
芹菜素对帕金森模型细胞氧化应激水平的影响及机制的研究
2022-01-17 02:25张跃其胡炳蕾王默然马海强陈学丛
中国比较医学杂志 2021年12期
张跃其,胡炳蕾,王默然,马海强,张 君,陈学丛∗
(1.潍坊市人民医院神经内科,山东 潍坊 261000;2.潍坊市中医院妇科,山东 潍坊 261000;3.潍坊市中医院老年科,山东 潍坊 261000;4.潍坊市中医院骨科,山东 潍坊 261000;5.山东第一医科大学第一附属医院神经内科,济南 250013)
帕金森氏病(Parkinson’s disease, PD)是一种神经退行性疾病,主要表现为静息性震颤、姿势异常、肢体僵硬和运动障碍,60 岁以上的人群中患病率约为1%[1],而未来20 年罹患人数将增加一倍,全世界PD 患者预计将超过1400 万[2],因此,针对PD 治疗措施的研究具有重要的社会及临床意义。PD 的神经化学特征包括活性氧生成、线粒体功能障碍、炎症、错折叠蛋白的积累和泛素蛋白酶体系统功能障碍。 目前研究发现自噬途径调控异常在PD 的发生发展过程中发挥着重要作用[3],其参与了PD 患者中多种病理过程,最终出现清除受损蛋白质和/或细胞器功能异常。
芹菜素(4’、5、7-三羟基黄酮,apigenin, AGN)是存在于水果和蔬菜(例如洋葱,橙子)中的类黄酮的一个子类[4],而类黄酮可以保持黑质纹状体的完整性和功能性[5]。 AGN 在各种细胞类型中具有多种生物活性,例如抗氧化、抗炎和抗肿瘤发生[4]。研究发现AGN 可保护脑神经血管抵抗淀粉样蛋白-β25-35诱导的小鼠神经毒性[6],此外AGN 可预防小胶质细胞介导的炎症毒性,并维持适当的神经胶质神经元相互作用[7]。 AGN 可以作为潜在的神经保护剂来对抗与PD 相关的病理过程[4]。 自噬是细胞降解自身受损细胞器和大分子物质的生理过程,其与PD 的发展密切相关[8],目前尚未有关于AGN 对PD 细胞模型中的神经保护作用及自噬影响的报道。 因此,在本研究中,我们研究了AGN 对MPP+诱发的PD 模型中细胞自噬、氧化应激的影响及神经保护作用,为新的PD 临床治疗措施提供理论基础。
1 材料和方法
1.1 细胞
人神经母细胞瘤SH-SY5Y 细胞购自美国ATCC(ATCC-CRL-2260TM)公司。
1.2 主要试剂与仪器
芹菜素购自Sigma-Aldrich 公司(纯度98%,12806KH);胎牛血清(FBS)以及DMEM 培养基购自美国Gibco 公司;链霉素及青霉素购自美国Invitrogen 公司;CCK-8 试剂盒购自上海碧云天公司;凋亡检测试剂盒购自法国Transgene 公司;Capase-3 兔单克隆抗体、Bcl-2 兔单克隆抗体及Beclin 兔单克隆抗体购自美国Abcam 公司;LC3-II兔单克隆抗体、Bax 兔单克隆抗体、ULK1 兔单克隆抗体、β 单克隆抗体-鼠单克隆抗体购自美国CST公司;二抗及ECL 发光试剂盒购自上海碧云天公司;PVDF 膜购自美国Millipore;蛋白电泳系统购自北京六一仪器厂;酶标仪购自美国Bio-Rad;激光共聚焦显微镜购自德国Leica 公司;流式细胞仪购自美国BD(FACSAria III)公司。
1.3 实验方法
1.3.1 细胞培养
SH-SY5Y 细胞培养于含10%胎牛血清、1%青霉素和链霉素的DMEM 培养基中。 细胞培养箱孵育条件为37℃、5% CO2。 细胞密度90%时胰酶消化传代,按照每孔2×103个细胞接种到96 孔板进行后续相关实验。
1.3.2 CCK-8 筛选AGN 有效浓度
以CCK-8 法检测细胞活力。 向每孔加入CCK-8 溶液20 μL,将培养板在培养箱中孵育2 h,用酶标仪测定450 nm 处的吸光度,计算细胞活性。 实验重复3 次。 本部分实验分6 组:①对照组:DMEM 培养基;②MPP+组:DMSO(浓度0.5%)+2.5 mmol/L 的MPP+诱导细胞24 h;③A 组:加入AGN 孵育细胞2 h 后再加入MPP+,AGN 终浓度10 μmol/L,MPP+终浓度2.5 mmol/L;④B 组:AGN 终浓度20 μmol/L,MPP+终浓度2.5 mmol/L;⑤C 组:AGN 终浓度40 μmol/L,MPP+终浓度2.5 mmol/L;⑥D 组:AGN 终浓度100 μmol/L,MPP+终浓度2.5 mmol/L。
1.3.3 细胞凋亡率检测
确定AGN 有效浓度后,将细胞分为3 组:对照组、MPP+组及芹菜素有效剂量组。 采用AnnexinV与PI 共染法检测细胞凋亡。 将细胞培养于96 孔板,每孔加入Annexin V-FITC 结合液,轻轻重悬细胞,加入5 μL Annexin V-FITC,轻轻混匀,加入10 μL 碘化丙啶染色液轻轻混匀,室温(25℃)避光孵育20 min,随后置于冰浴中,用流式细胞仪FITC 和PI 通道检测细胞凋亡。 实验重复3 次。
1.3.4 蛋白质免疫印迹检测
收集细胞经匀浆后,经RIPA 法裂解提取蛋白。超声裂解仪裂解10 次后低温离心机中离心30 min,取上清并用BCA 法定量蛋白浓度,蛋白经SDSPAGE 电泳后电转至PVDF 膜,脱脂牛奶摇床上封闭1 h,加入一抗后置4℃恒温摇床过夜。 洗膜3 次后室温孵育二抗1 h,利用化学发光试剂盒显影蛋白条带,采用Image-J 软件分析定量蛋白条带,β-actin为内参。 实验重复3 次。
1.3.5 氧化应激检测
分别收集3 组细胞到离心管内,离心后超声波破碎细胞,4℃离心10 min 后取上清,采用丙二醇(malonaldehyde, MDA)检测试剂盒、超氧化物歧化酶(superoxide dismutase, SOD)检测试剂盒和谷胱甘肽过氧化物酶(glutathione peroxidase, GPX)检测试剂盒用来检测各组中氧化应激相关底物的改变,分光光度计检测吸光度,具体方法参照试剂盒说明书。 每组实验均重复3 次。
1.3.6 GFP-LC3 质粒转染
将SH-SY5Y 细胞按照每孔2×105个接种6 孔板,每孔转染2 μg GFP-LC3 质粒,孵育6 h 更换新培养基,加入相应的药物处理24 h 后进行GFP-LC3斑点计数分析。 在胞浆及胞核中GFP-LC3 散在分布的细胞为自噬阴性细胞,出现GFP-LC3 斑点聚集的为自噬阳性细胞,各组计数至少100 个自噬阳性细胞中的GFP-LC3 斑点。 于激光共聚焦显微镜下扫描转染细胞。 实验重复3 次。
1.4 统计学方法
采用SPSS 21 软件进行统计分析。 计量资料用平均数±标准差(±s)表示,多样本均数的比较采用ANOVA 分析检验,事后各组间的两两比较采用Tukey 法,P<0.05 为差异有统计学意义。
2 结果
2.1 AGN 对MPP+诱导的SH-SY5Y 细胞的保护作用
本实验首先通过CCK-8 法对AGN 的有效保护浓度进行筛选。如图1所示, 20 μmol/L、40 μmol/L、100 μmol/L 三种浓度的AGN 能显著降低MPP+诱导的细胞毒性损伤,细胞活性明显改善(相较MPP+组,P值分别<0.05、<0.01、<0.01)。AGN 浓度在40 μmol/L 时保护作用最强,选择40 μmol/L 作为最佳药物保护浓度进行后期实验研究。
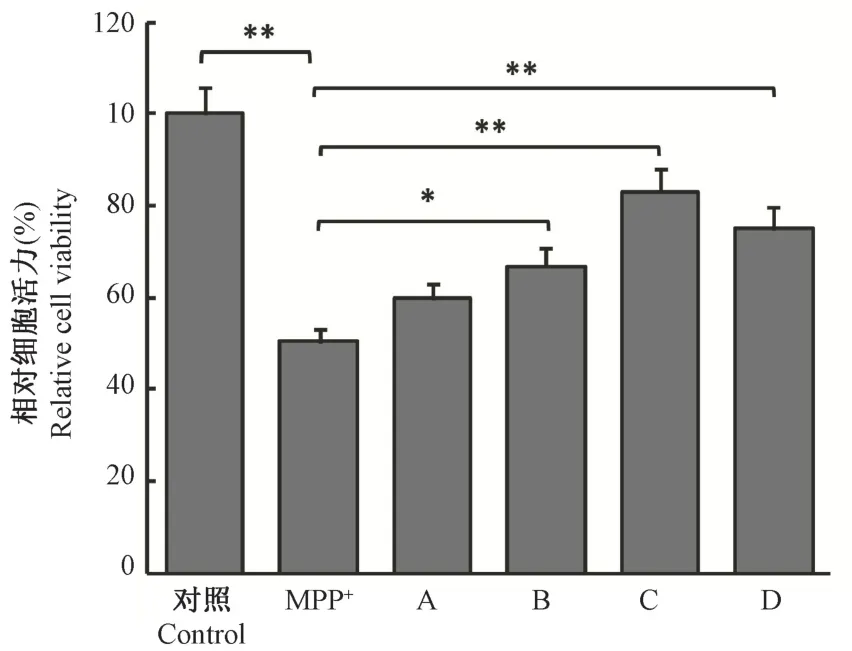
图1 不同剂量AGN 对MPP+诱导的SH-SY5Y细胞活性的影响Note. Compared with the MPP+ group, ∗P <0.05, ∗∗P <0.01.Figure 1 Effects of different doses of AGN on the viability of SH-SY5Y cells induced by MPP+
2.2 AGN 对MPP+诱导模型细胞凋亡及凋亡相关蛋白的影响
后续实验分为3 组:对照组、MPP+组、AGN+MPP+组(AGN 40 μmol/L)。 如图2A、2B 所示,MPP+组细胞凋亡率为11.03%,较对照组升高(P<0.01);AGN+MPP+组细胞凋亡率为6.12%,较MPP+组下降(P<0.05)。 随后我们比较了各组Caspase3 及Bcl-2、Bax 蛋白的表达。 如图2C、2D 所示,相较于对照组,MPP+组细胞Caspase3 表达明显升高,Bcl-2/Bax 值明显降低(P<0.01);而AGN+MPP+组相较MPP+组,Caspase3 表达降低,Bcl-2/Bax 值明显升高(P<0.01)。 以上结果表明AGN 可改善MPP+诱导的细胞凋亡。
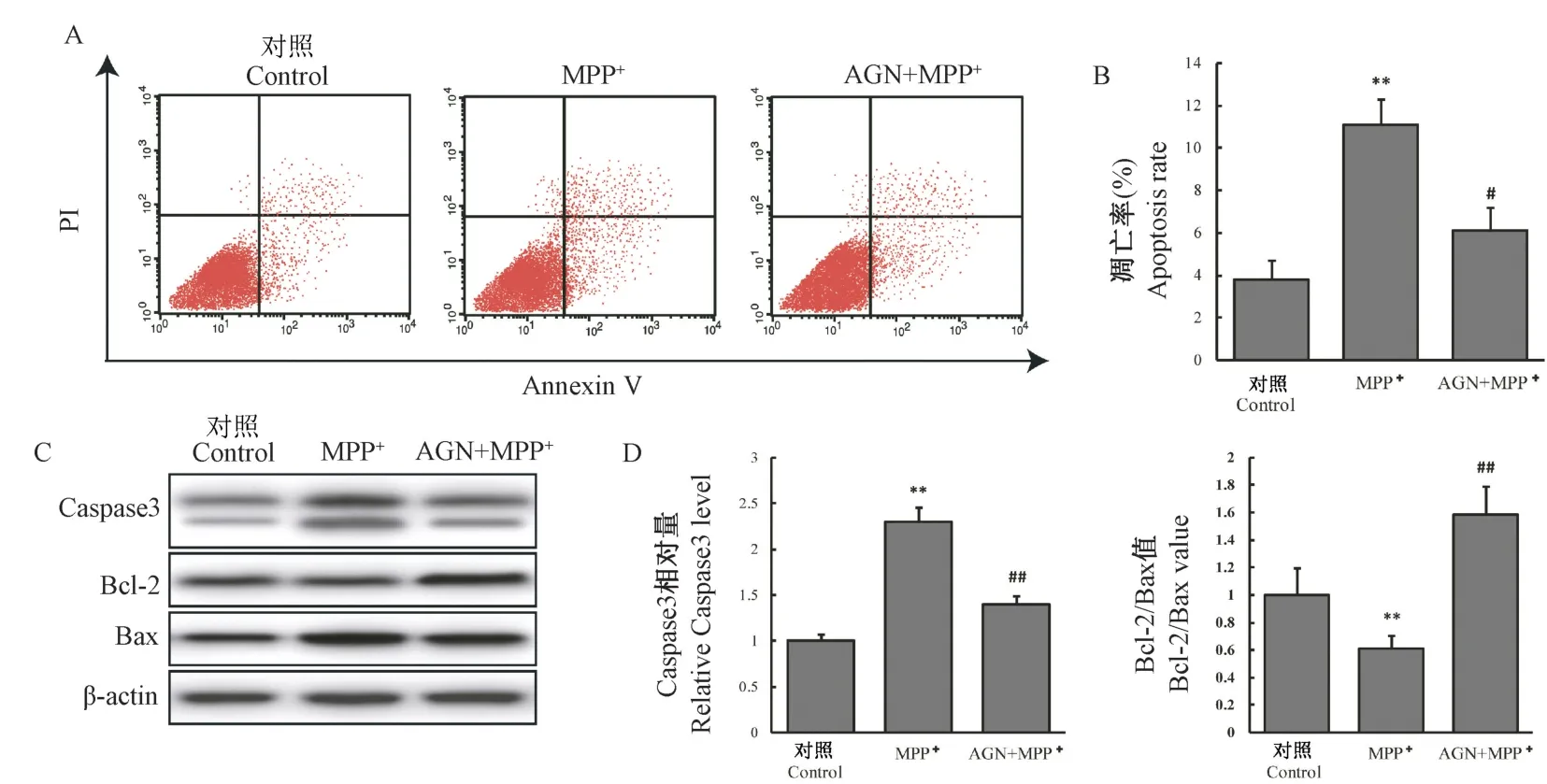
图2 AGN 对细胞模型凋亡及凋亡相关蛋白的影响Note. A, Apoptotic rate of each group was detected by flow cytometry. B, Comparison of cell apoptosis rate in each group,Compared with control group,∗∗P<0.01. Compared with MPP+ group, #P<0.05. C, Western blot was used to detect the expression of Caspase3, Bcl-2 and Bax protein in each group. D, Comparison of the relative level of Caspase3 and Bcl-2/Bax value in each group. Compared with Control group,∗∗P<0.01. Compared with MPP+ group, ##P<0.01.Figure 2 Effect of AGN on cell apoptosis and apoptosis-related proteins
2.3 AGN 对MPP+诱导模型细胞氧化应激的影响
进一步检测各组中氧化应激相关底物表达及酶活性。 和对照组相比,MPP+促进了MDA 表达(P<0.01),而与MPP+组比较AGN 可抑制MDA 的表达(P<0.05)。 和对照组相比,MPP+组细胞GPX和SOD 活性降低,而与MPP+组比较AGN 能恢复GPX 和SOD 的活性(P<0.05)。 见图3。
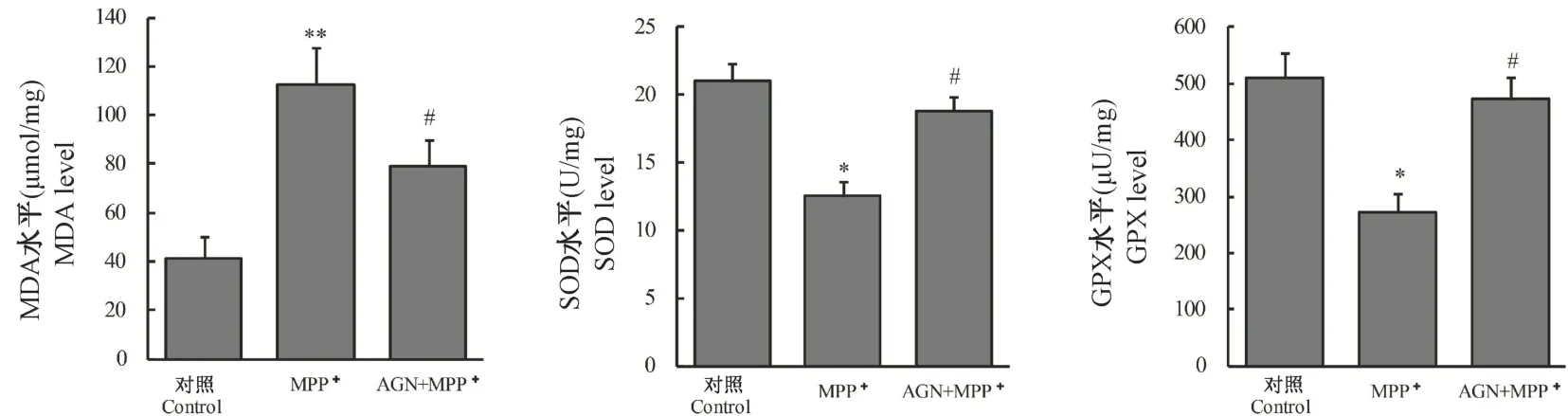
图3 AGN 对模型细胞氧化应激的影响Note. Compared with Control group, ∗P<0.05, ∗∗P<0.01. Compared with MPP+ group, #P<0.01.Figure 3 Effect of AGN on oxidative stress of model cells
2.4 AGN 对MPP+诱导模型细胞GFP-LC3 斑点形成及自噬蛋白的影响
为进一步明确AGN 对MPP+诱导模型细胞自噬的影响,我们进行了GFP-LC3 斑点形成实验。GFP-LC3 斑点作为胞浆内的自噬标志物,可以反映出自噬体对LC3 蛋白的募集情况。 将GFP-LC3 质粒转染SH-SY5Y 细胞,结果显示MPP+组细胞内GFP-LC3 斑点较对照组显著增多(P<0.01),而AGN+ MPP+组细胞内GFP-LC3 斑点较MPP+组显著减少(P<0.05)。 见图4A、4B。 随后我们观察了LC3、Beclin-1 及ULK-1 蛋白的表达水平。 MPP+处理后,LC3-II/LC3-I 比值及Beclin-1、ULK1 表达较对照组显著增加(P值均<0.01);而对比MPP+组,AGN+MPP+组LC3-II/LC3-I 比值及Beclin-1、ULK1表达显著降低(P值均<0.05),见图4C、4D。 以上结果进一步证明了AGN 可显著降低MPP+诱导模型细胞的自噬体形成及自噬水平。
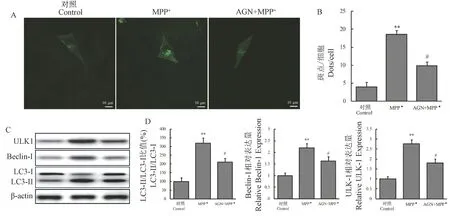
图4 AGN 对GFP-LC3 斑形成及自噬蛋白的影响Note. A, GFP-LC3 spots in each group observed by the confocal laser microscope. B, Comparison of GFP-LC3 spots in each group.Compared with control group, ∗∗P<0.01. Compared with MPP+ group, #P<0.05. C, Western blot was used to detect the expression of LC3, Beclin-1 and ULK1 proteins in each group. D, Comparison of the LC3-II/LC3-I ratio and the expression of Beclin-1, ULK1 in each group. Compared with Control group, ∗∗P<0.01. Compared with MPP+ group, #P<0.05.Figure 4 Effect of AGN on GFP-LC3 plaque formation and autophagy protein
3 讨论
PD 病理特征是黑质中的多巴胺能神经元逐渐丧失,随后纹状体中多巴胺水平下降,当神经元减少50%~70%时会表现出临床症状[9]。 目前研究证实氧化应激是PD 致病机制的重要因素,导致线粒体稳态失调,进而出现细胞凋亡、神经元减少[10]。在本研究我们发现AGN 能减轻MPP+诱导的SHSY5Y 细胞凋亡,增强抗氧化能力,并能降低细胞自噬水平,提示AGN 对帕金森模型细胞的保护作用与降低细胞自噬水平相关。
氧化应激在PD 中的神经变性改变起着关键作用,可促炎性细胞因子、活性氧(ROS)、诱导型一氧化氮合酶(iNOS)以及超氧化物的释放而加速神经退行性过程,加剧了多巴胺能神经元的损伤[10-11]。PD 患者纹状体和黑质致密部的尸检表明,氧化应激会损害脂质、蛋白质和DNA 功能,同时降低SOD、过氧化氢酶和GPX 的含量[12-14]。 SOD 和GPX 是清除活性氧自由基的主要酶,其水平降低会使细胞更容易受到氧化损伤[13]。 PD 患者GPX、SOD 显著耗竭,蛋白质及DNA 氧化水平显著增加[15]。 人体解剖数据表明,氧化应激的启动最终导致神经元死亡[14]。 另一项临床研究表明,PD 患者黑质多巴胺能神经元的核因子kB(NF-κB)的核易位倍数增加,并且NF-κB 由氧化应激触发,放大了炎症和凋亡程序的过程[16]。 因此,具有神经保护、抗氧化效能的药物具有减缓神经变性进程的潜力,有助于促进神经元细胞存活,从而减轻疾病表现。 在各种PD 实验模型中,用抗炎药抑制神经炎症可减轻多巴胺能神经退行性变[17]。
在本研究中,AGN 作用下SOD、GPX、MDA 水平的改变证实了AGN 对PD 模型细胞具有很好抗氧化应激作用。 AGN 是多种植物中存在的多酚类物质,已知其具有多种生理益处,尤其是在清除自由基、改善认知障碍、改善学习和记忆方面[18]。 有研究对AGN 在转基因帕金森果蝇模型中的治疗效能进行了检测,发现其可改善果蝇的运动能力[19]。 对小胶质细胞的研究证明,AGN 对炎性介质具有抑制作用,表明其在神经退行性疾病中可能具有神经保护作用[18]。 另一项研究表明AGN 能促进成年小鼠的神经发生,提示其具备神经保护和神经营养能力[20]。
自噬是一种降解和回收细胞组分的溶酶体依赖性的内源性过程,目前已经证实其与PD 的发生发展密切相关[3]。 在本研究中,AGN 在帕金森模型细胞中可抑制LC3-II/LC3-I 比值及Beclin-1、ULK-1,并抑制LC3 斑点形成,减轻自噬水平。 MPP+是线粒体呼吸链复合体的抑制剂,MPP+可模拟PD 病理过程,其诱导的氧化损伤类似于PD 患者大脑中发现的氧化损伤,损害细胞线粒体功能,而这会触发细胞自噬水平[21]。 此外,在PD 患者中线粒体膜上a-Syn 过度积累可导致心磷脂暴露于线粒体外膜,从而募集LC3 连接蛋白,诱导线粒体自噬[22]。Zhu 等[23]在2012 年的一项研究发现,加入ERK 信号通路特异性抑制剂可抑制MPP+导致的自噬水平升高。 以UO126 抑制MPP+所致的自噬水平升高,会出现线粒体膜电位水平升高,线粒体活性氧水平降低[24]。 半胱氨酰白三烯受体拮抗剂可同步抑制花生四烯酸相关的炎性应激及自噬水平,从而保护神经元,发挥对全脑缺血的治疗作用[25]。 与上述研究类似,本研究中MPP+可增强自噬及氧化应激水平,而AGN 可抑制自噬及氧化应激水平。 UNC51样激酶(ULK1)是启动自噬级联反应的丝氨酸/苏氨酸激酶。 研究发现,miR-132-5p 可靶向抑制ULK1,改善细胞活力并减少MPTP 诱导的SHSY5Y 细胞凋亡[26],百可利能抑制ULK1 蛋白过度活化,减轻自噬水平,使帕金森模型小鼠纹状体中酪氨酸羟化酶表达上调,改善小鼠的运动障碍[27]。另一项研究表明,在MPP+处理的MN9D 细胞中ULK1 表达增加,而敲低ULK1 则增加了神经元细胞的活力[28]。 本研究中AGN 抑制了MPP+诱导的ULK1 表达增高,结合前述研究结果分析,AGN 可能是通过调控自噬通路影响了线粒体的氧化应激,进而发挥细胞保护作用。
综上,我们的结果表明,AGN 能缓解MPP+诱导的氧化应激,减轻细胞凋亡,降低细胞自噬水平。本研究揭示了AGN 可能通过影响自噬来降低氧化应激水平,从而减弱了MPP+介导的毒性,发生细胞保护作用。 但由于氧化应激的复杂性及自噬调控的多样性,AGN 调控自噬及氧化应激的具体分子机制仍需我们进一步研究,为PD 的转化治疗应用提供更多理论依据。
猜你喜欢
实用手外科杂志(2022年2期)2022-08-31
现代临床医学(2022年1期)2022-02-12
数学物理学报(2021年4期)2021-08-30
新世纪智能(数学备考)(2020年10期)2021-01-04
世界科学技术-中医药现代化(2020年2期)2020-07-25
中成药(2017年12期)2018-01-19
中国交通信息化(2017年8期)2017-06-06
Coco薇(2015年5期)2016-03-29
西南军医(2016年6期)2016-01-23
癌变·畸变·突变(2015年3期)2015-02-27
