
水稻白条纹转绿突变体wsl887的鉴定和基因定位
2022-01-04 01:10鄢小青陈能刚余显权
核农学报 2021年11期
鄢小青 陈能刚 李 欢 陈 锋 宋 泽 余显权
(1 贵州大学农学院,贵州 贵阳 550025;2 贵州省农业科学院农作物品种资源研究所,贵州 贵阳 550006)
水稻叶色突变体的变异类型十分丰富,一般根据苗期不同的叶色表现,可将叶色突变体划分为转绿型、白化、黄化、条纹、斑马纹以及黄色斑点共6种类型[1]。水稻叶色突变体是研究植物叶绿体发育、光合作用和光形态建成等生物学过程分子调控机制的理想材料。其中温敏型转绿型突变体的叶色突变表型受温度诱导,在自然条件下转绿型突变体的产量性状不受负面影响,可用作水稻杂交育种中的标记性状。因此,对水稻转绿型叶色突变的鉴定及基因定位,可以为水稻高光效育种提供理论依据,也可以作为标记性状应用于水稻不育系繁殖和杂交制种等领域。水稻叶色表型大多数为一对隐性核基因控制,近年来在水稻中克隆得到与叶色相关的基因已超过120个,其中白化转绿基因有51个,主要涉及光合色素代谢途径及叶绿体发育调控[2]。转绿突变体的表型变化主要受到外界温度或生育进程的调控[3],如V2[4]、WSL[5]、WSL3[6]、WSL4[7]、WSL6[8]、WSL9[9]和TSV3[10]等的突变表型受外界温度调控;Rey(k2)[11]、WGL2[12]、GRA78[13]等的突变表型受生育进程调控。V2编码一个鸟苷酸激酶,参与质体的转录和翻译系统的建立[4],WSL编码一个PLS亚家族的PPR蛋白,参与叶绿体基因RNA剪切[5];WSL3编码一个质体RNA聚合酶的非核心亚基OsPAP1/OspTAC3,影响叶绿体的早期发育[6];WSL4编码一个属于P亚家族的PPR蛋白,对水稻叶绿体RNA Ⅱ内含子剪接至关重要[7];WSL6编码一个定位于叶绿体中的era型鸟苷-5′-三磷酸(era-type granosine-5′-triphosphate(GTP)-binding protein)结合蛋白[8];WSL9编码一种具有HNH结构域的新蛋白[9];TSV3编码一个类ObgGTPase蛋白,参与调控叶绿体50S核糖体大亚基的生物合成[10]。Rey(k2)编码一个磷酸果糖激酶B型叶绿体蛋白,参与核质信号的传递[11];WGL2编码一种保守的核糖体蛋白,是水稻叶绿体发育的重要组成部分[12];TSV3编码一个类ObgGTPase蛋白,参与调控叶绿体50S核糖体大亚基的生物合成[12];GRA78编码一个定位于叶绿体的硫氰酸合成酶[13]。尽管已经克隆了很多水稻转绿型叶色突变基因,但其分子调控机制尚不明确,因此有必要定位和克隆更多的水稻叶片转绿型叶色突变基因,为系统阐明转绿型叶色突变的分子机制奠定基础。
本研究利用60Co辐射诱变贵州地方稻种资源桥港珍珠米,获得新的温敏型白条纹转绿突变体wsl887,对该突变体在自然环境中的表型特征和相关农艺性状进行鉴定,并进行不同温度条件下的表型变化、叶绿体超微结构观察以及相关基因表达分析、基因定位等研究,旨在为目的基因的克隆及功能分析奠定基础。
1 材料与方法
1.1 试验材料
利用贵州地方粳稻资源桥港珍珠米通过60Co辐射诱变获得白条纹转绿叶色突变体wsl887,经过连续多代自交,白条纹转绿性状稳定遗传。2019年3月在海南三亚海螺村贵州水稻南繁育种基地以桥港珍珠米和籼稻R1638为父本,分别与wsl887杂交获得F1代(wsl887/桥港珍珠米和wsl887/R1638)种子,同年4月在贵阳(贵州省农作物品种资源研究所水稻试验田)种植F1代,F1自交构建F2遗传群体和F2定位群体。
1.2 表型及农艺性状分析
分别在不同温度(20、25和30℃)的光照培养箱(12 h光照/12 h黑暗)中培养野生型和突变体wsl887,观察幼苗表型。于2019年4月20日在贵阳试验田分别种植wsl887和野生型亲本桥港珍珠米,5月25日移栽大田,设3次重复,每重复种植40株,株间距20 cm×30 cm。在成熟后从种植小区中随机取5株进行农艺性状的调查。
1.3 光合色素含量测定
在光照培养箱(12 h光照/12 h黑暗)中培养野生型和突变体wsl887幼苗,在不同温度(20、25和30℃)条件下萌发后20 d,选择不同温度条件下,野生型和突变体的全展叶,分别称取新鲜叶片0.2 g,剪碎后放入10 mL 95%乙醇中,4℃避光放置48 h,期间多次手动摇匀。将浸泡后的提取液移入25 mL 容量瓶中,以95%乙醇定容。使用722N型分光光度计(上海菁华科技仪器有限公司)测量665、649和470 nm下的吸光值。按Lichtenthaler[14]的方法,计算叶片单位鲜重的叶绿素(Chl)和胡萝卜素(Caro)含量。计算公式如下:
Chla=13.95×A665-6.88×A649
Chlb=24.96×A649-7.32×A665
Caro=(1000×A470-2.05×Ca-114.8×Cb)/245
叶绿体色素含量=C×V/[W×1000]
式中,C为色素浓度(mg·L-1)、V为提取液体积(L)、W为样品鲜重(g)。
1.4 叶肉细胞超微结构观察
分别选取在光照培养箱(12 h光照/12 h黑暗)中培养不同温度(20和30℃)条件下萌发后20 d的野生型和突变体wsl887幼苗的全展叶,观察叶肉细胞叶绿体超微结构。材料固定时确定取材部位,尽量减小牵拉、挫伤与挤压等机械损伤,组织体积一般不超过1 mm×1 mm×1 mm,迅速投入电镜固定液固定,并用真空泵抽气直至沉底,室温放置2 h,然后转入4℃冰箱。0.1 mol·L-1磷酸缓冲液PBS(pH值7.4)漂洗3次,每次15 min;用1%的锇酸0.1 mol·L-1磷酸缓冲液PBS(pH值7.4)室温(20℃)固定5 h。0.1 mol·L-1磷酸缓冲液PBS(pH值7.4)漂洗3次,每次15 min;组织依次入30%~50%~70%~80%~90%~95%~100%~100%酒精进行脱水,每次1 h。无水乙醇∶丙酮=3∶1,0.5 h,无水乙醇∶丙酮=1∶1,0.5 h,无水乙醇∶丙酮=1∶3,0.5 h,丙酮1 h;置于丙酮∶812包埋剂=3∶1,2~4 h,丙酮:812包埋剂=1∶1,渗透过夜,丙酮∶812包埋剂=1∶3,2~4 h,纯812包埋剂5~8 h,将纯812包埋剂倒入包埋板,将样品插入包埋板后37℃烤箱过夜;用60℃烤箱聚合48 h;用超薄切片机切片(60~80 nm);铀铅双染色(2%醋酸铀饱和酒精溶液,枸橼酸铅,各染色15 min),切片室温干燥过夜;在hitachi HT7700透射电子显微镜(日立高新技术公司,日本)下观察,采集图像分析。
1.5 DNA的提取和PCR扩增
水稻新鲜叶片DNA提取按改良的十二烷基硫酸钠(sodium dodecyl sulfate, SDS)法提取[15],分别提取亲本、混池及F2群体的基因组用于基因定位。PCR体系25 μL,包括T3 Super PCR Mix 21 μL、DNA模板2 μL和上下游引物各1 μL。PCR程序为:98℃预变性2 min;98℃变性10 s,55~60℃退火10 s,72℃延伸10 s, 30个循环;72℃终延伸2 min。
1.6 总RNA的提取与qRT-PCR
选取不同温度下野生型和突变体wsl887叶片相同部位提取植物总RNA,采用宝生物工程(大连)有限公司的RNAiso Plus试剂盒提取总RNA;采用北京擎科生物GoldenstarTMRT6 cDNA Synthesis Kit Ver.2试剂盒进行反转录;采用北京擎科生物的2×T5 Fast qPCR Mix(SYBR Green Ⅰ)试剂盒进行荧光定量PCR。在Applied Biosystems ABI 7500荧光定量PCR仪(美国Applied Biosystems)上进行qRT-PCR,以OseEF-1α为内参基因,采用2-ΔΔCT法分析基因的相对表达量,每个样品重复3次。所有qRT-PCR引物由成都擎科梓熙生物技术有限公司合成。
选取在20和30℃条件下萌发生长20 d的野生型和wsl887幼苗叶片,对其光合色素、叶绿体发育、光合作用和线粒体电子传递相关基因的表达水平进行qRT-PCR分析(表2)。光合色素合成相关基因:CHLD、CHLH、CHLI和CHLG;光合作用相关基因:psaA/psbA(编码PSI/PSII反应中心蛋白)、rbcS/rbcL(编码Rubisco小亚基/大亚基)、CAB1R/CAB2R(编码PSII光捕获叶绿素a/b结合蛋白),其中CAB1R、CAB2R和rbcS属于细胞核的编码基因;psaA、psbA和rbcL属于细胞质的编码基因;线粒体电子传递复合体基因:编码线粒体电子传递复合体Ⅰ的基因nad2、nad4和nad5,编码线粒体电子传递复合体Ⅲ的基因cob,编码线粒体电子传递复合体Ⅳ的基因cox1、cox2和cox3,编码线粒体电子传递复合体Ⅴ的基因atp6和atp8。
1.7 基因定位
采用混合群体分离分析法(bulked segregant analysis, BSA)法[16]定位目标基因,从wsl887/R1638的F2群体中选取10株正常绿色的单株和10株叶片呈现白条纹的单株,分别等量混合成正常基因池和突变基因池。参照水稻基因组的简单重复序列(simple sequence repeats, SSR)分子标记,利用籼稻9311和粳稻日本晴基因组间的差异,寻找多态性较好的SSR分子标记。通过3%的琼脂糖凝胶进行电泳,筛选在突变体wsl887与R1638间的多态性分子标记。用多态性好的分子标记分析正常基因池与突变基因池之间的多态性,找到目标基因连锁的分子标记后用F2群体中的白条纹单株进行验证,待确定该分子标记连锁后,进一步寻找更多的多态性分子标记进行精细定位。
2 结果与分析
2.1 表型与农艺性状鉴定
在大田条件下,wsl887在幼苗期呈现白条纹表型(图1-A、B),随着植株生长开始转绿,新长出的叶片为正常绿色。在成熟期,wsl887与野生型相比形态特征无明显差异(图1-C),主要农艺性状调查发现,wsl887的株高、有效穗、穗长、结实率和千粒重较野生型无显著差异(表1)。
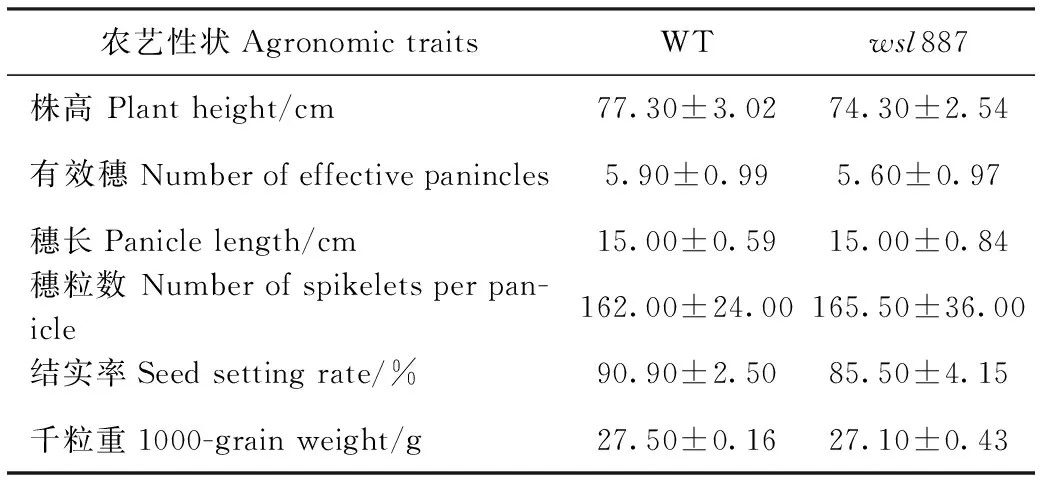
表1 野生型(WT)和突变体wsl887主要农艺性状比较Table 1 Comparison of major agronomic traits between WT and mutant wsl887
2.2 不同温度下叶肉细胞超微结构观察
透射电镜观察发现野生型在20和30℃条件下,叶肉细胞中叶绿体含量丰富,叶绿体形态饱满;而wsl887在20℃条件下,白化叶片的叶肉细胞中没有叶绿体或者叶绿体发育畸形,含较多的囊泡状结构,线粒体结构与野生型无明显差异;wsl887在30℃条件下,叶片为正常绿叶,叶肉细胞中叶绿体发育完整,数量与野生型无明显差异。表明在低温(20℃)条件导致wsl887叶绿体发育异常,表现为白化表型;在高温条件(30℃)下,wsl887叶绿体的数量与叶绿体结构的发育与野生型无显著差异,表现为正常绿叶表型。
2.3 不同温度光合色素的变化
不同温度处理试验中发现,在不同温度(20、25和30℃)光照培养箱中萌发20 d的秧龄突变体wsl887的幼苗间存在叶色差异。在20℃条件下,wsl887叶片呈现完全白化;25℃条件下,wsl887呈现带有白色条纹的叶片;在30℃条件下,wsl887叶色与野生型无明显差异(图4-A、B、C)。进一步对不同温度处理的野生型和wsl887的光合色素进行测定发现(图4-D、E、F),与野生型相比,在20℃条件下,wsl887叶片的总叶绿素、叶绿素a、叶绿素b和类胡萝卜素含量降幅最大,均达到极显著差异水平,分别降低至野生型的6.08%、5.87%、6.69%和12.48%;在25℃条件下,wsl887叶片的总叶绿素、叶绿素a、叶绿素b和类胡萝卜素含量降幅也较大,均达到极显著差异水平,分别降低至野生型的53.39%、53.45%、53.19%和59.96%;在30℃条件下,与野生型相比,wsl887叶片的总叶绿素、叶绿素a、叶绿素b和类胡萝卜素含量与野生型无显著差异。
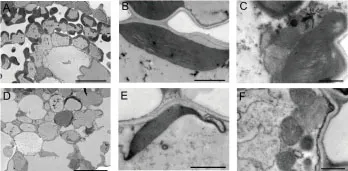
注:A、B、C:野生型叶肉细胞; D、E、F:突变体wsl887叶肉细胞。A、D: Bar=20 μm; B、E: Bar=2 μm; C、F: Bar=1 μm。Note: A、B、C:Mesophyll cells structure of wild type (WT). D、E、F: Mesophyll cells structure of wsl887 mutant. A、D: Bar=20 μm. B、E: Bar=2 μm. C、F: Bar=1 μm.图2 20℃温度下野生型(WT)和突变体wsl887叶肉细胞中叶绿体超微结构Fig.2 Ultrastructure of chloroplasts in mesophyll cells of the wild type (WT) and wsl887 mutant under 20℃ temperature conditions

注:A、B、C:野生型叶肉细胞; D、E、F:突变体wsl887叶肉细胞。A、D: Bar=20 μm; B、E: Bar=2 μm; C、F: Bar=1 μm。Note: A、B、C:Mesophyll cells structure of wild type (WT). D、E、F: Mesophyll cells structure of wsl887 mutant. A、D: bar=20 μm. B、E: Bar=2 μm. C、F: Bar=1 μm.图3 30℃温度下野生型(WT)和突变体wsl887叶肉细胞中叶绿体超微结构Fig.3 Ultrastructure of chloroplasts in mesophyll cells of the wild type (WT) and wsl887 mutant under 30℃ temperature conditions
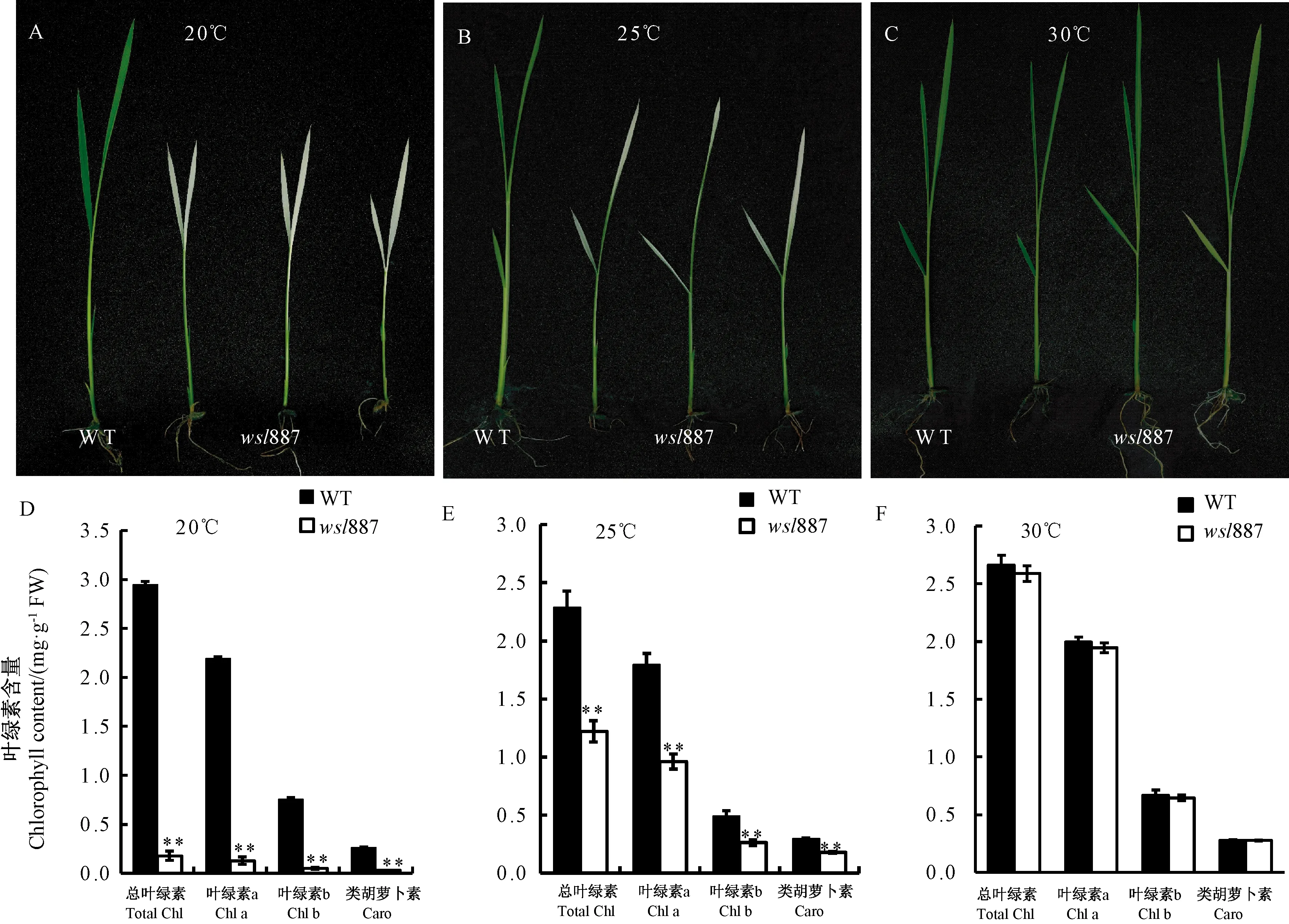
注: **: 在 0.01水平上差异显著。Note: **: Significant difference at 0.01 level.图4 不同温度下野生型(WT)和突变本wsl887幼苗表型及光合色素含量Fig.4 Phenotype and photosynthetic pigment content of the wild type (WT) and wsl887 mutant seedlings under different temperature conditions
2.4 光合色素合成、光合作用和线粒体电子传递复合体相关基因的表达分析
由于wsl887光合色素含量下降,叶绿体形态结构异常,推测在低温条件下,wsl887中叶绿素合成、叶绿体发育、光合作用以及线粒体电子传递相关基因(表2)的表达也受到不同的影响。由图5可知,在20℃条件下与野生型相比,wsl887叶片的叶绿素合成途径相关基因(CHLD、CHLH、CHLI、CHLG)的表达无显著差异;光合作用相关基因psaA、psbA、rbcS、rbcL、CAB1R和CAB2R的表达量不同程度降低,均达到显著水平;线粒体电子传递复合体基因cob、nad2、nad4和nad5、cox1和cox2的表达量显著增加,cox3和atp6的表达量无显著差异,atp8的表达量显著降低。说明在低温条件下,wsl887的呼吸作用增强。
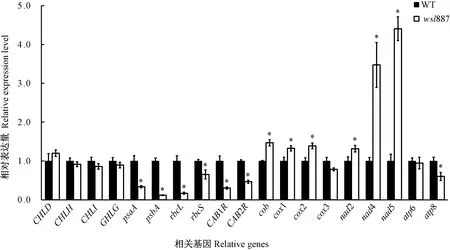
注:*表示在 0.05 水平上差异显著。下同。Note: * indicates significant difference at 0.05 level. The same as following.图5 20℃条件下野生型(WT)及突变体wsl887叶片中光合色素代谢、光合作用和线粒体相关基因的表达分析Fig.5 Expression analysis of genes in photosynthetic pigment metabolism, photosynthesis and mitochondrial genes of the wild type (WT) and wsl887 mutant leaves under 20℃
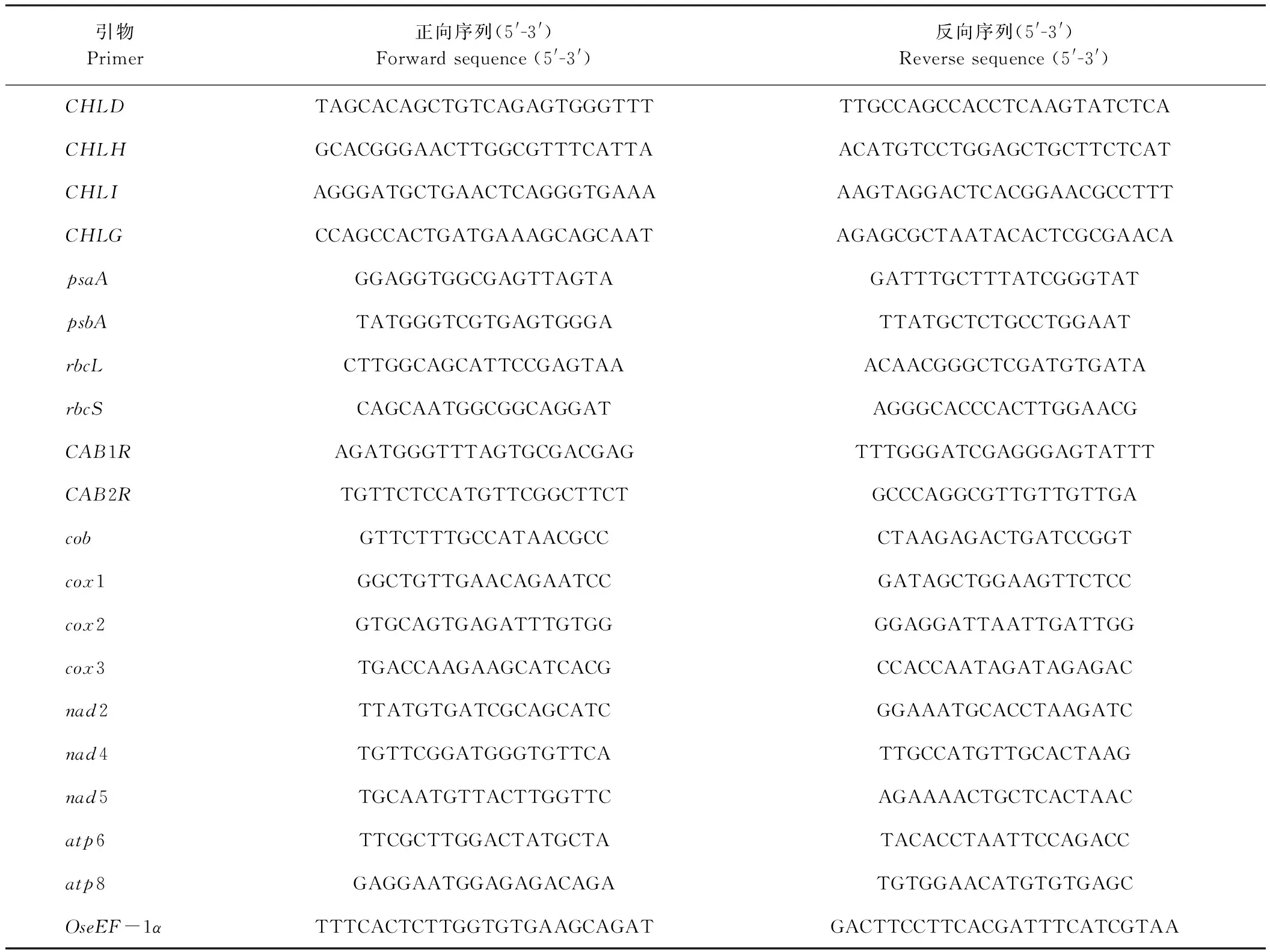
表2 光合色素代谢、光合作用以及线粒体相关基因的定量引物Table 2 Primers of photosynthetic pigment metabolism, photosynthesis mitochondrial related genes used for quantitative real-time PCR
由图6可知,在30℃条件下与野生型相比,wsl887叶片的叶绿素合成途径相关基因除CHLD显著减少,除GHLG无显著差异,其他基因的表达量均显著增加;在光合作用相关基因中,细胞质编码基因psaA、psbA和rbcL的表达量均显著减少,而细胞核编码基因CAB1R、CAB2R和rbcS的表达量均显著增加;线粒体电子传递复合体基因中,nad4的表达量无显著差异,其他基因的表达量均显著减少。说明在高温条件下,wsl887呼吸作用效率减少,光合色素合成效率增加。
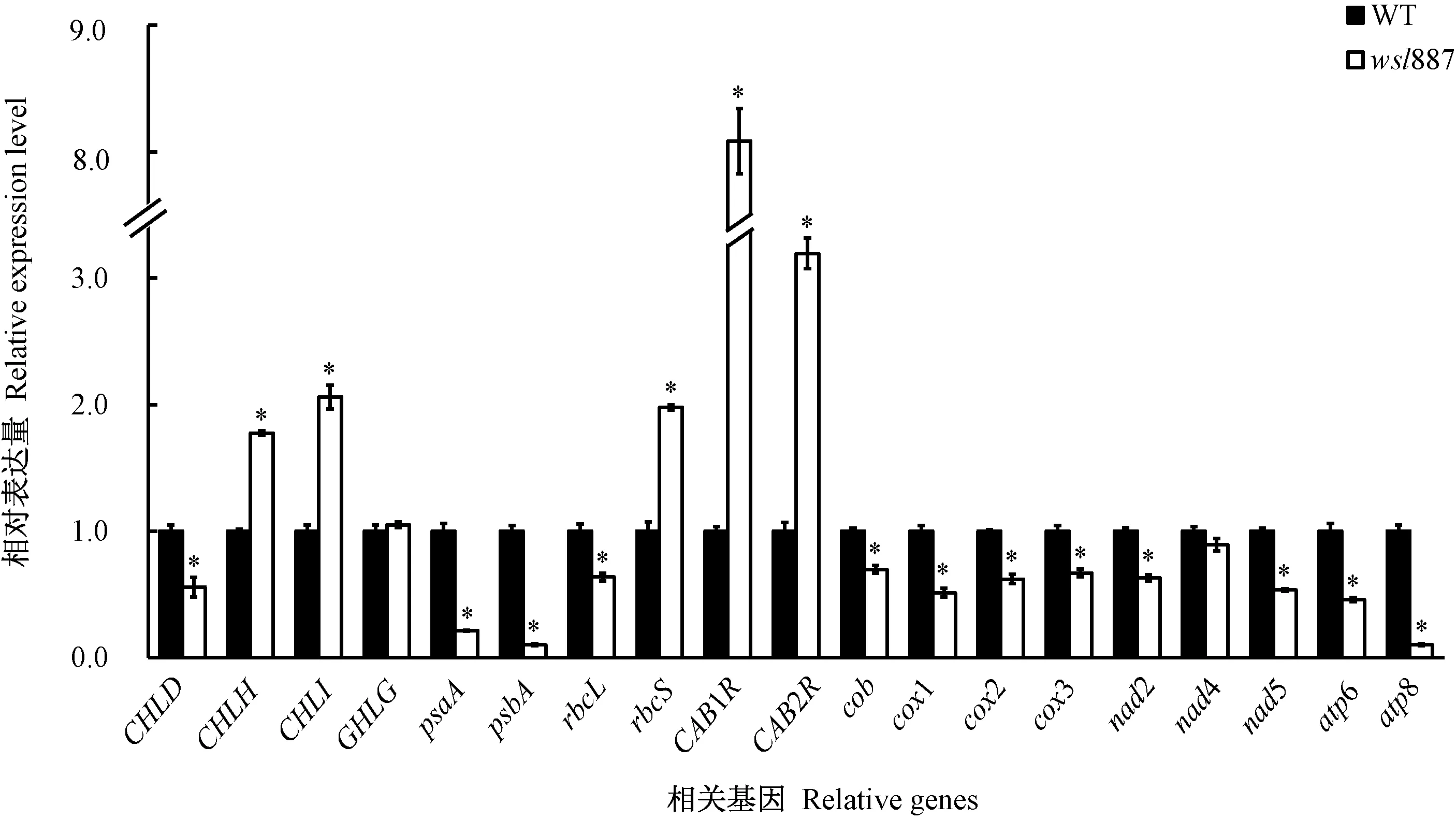
图6 30℃条件下野生型(WT)及突变体wsl887叶片中光合色素代谢、光合作用和线粒体相关基因的表达分析Fig.6 Expression analysis of genes in photosynthetic pigment metabolism, photosynthesis and mitochondrial genes of the wild type (WT) and wsl887 mutant leaves under 30℃
2.5 遗传分析与基因定位
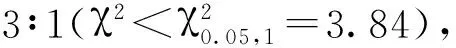
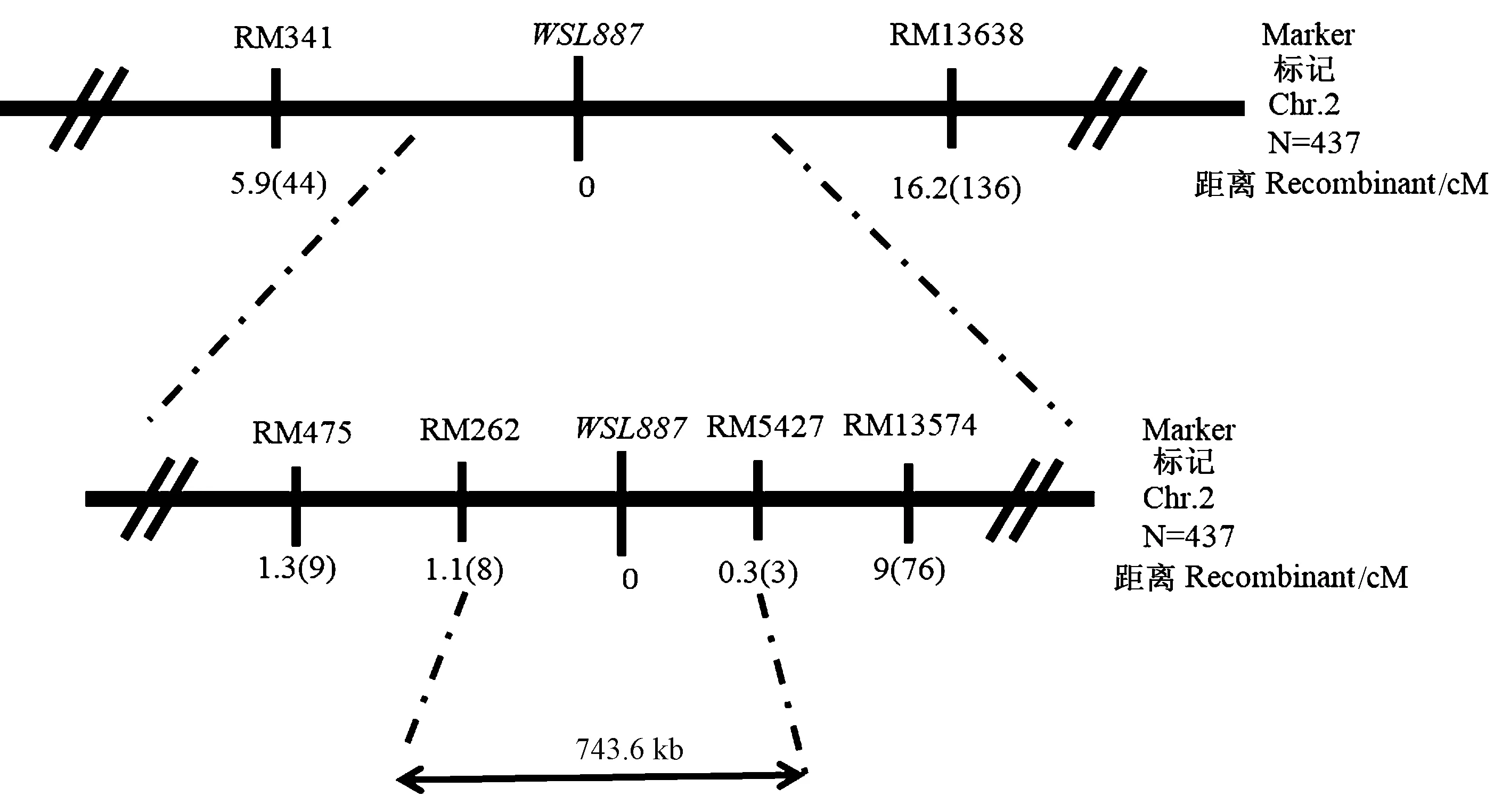
注:n: 群体大小; recombinant: 重组单株数。Note: n: Number of group. recombinant: Number of recombinant.图7 wsl887基因在2号染色体上的分子定位Fig.7 Molecular mapping of wsl887 on chromosome 2 in rice

表3 突变体wsl887的遗传分析Table 3 Genetic analysis of the wsl887 mutant
在743.6 kb区域内共存在63个候选基因(表4),经https://www.genscript.com/亚细胞定位预测,叶绿体上的基因可能有19个,其中包括一个已克隆的白条纹的基因WSL4 (LOC_Os02g35750),优先对该基因进行测序分析发现,突变体wsl887的该基因与野生型的序列完全一致,初步排除该基因的等位突变。根据 http://www.ricedata.cn/gene/和 http://www.gramene.org/提供的信息,对WSL887基因定位区间内可能定位于叶绿体上的19个基因进行注释分析(表5)。可知,LOC-Os02g34940、LOC-OsO2g35500和LOC-OS02g35750最有可能是WSL887的侯选基因。
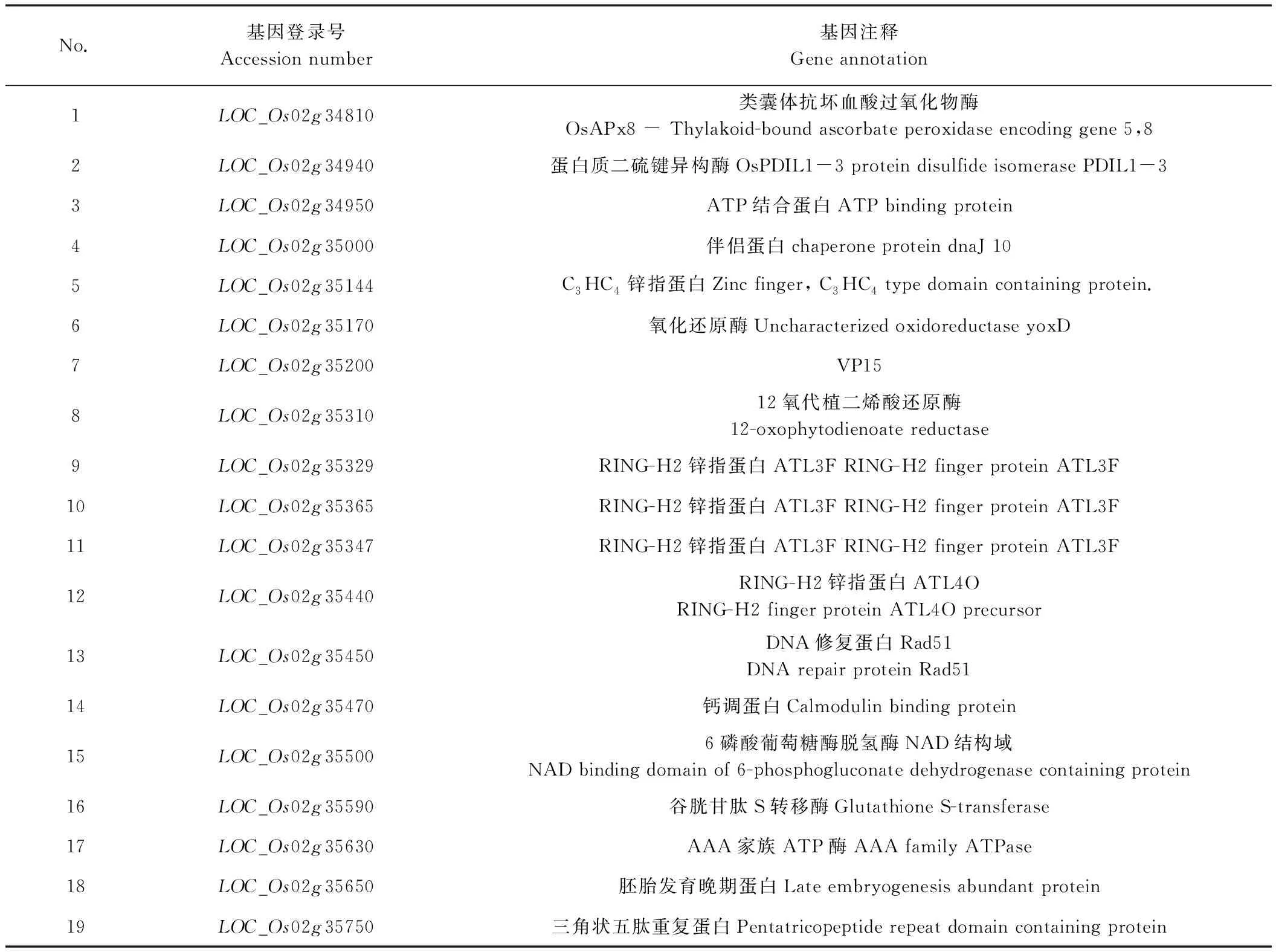
表5 wsl887基因定位区间内定位于叶绿体的注释基因Table 5 Annotated genes located in chloroplast in wsl887 gene mapping interval
3 讨论
叶绿体是植物进行光合作用的场所,对植物生长发育起重要作用。叶色突变是由于控制叶绿素生物合成和叶绿体发育相关基因的突变失活或沉默,导致叶片中叶绿素含量改变[17],因此叶色突变体是研究植物中叶绿素合成和叶绿体发育的理想材料[18-19]。近年来国内外学者对叶色突变基因的研究日益增多,其中已克隆的白化转绿基因包括V1[20]、V2[4]、V3[1]、St1[1]、Hsp70CP1[21]、TCD5[22]、WSL4[7]、OsV4[23]、DUA1[24]、TCD9[25]、TCD10[25]、OsSIG2A[26]、OsABCI8[27]、TCM12[28]、TCD11[29]、SPP[30]、VAL1[31]、TSV3[12]和VYL1[32]等,大部分叶色突变体由于叶绿素含量减少和叶绿体发育缓慢,对产量性状造成较严重的负面影响,只有少部分叶色突变体对产量性状影响较小。本研究的白条纹转绿突变体wsl887属于温敏型突变体,在20℃条件下萌发的幼苗呈白化表型,在25℃条件下萌发的幼苗呈白条纹表型,在30℃条件下突变体的表现能够恢复到野生型水平。在大田wsl887突变体苗期呈现白条纹表型,新生叶恢复正常绿色表型,在成熟期时突变体的农艺性状与野生型无显著差异,表明基因突变未影响该突变体的产量性状,在水稻生产中具有较好的应用前景。
本研究表明,wsl887受一对隐性核基因控制,将WSL887定位于2号染色体上的SSR标记RM262和RM5427之间,物理距离为743.6 kb。该区间内存在63个候选基因(表4),其中亚细胞定位于叶绿体上的基因有19个,影响叶绿体的发育。其中包括已报道的一个白条纹基因WSL4(LOC_Os02g35750)编码一种PPR蛋白[7],LOC_Os02g34940编码OsPDIL1-3蛋白二硫键异构酶,LOC_Os02g35500编码6-磷酸葡萄糖酸脱氢酶蛋白的NAD结合域,参与磷酸戊糖途径。WSL4属于P亚家族的PPR蛋白,对水稻叶绿体RNA Ⅱ内含子剪接至关重要。wsl4突变导致atpF、ndhA、rpl2和rps12的剪接缺陷。前人研究表明,在20℃时wsl4突变体呈白化表型,几乎检测不到光合色素;在30℃/25℃(白/昼)时呈白条纹表型,光合色素显著减少;在30℃条件下部分恢复表型,但光合色素含量与野生型仍然有较显著的差异[7]。本研究中突变体wsl887在20℃时,白化叶片中的叶绿素a、叶绿素b、总叶绿素及类胡萝卜素含量极显著降低;随温度升高,突变体wsl887的光合色素含量逐渐上升,在30℃条件下与野生型无差异,能够完全恢复到正常野生型的表型。本研究中,在20℃条件下wsl887叶片的叶绿素合成途径相关基因的表达量无显著差异;光合作用相关基因psaA、psbA、rbcS、rbcL、CAB1R和CAB2R的表达量显著降低;部分编码线粒体电子传递复合体基因cob、nad2、nad4和nad5、cox1和cox2的表达量显著增加。在30℃条件下与野生型相比,wsl887叶片的叶绿素合成途径相关基因除CHLD显著减少以外,其他基因的表达均显著增加;在光合作用相关基因中由细胞质编码的基因psaA、psbA和rbcL的表达量均显著减少,而由细胞核编码的基因CAB1R、CAB2R和rbcS的表达量均显著增加;线粒体电子传递复合体基因中除nad4以外,其他基因的表达量均显著减少。由细胞质编码的psaA、psbA和rbcL基因的表达量在不同温度条件下均显著减少,可以推测该基因可能与细胞质中叶绿体的发育有关。而wsl4中光合色素合成和光合作用相关基因CHLD、CHLH、CHLI、CAB1R、CAB2R、rbcS、psaA、psbA和rbcL的表达均显著下调[7],wsl887与wsl4的光合作用相关基因表达的变化趋势基本一致,均较野生型显著下调,而在光合色素合成相关基因的表达上存在较大差异。因此,推测wsl887可能是新的温敏型叶色突变体。
本研究对突变体wsl887与野生型中的WSL4基因的全长进行测序分析,未发现碱基突变,因此推测WSL887可能是一个新的温敏型叶片白条纹转绿基因。目前尚不能确定WSL887的侯选基因,有待进一步进行基因测序分析和候选基因的遗传互补验证试验。
4 结论
白条纹转绿突变体wsl887是一个新的温敏型叶色突变体,wsl887表型受温度诱导,在20℃下萌发的wsl887幼苗呈现白化,叶肉细胞中光合色素含量少,叶绿体发育缺陷。在25℃条件下,wsl887幼苗呈白条纹表型,叶片光合色素含量增加,但与野生型差异显著。在30℃条件下,wsl887幼苗呈现正常绿叶表型,叶片光合色素含量与野生型无显著差异。WSL887基因突变影响光合作用和叶绿体发育相关基因的表达,而对光合色素代谢和线粒体电子传递相关基因的表达影响较小。wsl887突变表型受一对隐性核基因控制,定位于2号染色体上的SSR标记RM262和RM5427之间,约743.6 kb 区域。本研究为下一步的WSL887基因克隆和阐释该基因对叶绿体发育的分子调控机制奠定了基础。
猜你喜欢
作物学报(2022年2期)2022-11-06
亚热带农业研究(2022年1期)2022-08-08
作物学报(2022年8期)2022-05-29
蔬菜(2021年7期)2021-11-27
语文世界(小学版)(2021年5期)2021-06-04
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
江苏农业科学(2016年2期)2016-04-11
