
转多基因107杨对目标害虫的抗性检测*
2022-01-18 06:56:48韩昆瑾郭娟娟吕梓晴李玉言王世杰杨敏生王进茂
林业科学 2021年11期
韩昆瑾 郭娟娟 吕梓晴 李玉言 王世杰 杨敏生 王进茂
(河北农业大学林学院 河北省林木种质资源与森林保护重点实验室 保定 071001)
杨树(Populus)是世界上分布范围最广、用途最多、适应性最强的树种之一。在我国,杨树主要应用于木材生产、生态防护和通道绿化等方面,由于其具有高生长率和易繁殖等特点而受到广泛重视。然而,杨树在栽培种植过程中易受到虫害威胁,食叶害虫中鳞翅目(Lepidoptera)昆虫美国白蛾(Hyphantriacunea)及鞘翅目(Coleoptera)昆虫柳蓝叶甲(Plagioderaversicolora)等危害较为严重。虫害除影响杨树生长和木材质量外,还会给林业经济效益和生态效益带来巨大损失。据调查数据显示,我国每年因病虫害损失约 880亿元,其中直接经济损失145亿元,生态服务价值损失 735亿元(苏宏钧等,2004),同时虫害造成的叶损失会使森林由碳汇变为碳源 (Kurzetal.,2008)。利用植物基因工程解决杨树虫害问题是一种较为有效的途径。杨树作为林木基因工程中的一种模式植物,其在抗虫转基因研究中也取得了许多突破性进展(Zuoetal.,2018;Zhangetal.,2016),目前,苏云金芽孢杆菌(Bacillusthuringiensis)晶体蛋白基因(Bt)、蛋白酶抑制剂类基因(PI)和昆虫毒素基因等已应用于杨树抗虫转基因研究(睢韡等,2019)。但这些外源基因能否在受体植物中稳定表达,还需要进行长期观测。Hawkins等(2003)检测了杨树的44个不同转基因株系,发现1个沉默株系,推测在木本植物中基因沉默相对较少,且在转录开始便已沉默。胡建军等(2007)观察并检测了7年生的转Bt基因欧洲黑杨(Populusnigra),调查结果表明Bt基因能够稳定表达,且未发现害虫对转基因植株产生耐受性。转基因杨树中外源基因稳定性与环境也存在一定的相关性,树木的生长周期较长,基因沉默的可能性增加(魏冰等,2009)。
本研究以河北农业大学林木遗传育种实验室培育的转多基因欧美杨107杨(Populus×euramericana‘Neva’)为植物材料,对6个转基因株系2年生田间苗通过PCR技术鉴定外源基因是否稳定存在,研究Bt基因表达情况以及生长季的毒蛋白含量,以美国白蛾和柳蓝叶甲为测试昆虫检测转多基因107杨的抗虫性。通过分析转多基因107杨的外源基因表达情况及表达效率等问题,为转多基因杨树的品种培育和推广提供依据。
1 材料和方法
1.1 试验材料
1.1.1 植物材料 试验材料为转Cry1Ac-Cry3A-BADH基因107杨(简称V1)株系ECAA1、ECAA2、ECAA3及转Cry1Ac-Cry3A-NTHK1基因107杨(简称V2)株系ECAB1、ECAB2、ECAB3。其中,V1载体(图1a)携带抗虫的Bt基因Cry1Ac和Cry3A,同时携带有甜菜碱醛脱氢酶基因BADH,基因Cry1Ac和Cry3A的启动子为CAMV35S启动子,Cry1Ac基因接近载体右臂,Cry3A接近载体左臂;V2载体(图1b)同样携带抗虫的Bt基因Cry1Ac和Cry3A,同时携带有烟草乙烯受体基因NTHK1,基因Cry1Ac和Cry3A的启动子及位置与V1载体相同。以未转基因107杨为对照。转基因植株为栽植在河北农业大学西校区林学院专用苗圃中的2年生幼树,正常管理,整个生长过程中未施用任何农药。
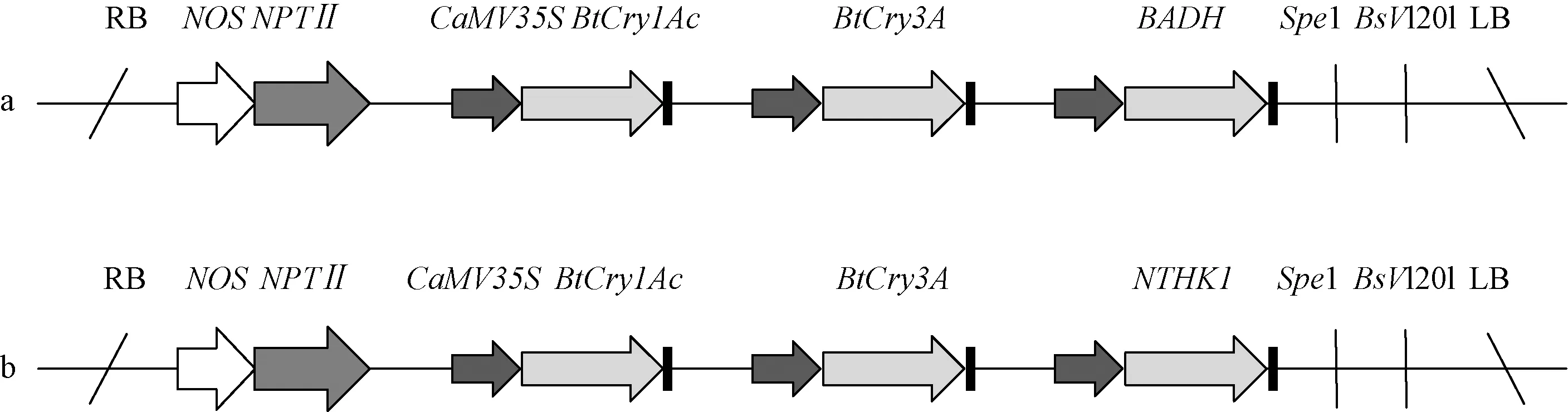
图1 载体结构Fig.1 The structure of vectora:载体V1结构图;b:载体V2结构图。a:The structure of vector V1;b:The structure of vector V2.
1.1.2 测试昆虫 美国白蛾属昆虫纲(Insecta)鳞翅目灯蛾科(Arctiidae),幼虫卵块由北京林业大学陈敏教授提供(2019年7月)。柳蓝叶甲属昆虫纲鞘翅目叶甲科(Chrysomelidae),成虫、幼虫或卵块取自保定满城和河北农业大学苗圃(2019年7月底至8月初)。
1.2 转基因植株的PCR检测
采集2年生转基因苗木当年生的长枝叶片,采用改良的 CTAB法提取各转基因株系及对照植株的基因组DNA。以基因组DNA为模板,分别用Cry1Ac、Cry3A基因的正反引物进行PCR检测,正反引物的DNA序列及目的基因扩增片段见表1。PCR检测采用20 μL的反应体系[ddH2O:12.8 μL;10×PCR Buffer:2 μL;引物R(10 pmol·L-1):1 μL;引物F(10 pmol·L-1):1 μL;DNTP:2 μL;2DNA:1 μL],以含有Cry1Ac和Cry3A基因的质粒DNA为阳性对照,以未转基因的107杨的DNA作为阴性对照。PCR反应程序为:94 ℃预变性10 min;94 ℃变性30 s,50 ℃复性40 s,72 ℃延伸90 s,共30个循环;72 ℃退火5 min。PCR仪购于新加坡Bio-Rad公司,型号为T1000。

表1 引物信息Tab.1 Primer information
1.3 Cry1Ac和Cry3A毒蛋白的ELISA检测
8月下旬于河北农业大学西校区苗圃采集各转基因株系及未转基因欧美杨107杨2年生幼树顶端完全展开的第3、4叶位的叶片,用于Cry1Ac、Cry3A毒蛋白检测。采用美国Agdia公司的Bt-Cry1Ab/1Ac和Bt-Cry3A ELISA试剂盒,分别进行检测,检测方法参照试剂盒说明书进行。检测毒蛋白的阳性对照为试剂盒自带,阴性对照为对照组的107杨,检测结果用 BioRad 550型酶标仪进行测定。蛋白浓度以每g新鲜叶片所含毒蛋白质量(ng)计算。每个处理进行3次生物学重复。
1.4 抗虫性检测
美国白蛾的1龄、2龄幼虫用于检测Cry1Ac毒蛋白的抗虫性。1龄供试幼虫为室内孵化1~2天的初孵幼虫,2龄幼虫在未进行试验前以107杨新鲜叶片饲养,其后用转基因叶片进行抗虫性检测,均设置对照组,饲喂至少量幼虫结茧为止,历时45天。
柳蓝叶甲的1龄、2龄幼虫及成虫用于检测Cry3A毒蛋白的抗虫性,其幼虫饲喂试验方法与美国白蛾大致相同,增加2龄供试幼虫与1龄幼虫进行混龄抗虫性试验,同时增加成虫抗性试验。各试验均以未转基因107杨作为对照组,饲喂至试验幼虫发育成蛹或成虫出现大面积死亡为止,历时20天。
采集各转基因株系和对照组当年生枝条新鲜叶片,将测试昆虫均匀放置于叶片表面,尽量减少对试验昆虫造成损伤,然后将叶片(插于花泥保鲜)安放在透明塑料盒中,盒盖扎上小孔,保证透气性。
试验中,美国白蛾初孵幼虫各株系设置2~4个重复,每处理20~30头;美国白蛾2龄幼虫各株系设置3个重复,每处理10头。柳蓝叶甲初孵幼虫各株系设置4个重复,每处理30头;其余虫期各株系设置3个重复,1龄、2龄混龄幼虫每处理20头,成虫每处理15头。
计算公式如下:
总死亡率=Σ(各重复昆虫死亡数/该重复内测试昆虫总头数)/重复数;
校正死亡率=(转基因株系死亡率-对照死亡率)/(1-对照死亡率);
总增长率=(最后一天的体质量/体长-第1天的体质量/体长)/天数;
取食叶面积率=取食面积/叶片总面积。
2 结果与分析
2.1 转基因植株的PCR检测
利用Cry1Ac和Cry3A基因的特异性引物对6个转基因株系2年生田间苗DNA进行PCR 扩增,以带有目的片段的质粒为阳性对照,未转基因107杨为阴性对照。PCR扩增结果显示,在6个转基因107杨株系DNA中,均能检测到Cry1Ac基因(图2a,特异性条带为546 bp)和Cry3A基因(图2b,特异性条带为667 bp),而对照组均未检测到特异性条带,说明转多基因107杨在生长发育过程中,未发生外源目的基因的丢失。

图2 外源基因的PCR检测Fig.2 PCR amplification of the exogenous genes of various transgenic linesa:Cry1Ac基因的PCR检测;b:Cry3A基因PCR检测。CK+:质粒p09687199;CK-:阴性对照(未转基因植株);M:DL2000 DNA Marker(自上而下为2 000,1 000,750,500,250,100 bp)。ECAA1-3:转Cry1Ac-Cry3A-BADH基因的3个株系;ECAB1-3:转Cry1Ac-Cry3A-NTHK1的3个株系。a:PCR detection of Cry1Ac;b:PCR detection of Cry3A. CK+:Plasmid p09687199;CK-:Control (Untransformed line);M:DL2000 DNA Marker (From up to down,these are 2 000,1 000,750,500,250,100 bp).ECAA1-3:Three lines transformed with Cry1Ac-Cry3A-BADH gene;ECAB1-3:Three lines transformed with Cry1Ac-Cry3A-NTHK1 gene.
2.2 转基因植株的Bt毒蛋白检测
8月下旬采用ELISA法对转多基因107杨叶片中Cry1Ac和Cry3A毒蛋白进行检测,结果(图3)表明,转基因株系均含有2种Bt毒蛋白,但对照组均未检测到。各转基因株系Cry1Ac毒蛋白含量(3.60~20.76 ng·g-1)均明显低于Cry3A毒蛋白含量(3 479.66~7 230.29 ng·g-1),二者相差1 000倍左右。各株系间Cry1Ac毒蛋白含量存在显著差异,株系ECAA1和ECAB3中含量较高,而株系ECAA3和ECAB2中含量较少。显著低于其他株系。各转基因株系中Cry3A毒蛋白含量均较高,且各株系间也存在显著差异,株系ECAA1中毒蛋白含量最高,株系ECAA3与ECAB1中毒蛋白含量较少。在各株系中ECAA1的Cry1Ac和Cry3A毒蛋白含量均是最高的。

图3 转基因107杨不同株系的Cry1Ac和Cry3A毒蛋白含量Fig.3 Content of Cry1Ac and Cry3A toxin protein in different lines of transgenic 107 poplar
2.3 转基因株系的抗虫性检测
2.3.1 对美国白蛾幼虫的抗性 用转多基因107杨饲养美国白蛾1龄幼虫14天,结果如表2所示。各株系之间美国白蛾的致死效果存在显著差异,株系ECAA2、ECAA3和ECAB1对1龄幼虫的致死效果较好,其中ECAB1抗性最高,其校正死亡率为100%。美国白蛾幼虫在前6天死亡个数较多,随后死亡率降低,但在12~14天,幼虫的死亡个数突然增加,这可能与Cry1Ac毒蛋白的积累有关。

表2 转基因株系对美国白蛾1龄幼虫的致死作用①Tab.2 Lethal effect of transgenic lines on the 1st instar larvae of Hyphantria cunea
2.3.2 对美国白蛾的滞育效应 用各转基因株系与对照组107杨饲喂美国白蛾1龄幼虫,图4为饲喂不同天数时美国白蛾的各龄级分布。从图可以明显看出,美国白蛾的发育进程,对照组植株在7天时已全部进入2龄以上,而转基因株系ECAA1、ECAA2和ECAA3中,仍存在很多1龄幼虫,其中株系ECAA2的滞育表现最好。其他株系在7天时均出现3龄幼虫,仅ECAB1的幼虫全部处于2龄阶段。到达13天时,对照组已大部分进入4、5龄,此时转基因株系ECAB1的幼虫已全部死亡,转基因株系ECAA2仍存在1龄幼虫,ECAA1、ECAA3有较多的2龄幼虫。由此可见,转基因株系ECAA1、ECAA2、ECAA3和ECAB1对美国白蛾1龄幼虫有较为明显的抑制其生长发育的作用,即延缓其发育进度;而另外2个株系对美国白蛾生长发育无明显抑制作用。

图4 美国白蛾初孵幼虫发育进度Fig.4 Developmental progress of the first hatched larvae of Hyphantria cunea
用转基因株系和对照饲喂的2龄幼虫,结果见表3、图5和图6,可以看出各转基因株系取食率皆低于对照组107杨。在8月6日至7日和8月14日至15日期间对照组美国白蛾的采食率、体质量及体长均高于转基因各株系,特别是株系ECAA1、ECAA3、ECAB1、ECAB2均明显抑制了美国白蛾2龄幼虫的取食活动,进而影响其生长发育,且各株系间统计学差异显著。由表3可看出,在8月6日至7日时间段内,对照组107杨取食面积最高,而从图6也可以看出转基因107杨株系ECAA3、ECAB1、ECAB2的取食面积较低,由此表明目的基因在受体107杨中得到较好表达。

表3 美国白蛾2龄幼虫平均采食率与死亡数①Tab.3 Average feeding rate and mortality of the 2nd instar larvae of Hyphantria cunea
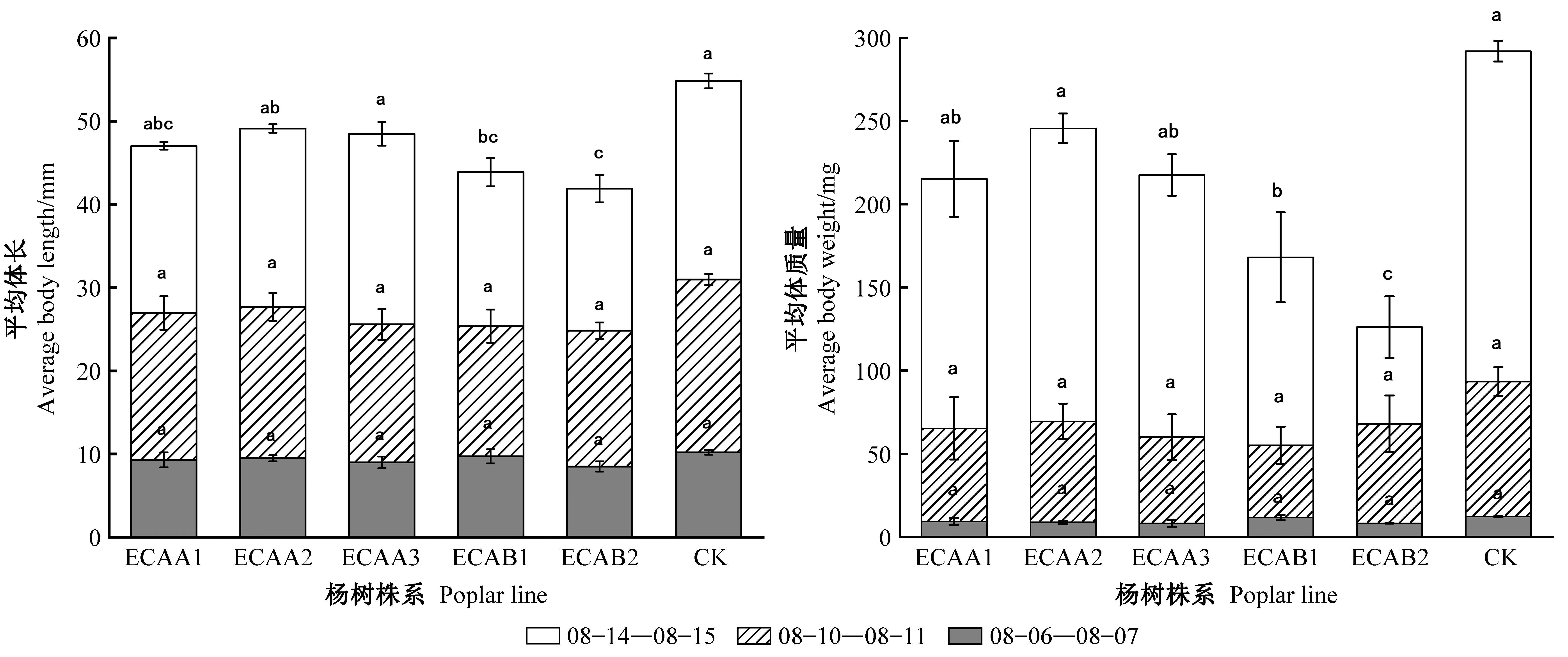
图5 美国白蛾2龄幼虫平均体长、质量Fig.5 Average body length or weight of the 2nd instar larvae of Hyphantria cunea
2.3.3 转基因株系对柳蓝叶甲的抗性检测 图7结果显示,转基因株系ECAA2对柳蓝叶甲无论幼虫还是成虫致死率皆为最低,表现中低抗虫性。除株系ECAA2外,其他转基因株系对柳蓝叶甲1龄幼虫的致死率均达到100%。对1、2龄混龄幼虫的致死率均在85%~100%之间,其中株系ECAB2 和ECAB3的致死率为100%。对于成虫,株系ECAA3、ECAB1、ECAB2和ECAB3的致死效果较好,致死率在90%以上,且6个转基因株系之间存在显著性差异。图8可以看出转基因株系明显抑制了柳蓝叶甲成虫的取食。
3 讨论
植物的大多数性状和生理功能都是通过多基因的协调表达实现的,而转多基因植株可通过协调控制多种外源基因的表达来综合改善植物的性状(初易洋等,2016)。多基因转化指将2个及以上的外源基因转入受体植株中,且转化的多基因可以是不同的抗性基因或是改良植物性状的基因。杨梦悦(2006)将DREB(调节逆境胁迫)、IRT1(提高耐缺铁性)和rolC(毛状根诱导)融合三价基因 (Rirol)转入八棱海棠(Malusrobusta),其转基因植株明显表现出耐盐、矮化和生根力强等性状。王建革等(2006)发现转SacB、vgb、BtCry3A+OC-Ⅰ、调节基因JERF36及报告基因NPTⅡ等基因的库安托杨(Populus×euramericana‘Guariento’)具有抗盐碱能力的同时,又具有Cry3A基因的抗虫性。另外在对Bt基因的研究中发现,当2个Bt基因同时存在时,杨树表现为双抗性,从而扩大抗虫谱,提高抗虫效果(张超等,2019)。
本研究中,采用2年生转多基因107杨株系的叶片进行外源基因表达检测和抗虫试验,显示外源基因仍存在于基因组中,2种Bt毒蛋白均存在,且对美国白蛾和柳蓝叶甲表现双抗性,说明2年生转基因杨树的外源基因依旧稳定表达。李秀芬(2008)在转Bt基因的南林895杨(Populus×euramericana‘Nanlin895’)研究中证明其继代培养4年后仍然携带外源目的基因;陈虞超等(2014)在对9年生抗虫转基因银河杨(Populusalba×P.hopeiensis)的抗虫性检测中发现,该转基因银河杨的外源抗虫基因仍然表达正常。充分证明转多基因植物的外源基因能够保持长时间稳定存在和表达。
本试验中,8月末各转基因株系中Cry1Ac毒蛋白在叶片中含量远远低于Cry3A毒蛋白在叶片中的含量,Cry3A毒蛋白在叶片中含量约是 Cry1Ac毒蛋白的103~104倍。张超等(2019)对1年生转Bt基因107杨毒蛋白的时空表达进行检测,发现Cry1Ac毒蛋白含量在9月份达到最高,Cry3A毒蛋白含量在8月份达到最高;徐丽娜等(2015)认为蛋白质含量的季节性和时空变化是植物体内毒蛋白的积累和其代谢过程所造成的,理论上蛋白含量应增加。但经试验发现转抗虫基因三倍体毛白杨的抗虫性虽表达稳定,但随着树龄的增大Bt基因的表达强度会减弱(李立,2009),具体原因有待研究。
本试验的转化载体V1及V2的启动子均为CaMV35S启动子,4个外源基因分别为Cry3A、Cry1Ac、BADH和NTHK1,其中,Cry1Ac基因处于上游,Cry3A处于下游。试验结果显示,Cry1Ac毒蛋白含量明显低于Cry3A。Xu等(2016)认为在载体中上游Bt基因的表达水平较低;而张冰玉等(2006)认为是由于Cry1Ac基因和Cry3A基因的同源性高,易出现共抑制状况。转多基因工程中存在的基因沉默等问题,成为了影响受体植物外源基因表达稳定性和高效性的巨大障碍,而引起基因沉默的原因大致分为3类:第1种为位置效应引起的基因沉默,可以通过调整目的基因在载体中的排列位置与方向,或是在外源基因的两侧加上MAR结构,显著减少共抑制状况,提高外源基因表达水平的稳定性和效率,如Qiu等(2017)发现2个Bt基因处于相反方向排列时,对2个目的基因的表达均有较强的促进作用;第2种为转录水平的基因沉默,主要是由启动子甲基化引起,Giho等(2012)研究发现转入受体植物本氏烟草(Nicotianabenthamiana)的花椰菜病毒,因35S启动子发生甲基化导致其基因沉默,启动子的甲基化对转基因的遗传稳定性产生负面影响,可使用去甲基化试剂、修饰外源基因的序列或是选取有效促动剂等方法解决转录水平的基因沉默问题;第3种转录后水平的基因沉默,应通过尽量避免或是减少基因之间的同源性等方法解决(李宝健等,2005)。
本试验通过统计幼虫、成虫的致死率、发育进度、虫长和虫质量,筛选出具中高双抗性株系ECAA3和ECAB1,但发现这2个株系的Cry1Ac毒蛋白和Cry3A毒蛋白含量与抗虫性之间无明显规律性,毒蛋白含量与抗虫性关系还有待进一步研究,同时再次验证转基因杨树能够显著毒杀靶标害虫。另外,转基因株系对低龄幼虫有较强致死效果,因低龄幼虫对Bt毒蛋白较为敏感,其作用机理为Bt毒蛋白进入幼虫体内转化成活化了的毒素与肠道上皮的特异受体结合,形成穿孔导致幼虫死亡(Batesetal.,2005);而对高龄幼虫的滞育效果较明显,可能是随着虫龄的增长,虫体的解毒酶活力增加,而抗虫毒蛋白被降解,故死亡率降低,但可以有效地抑制幼虫的体长与体质量的增长和发育速率(田亚坤等,2014)。转多基因库安托杨具有抗旱、抗涝、抗盐碱和抗鞘翅目昆虫的特点(李环,2008),这种转入的多个基因均能在杨树中稳定表达,与本次试验中所选出的高抗株系中多基因稳定表达的结果相似。
抗虫基因转化有诸多优点,不仅可以实现定向改造植株的目标,而且可以扩大林木的抗虫谱,达到一种林木具多抗虫性的目的。但仍存在转化效率低、重复性差和在受体中不能稳定表达等问题,特别是外源基因能否稳定保留在基因组中的问题(Zhangetal.,2016)仍需深入研究。因此,为了使外源基因安全稳定表达、提高林木品质仍需进一步的研究和探讨。同时,本试验中的毒蛋白含量与抗虫性关系还待进一步研究,且未对BADH和NTHK1这2种耐盐基因进行检测和表达分析,未来需要进一步探究。
4 结论
转多基因107杨2年生田间苗各株系的外源基因仍然存在且稳定表达,不同转基因株系间抗虫性差异显著,株系ECAA3对美国白蛾1龄幼虫具有明显抑制其生长发育的作用,株系ECAB1对美国白蛾的致死率最高,ECAA3和ECAB1株系对柳蓝叶甲和美国白蛾均有较高抗性。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17 12:39:32
林业科技(2020年3期)2021-01-21 08:28:46
四川农业科技(2019年5期)2019-07-01 09:46:46
现代园艺(2018年3期)2018-02-10 05:18:38
现代园艺(2017年19期)2018-01-19 02:50:23
现代园艺(2017年23期)2018-01-18 06:58:17
山东林业科技(2016年5期)2016-07-05 00:43:04
浙江柑橘(2016年1期)2016-03-11 20:12:31
江苏农业科学(2015年5期)2015-10-20 20:48:15
江苏农业科学(2015年3期)2015-07-31 14:09:16
