
异养硝化-好氧反硝化菌氮代谢机理的研究进展
2022-02-17 14:07赵天涛陈沛沛
重庆理工大学学报(自然科学) 2022年1期
赵天涛,陈沛沛,张 晟,封 丽
(1.重庆理工大学 化学化工学院, 重庆 400054;2.重庆市生态环境科学研究院, 重庆 400014)
水中含氮化合物超标会促进藻类等浮游生物的过度繁殖,引起水体富营养化,导致水质恶化,对生态环境以及人体健康造成严重威胁[1]。相比于物化法,生物脱氮由于经济、高效、无二次污染等特点,被公认为是最有前途的脱氮方法[2]。传统生物脱氮过程中,氨氮首先被自养硝化菌在好氧条件下转化为硝态氮,然后由异养反硝化菌在厌氧或兼氧条件下将硝态氮还原为含氮气体[3]。但是由于不同脱氮菌群的生理机制差异,导致硝化和反硝化过程在空间和时间上无法统一,造成系统水力停留时间长、抗冲击负荷能力弱以及运行成本高等问题[4]。因此,开发新型生物脱氮工艺一直是废水处理领域的热点。
1983年,Robertson等[5]首次从废水中发现了异养硝化-好氧反硝化(HN-AD)菌Paracoccuspantotrophus,并提出了异养硝化-好氧反硝化的概念。此后,研究者开展了大量HN-AD菌的分离鉴定工作,截至目前,约有20个属近100多种HN-AD菌从环境中被分离出来[6],如Acinetobactersp.SYF26[7],PseudomonasmendocinaX49[8],BacilluscereusGS-5[9],AlcaligenesfaecalisWT14[10],AnoxybacilluscontaminansHA[11]等。这类细菌在好氧条件下,可以快速将氨氮、硝态氮和亚硝态氮转化为含氮气体,且氨氮代谢过程中几乎没有硝态氮和亚硝态氮的积累[12]。较传统脱氮微生物,HN-AD菌具有脱氮速率快、世代周期短等优势,在实际废水处理中可以简化工艺流程,缩短水力停留时间,降低运行能耗。此外,基于HN-AD菌研发的生物膜技术可实现污泥近零排放,碳排放显著降低[13]。
目前关于HN-AD菌的研究主要集中在菌株的分离鉴定、脱氮影响因素和关键酶分析等方面,相关的综述也多是上述研究的概括与分析,而HN-AD菌具体的脱氮机制尚缺乏系统、清晰的认识和总结[14-16]。据此,对HN-AD菌相关研究开展了广泛调研,并结合本课题组在HN-AD菌脱氮机理以及工艺方面的研究,从氮平衡角度对脱氮途径进行了系统性分析,同时总结了HN-AD菌氮代谢作用相关酶系,对未来HN-AD菌的应用提出了展望,以期为此类菌脱氮机制的研究及在实际废水处理中的应用提供参考。
1 HN-AD菌氮代谢途径研究
HN-AD菌可以利用氨氮、亚硝态氮或硝态氮为氮源,通过同化作用(assimilation)转化为细胞有机氮;也可以通过异化作用(dissimilation)将氮素转化为含氮气体(NO、N2O或N2)[17]。本文根据不同的氮源类型,对HN-AD菌的氮代谢途径进行了总结。
1.1 HN-AD菌的氨氮代谢途径

传统的硝化细菌为自养型细菌,在硝化反应过程中有机物浓度不宜过高,否则会使异养细菌快速增殖,硝化细菌无法成为优势菌属,硝化反应无法进行[23]。而HN-AD菌的好氧同化过程可同时去除废水中的有机物和氨氮。如陈均利等[24]在利用AlcaligenesfaecalisWT14处理实际养猪废水时氨氮、硝态氮、总氮、COD平均去除率均在90%以上。谭俊杰等[25]发现菌株GNR在猪场废水处理中,单位生物量COD和氨氮去除量最高可达129.73 mg/(L·g)。传统硝化细菌和反硝化细菌生理机制的巨大差异使污水处理系统中本有的有机物无法被充分利用。如果硝化在前,反硝化过程中需要外加碳源;如果反硝化工艺段前置,则需要大规模回流硝化废水,这些均不利于传统生物脱氮工艺的高效运行[23]。HN-AD菌能够解决这一难题,可以为一些高浓度有机废水的脱氮除碳处理提供新思路和新菌源。

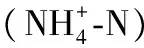
表1 不同HN-AD菌脱氮过程的氮平衡分析结果
1.2 HN-AD菌的硝态氮和亚硝态氮代谢途径



表2 不同HN-AD菌脱氮过程的氮平衡分析结果
1.3 不同生物脱氮工艺氮代谢途径比较

图1 不同生物脱氮工艺的氮代谢途径示意图
2 HN-AD菌氮代谢作用酶系
HN-AD菌种属多样,在不同菌株中,氮代谢相关酶基因的表达存在较大差异,这也是造成氮代谢途径有所不同的原因。参照传统脱氮微生物并根据HN-AD菌的氮代谢产物可以推测,菌株氮代谢过程中可能涉及的酶有氨同化酶、氨单加氧酶、羟胺氧化酶、硝酸盐还原酶、亚硝酸盐还原酶、一氧化氮还原酶和一氧化二氮还原酶[42]。
2.1 氨同化酶
氨同化的关键酶包括谷氨酸脱氢酶(glutamate dehy drogenase,GDH)、丙氨酸脱氢酶(alanine dehydrogenase,AlaDH)和谷氨酰胺合成酶(glutamine synthetase,GS)等[43]。谷氨酸脱氢酶分布广泛,近年来许多关于谷氨酸脱氢酶的研究已经证明其主要以六聚体和四聚体2种形式存在。谷氨酸脱氢酶是谷氨酸合成的关键酶,也是HN-AD菌发挥同化作用的关键酶之一,在菌株ArthrobacterureafaciensCZ31[44],Klebsiellasp.KSND[45]的氮代谢途径中,都发现有谷氨酸脱氢酶的活跃作用。丙氨酸脱氢酶能够催化丙氨酸与丙酮酸相互转化,与氨基酸代谢和糖代谢有关,在维持生物体内氮平衡(固氮作用)中起重要作用[46]。谷氨酰胺合成酶是生物体中最古老、存在最广泛的酶,可催化L-谷氨酸与L-谷氨酰胺的相互转化,参与许多生物体的氮代谢和碳代谢[47]。
2.2 氨单加氧酶(AMO)
2.3 羟胺氧化酶(HAO)
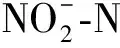
2.4 硝酸盐还原酶
2.5 亚硝酸盐还原酶
2.6 NO还原酶及N2O还原酶
NO还原酶(NOR)可催化NO还原为N2O,在HN-AD菌中已有较多报导,如ExiguobacteriummexicanumSND-01[34],BacillusthuringiensisWXN-23[58]等。NOR活性较强,使得NO一般不容易在系统中积累,避免了NO对微生物的毒性影响[59]。N2O还原酶(NOS)可以将N2O还原为N2。大多数HN-AD菌中分离得到的NOS都是可溶性的周质酶。如菌株Pseudomonas的NOS是由nosZ编码的亚基组成的二聚体[60]。值得注意的是,N2O还原酶对氧气敏感,控制适宜的溶解氧对N2O的还原至关重要[61]。
3 HN-AD菌氮代谢机理组学研究
HN-AD菌表现出的脱氮潜力和独特代谢机制吸引了越来越多学者对这类微生物的关注。目前,组学技术正在成为研究生物体内分子的综合方法[62],包括基因组学、转录组学、蛋白质组学和代谢组学。应用组学技术可以从多个层次阐明HN-AD菌的氮代谢机理。例如,在以往的研究中,Peng Jin等[45]通过基因组学发现与HN-AD菌氮同化和异化途径有关的基因,并利用实时定量PCR技术验证分析了不同氮源下基因的变化水平。Ruan等[63]通过转录组分析发现了高、低不同硝酸盐浓度下napA、nirS、norB、nosZ等反硝化基因表达的差异。碳源在好氧反硝化中起着重要作用,利用代谢组学可以分析不同碳源条件下好氧反硝化过程中关键代谢产物的变化[64]。据报道,铁的添加可以增强HN-AD菌的脱氮性能[65]。然而目前铁引发的机理尚不清楚,故Yang等[66]提出可以利用蛋白质组学来阐明铁是否通过帮助合成某些反硝化酶的活性中心来促进好氧反硝化。

图2 菌株TAC-1的氮代谢机制示意图

4 结论
上述工艺研究在一定程度上丰富了HN-AD菌的研究范围,但由于HN-AD菌的种类繁多,脱氮过程的酶系及其编码基因存在差异,导致HN-AD菌脱氮途径和机制尚不清晰,未来还需要更深入的研究和拓展,可以从以下4个方面进行:① 选择模式菌株,全面深入地解析不同HN-AD菌属的特征酶以及功能基因,通过酶的活性测定、分离纯化、体外表达以及功能基因敲除等方法,推测HN-AD菌同步脱氮的途径和机理;② 考察环境因子对HN-AD菌的影响,明晰菌体胞内生化反应的调节机制。通过设计正交实验或响应面分析法寻找最优水平组合和工艺参数;③ 充分利用新一代高通量测序技术,如蛋白质组学、代谢组学等多组学测序对HN-AD菌进行分析,研究HN-AD菌基因表达水平和各种代谢路径的底物和产物的小分子代谢物,从分子水平上推测其脱氮途径及机制;④ 研究HN-AD菌在各类真实废水中的作用,分析此类菌在氮循环过程中的地位以及如何优化工艺和条件参数使其成为优势脱氮菌,加强HN-AD菌的实际应用研究。
猜你喜欢
水土保持学报(2022年3期)2022-05-26
建材发展导向(2022年6期)2022-04-18
中国瓜菜(2022年1期)2022-02-18
中国生态农业学报(中英文)(2022年1期)2022-01-15
家庭科学·新健康(2021年5期)2021-06-21
科学与财富(2020年1期)2020-03-02
环境与发展(2019年12期)2019-02-12
家庭医药·快乐养生(2017年11期)2017-11-21
江苏农业科学(2016年7期)2016-10-20
农家顾问(2016年5期)2016-05-14
