mRNA expression by interfering miR-125
2.5 miR-125靶向SMAD4并抑制该基因表达
过表达miR-125后,SMAD4的野生型3′-UTR相对荧光素酶活性出现了明显的下降(P<0.05);但SMAD4的3′-UTR区被突变后,相对荧光素酶活性未出现明显改变(图5)。

*P<0.05 compared with control group图5 HCoEpiCs细胞共转染miR-125 mimic和SMAD43′-UTR后荧光素酶活性Fig 5 Luciferase activities of HCoEpiCs cells who cotransfected with miR-125 mimic and relative SMAD4 3′-UTR
3 讨论
正常结肠细胞向肿瘤细胞转化过程常与结肠上皮损伤、化学促癌试剂和某些致癌生物毒素等外源性因素相关[7],也受患者自身基因突变、生活习惯和自身免疫等内源性因素影响。结肠上皮细胞癌变过程通常存在着许多促癌因素的诱导和多种关键基因的调控[8]。由于肠道菌群紊乱而导致革兰阴性菌过度释放LPS, 致使肠道处于长期慢性炎性刺激状态[9],进而导致结直肠癌的发生和发展。因此,观察LPS对正常结肠上皮细胞向肿瘤细胞转化的影响,探索该过程涉及的分子机制可能对结肠癌的防治具有重要意义。
本研究以人正常结肠上皮细胞HCoEpiCs为受试对象,通过CCK-8法探索了不同浓度LPS对该细胞活力的影响,发现LPS刺激浓度为10 μg/mL时细胞活力开始出现明显抑制,且该浓度与参考文献[10]在肺癌上皮细胞中的刺激浓度一致,因此选用该浓度为后续长期慢性炎性刺激的浓度。LPS对HCoEpiCs细胞连续刺激40代后,发现细胞周期相关基因cyclinD1、癌基因c-myc和抗凋亡基因Bcl-2均出现明显的增高,且平板克隆和软琼脂实验发现该细胞出现明显的细胞团,说明LPS可诱导HCoEpiCs细胞向肿瘤细胞转化。已有研究报道miR-125和TGF-β信号通路下游重要的效应分子SMAD4在肺癌、食管癌和乳腺癌等疾病中存在重要的靶向关系[11-12],但在LPS诱导的正常结肠上皮细胞中的表达情况和作用尚未可知。因此,本研究发现P40代细胞中miR-125出现明显的升高,而SMAD4出现显著的降低。利用mimic和inhibitor对miR-125干预后,SMAD4的表达均出现明显的改变,这提示miR-125和SMAD4在LPS诱导的HCoEpiCs向肿瘤细胞转化过程中可能存在重要的关系。通过检测癌基因c-myc的变化,结果提示干预miR-125可能通过靶向SMAD4对HCoEpiCs细胞癌变过程具有重要影响。随后的双荧光素酶基因报告实验也证明miR-125和SMAD4之间存在重要的靶向关系。因此本研究认为miR-125可靶向SMAD4进而抑制LPS诱导的结肠上皮细胞向肿瘤细胞转化。
本研究的局限性在于未探索沉默SMAD4后对LPS诱导的结肠上皮细胞向肿瘤细胞转化的影响,SMAD4作为TGF-β信号通路下游重要的效应分子,其在LPS诱导结肠上皮细胞癌变过程中的作用可能与结肠癌细胞的作用存在相同的机制,因此后续研究应重点探索SMAD4在LPS诱导结肠上皮细胞癌变过程中是否具有前期的抑制效应及后期的促进向肿瘤细胞转化的作用。
猜你喜欢
核科学与工程(2022年3期)2022-10-18
农产品加工(2022年17期)2022-10-17
福建医科大学学报(2022年3期)2022-09-10
中国农村水利水电(2022年8期)2022-08-28
上海交通大学学报(2022年8期)2022-08-25
化工进展(2022年7期)2022-08-01
应用数学和力学(2022年1期)2022-02-18
新农业(2021年9期)2021-06-20
家禽科学(2020年3期)2020-05-13
医食参考(2017年12期)2017-04-01
