
鸡Ii结合Rab5a和Rab7b分子的分析
2021-12-31 01:24陈芳芳桂亚萍于凤梅李锦春刘翠艳查丽莎
畜牧兽医学报 2021年12期
陈芳芳,桂亚萍,于凤梅,张 俊,谈 阳,李锦春,刘翠艳,查丽莎
(安徽农业大学动物分子与应用免疫创新团队,合肥 230036)
恒定链(invariant chain,Ii)是一种非多态的Ⅱ型跨膜蛋白。Ii参与主要组织相容性复合体(major histocompatibility complex,MHC)Ⅱ类分子的装配、成熟和转运,在内吞体中协助MHC Ⅱ类分子进行抗原肽加工和提呈;后来发现,Ii还可以协助MHC Ⅰ类分子对抗原肽的转运,即Ii参与了抗原的交叉递呈[1-3]。Ii还是适应性免疫的多功能调节器,如负责将MHCⅠ类和Ⅱ类分子以及其他Ii相关分子(如Fc受体)分选进入特定的内吞体途径和调节B细胞的发育和成熟等[3-6]。Ii的活性片段可携带抗原肽在细胞内吞体中分别与MHCⅠ类和Ⅱ类分子共定位和结合,参与抗原递呈,这个特性使Ii活性片段作为疫苗载体可以直接进入内吞体途径,易化抗原在细胞内的转运[7]。所以,以Ii为载体的疫苗得到广泛应用[8-9]。此外,Ii还是一些细胞因子和病毒的受体[10-13]。这些功能的发现和应用均与Ii在细胞中的定位有关,尤其是在内吞体中的定位。当Ii表达时,内吞体的成熟和蛋白酶降解被延迟,在一些非抗原提呈细胞内,内吞体增大,其机制与内吞体中SNARE[soluble N-ethylmaleimide sensitive-factor(NSF)attachment protein (SNAP)receptor]蛋白的Vti1b有关[14]。
Rab是介导细胞内分子转运的重要家族,它们分布于真核细胞所有与浆膜相关的细胞器(如细胞核、高尔基体和内吞体等),在细胞器间转运活性蛋白过程中起重要作用,其中,Rab5a是介导细胞吞噬过程的必要调节蛋白,对抗原及抗原递呈分子的转运具有重要作用。研究发现其影响MHC Ⅱ类分子对抗原肽的递呈,应用辛伐他汀可以抑制这种作用[15-17]。Rab7b在哺乳动物中主要定位于晚期内吞体、溶酶体和高尔基体。其不仅参与胞内活性蛋白从早期内吞体到晚期内吞体/溶酶体的转运,而且还是内吞体到高尔基体或/和跨高尔基体系统(trans-Golgi network,TGN)的重要调节蛋白[18-19]。
课题组此前发现鸡Ii可以分别与Rab5a和Rab7b结合,并在细胞内与之共定位[20-21]。但是并不清楚在Ii的多个结构域中与Rab分子结合以及在细胞定位中的作用。为此,本研究构建了Ii的不同结构域片段以及Ii突变体,探明它们与Rab5a和Rab7b结合和共定位的特性,以进一步解析Ii在免疫反应中细胞内转运机制。
1 材料与方法
1.1 主要试验材料
PCR plus mix、DNA Marker DL2000、质粒提取试剂盒、限制性内切酶和蛋白Marker购自东盛生物(广州)有限公司;T4连接酶购自东洋纺(日本)有限公司;转染试剂lipo8000购自碧云天(上海)有限公司;广谱彩虹预染蛋白Marker购自康为世纪(北京)有限公司;胎牛血清(fetal bovine serum,FBS)和DMEM培养基购自Hyclone(美国)有限公司;镍柱、Gluttathione Sepharose 4B柱和电化学发光(electrogenerated chemiluminescence,ECL)试剂盒购自GE(瑞士)有限公司;小鼠抗GFP、RFP、His抗体和酶标记山羊抗鼠抗体购自中杉金桥(北京)有限公司;重组质粒pmCherry-C1、pEGFP-C1、pEGFP-C1-cIi、pGEX-4T-1、pET-32a-cIi、pGEX-4T-1-cRab5a、pGEX-4T-1-cRab7b、pmCherry-C1-cRab5a、pmCherry-C1-cRab7b、人胚胎肾细胞系293 T细胞和大肠杆菌(E.coli)Rostta(DE3)菌株由本实验室保存。
1.2 Ii功能域和Ii突变体基因的扩增
用自行设计的引物,以实验室保存的含有cIi质粒为模板,分别克隆含Ii胞浆区和跨膜区结构域的cIi(Cyt-Tra)和含构成内质网腔区的CLIP和三聚体区的cIi(CLIP-TRIM);同时,应用重叠延伸PCR法获得3个Ii突变体cIi(M81-87aa)、cIi(M91-99aa)和cIi(M81-99aa),突变范围在CLIP片段内,分别为突变CLIP的81-87位(M81-87aa)、91-99位(M91-99aa)和81-99位(M81-99aa)氨基酸。PCR和重叠延伸方法参照陈芳芳等[21]。
1.3 重组质粒的构建和鉴定
利用双酶切方法分别将上述克隆的5个基因片段定向插入真核和原核表达载体pEGFP-C1和pET-32a中,构建相应的重组质粒。然后转化至感受态细胞Rosetta(DE3)中,分别构建含6个真核表达质粒和6个原核质粒的重组菌Rosetta(DE3)pEGFP-C1-cIi(M81-87aa)、Rosetta(DE3)pEGFP-C1-cIi(M91-99aa)、Rosetta(DE3)pEGFP-C1-cIi(M81-99aa)、Rosetta(DE3)pEGFP-C1-cIi(Cyt-Tra)、Rosetta(DE3)pEGFP-C1-cIi(CLIP-TRIM)、Rosetta(DE3)pEGFP-C1、Rosetta(DE3)pET-32a-cIi(M81-87aa)、Rosetta(DE3)pET-32a-cIi(M91-99aa)、Rosetta(DE3)pET-32a-cIi(M81-99aa)、Rosetta(DE3)pET-32a-cIi(Cyt-Tra)和Rosetta(DE3)pET-32a-cIi(CLIP-TRIM)以及对照的空载体Rosetta(DE3)pET-32a。构建的重组菌经PCR和酶切初步鉴定后送擎科(南京)生物公司测序。质粒构建和鉴定的方法同陈芳芳等[21]。本研究中所用引物、基因序列和构建的重组质粒名称见表1。
1.4 真核转染和激光共聚焦观察
将构建的含有绿色荧光的重组质粒与本实验室保存的带有红色荧光的重组质粒pmCherry-C1-cRab5a和pmCherry-C1-cRab7b分别共转染293 T细胞。继续培养24 h后用PBS清洗。4%多聚甲醛固定10 min,PBS洗3次后封片,在LSM 880 with Airyscan激光共聚焦显微镜观察真核重组质粒共转染后在细胞内的共定位。同时为了验证转染的重组质粒在细胞内蛋白的表达,应用Western blot进行蛋白的特异性鉴定。细胞转染、观察和鉴定的方法同陈芳芳等[21]。
1.5 原核重组菌的蛋白表达与鉴定
将上述经鉴定正确的原核重组表达菌和本实验室保存的重组菌Rosetta(DE3)pGEX-4T-1-cRab7b、Rosetta(DE3)pGEX-4T-1-cRab5a和Rosetta(DE3)pGEX-4T-1进行蛋白表达、纯化和鉴定。重组菌用异丙基-β-D-硫代半乳糖苷(isopropyl β-D-thiogalactoside,IPTG)0.5 mmol·L-1,16 ℃进行诱导表达24 h。离心收集的菌体用180 W,5 s,间隔10 s超声波破碎菌体,12 000g离心15 min,上清中的蛋白用十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)进行电泳和染色观察。His和GST标签的蛋白分别用镍柱和GST亲和层析柱(glutathione sepharose 4B)进行纯化,纯化后蛋白用SDS-PAGE电泳进行鉴定。
1.6 拉下法(Pull-down)检测目的蛋白间的结合
将带有GST标签蛋白的GST/cRab5a、GST/cRab7b、GST蛋白分别加入GST亲和层析柱内作为锚定蛋白,放入4 ℃冰箱内平衡30 min左右,将携带His标签的蛋白His/cIi、His/cIi(M81-87aa)、His/cIi(M91-99aa)、His/cIi(M81-99aa)、His/cIi(Cyt-Tra)、His/cIi(CLIP-TRIM)以及His分别加入柱内,孵育1 h后用10 mmol·L-1谷胱甘肽洗脱液洗脱,收集蛋白洗脱液,分别进行SDS-PAGE凝胶电泳,根据电泳条带和免疫印迹法的特异性鉴定结果判定蛋白间有无相互作用关系。具体操作方法同陈芳芳等[21]。
2 结 果
2.1 Ii功能域和突变体基因的克隆及其鉴定
首先,应用PCR和重叠延伸扩增技术获得了5个目的基因片段,在琼脂糖核酸凝胶电泳检测中,cIi(M81-87aa)、cIi(M91-99aa)和cIi(M81-99aa)大小均为672 bp(图1泳道1、3、5),cIi(Cyt-Tra)和cIi(CLIP-TRIM)大小分别为258和450 bp(图1泳道7、9),这些扩增产物大小与预期一致。其次,将这些片段插入相应载体,构建成重组质粒,经双酶切鉴定,其产物在相应泳道中均出现预期大小的片段(图1泳道2、4、6、8和10)。


M.DNA相对分子质量标准;1、3、5、7、9.PCR产物;2、4、6、8、10.质粒Xho I/EcoR I 双酶切产物M.DNA marker;1,3,5,7,9.PCR amplified proudct;2,4,6,8,10.Recombinant plasmids digested with Xho I and EcoR I图1 基因的扩增和重组质粒双酶切鉴定Fig.1 Amplification of genes and identification of recombinant plasmids by restriction endonuclease digestion
2.2 真核表达的Ii结构域、Ii突变体与Rab5a和Rab7b的细胞内共定位
为确定所构建的重组质粒在细胞内的定位特性,将含绿色荧光的重组质粒pEGFP-C1-cIi、pEGFP-C1-cIi(M81-87aa)、pEGFP-C1-cIi(M91-99aa)、pEGFP-C1-cIi(M81-99aa)、pEGFP-C1-cIi(Cyt-Tra)、pEGFP-C1-cIi(CLIP-TRIM)、pEGFP-C1与含红色荧光蛋白的重组质粒pmCherry-C1-cRab5a(或者pmCherry-C1-cRab7b)共转染293 T细胞。应用免疫印迹法进行蛋白质特异性的鉴定。结果显示,细胞内表达的重组绿色荧光蛋白/红色荧光蛋白分别被抗GFP/RFP抗体所识别,出现与预期大小相符合的条带(图2A、B),这说明所构建的重组蛋白在细胞内得到表达。在激光共聚焦显微镜下检测相关目的蛋白的共定位关系(图3)。对照组GFP/cIi与RFP/cRab5a(或RFP/cRab7b)定位于细胞内同一部位而在叠加图中呈现黄色(图3A、B中a);同样,GFP/cIi(M81-87aa)、GFP/cIi(M91-99aa)、GFP/cIi(M81-99aa)、GFP/cIi(Cyt-Tra)也均与RFP/cRab5a(或RFP/cRab7b)在细胞内共定位而呈现黄色(图3A和3B b、c、d和e);GFP/cIi(CLIP-TRIM)均匀分布于整个细胞(图3A、B f),而空载体GFP则更多定位于细胞核,两者在叠加图中均不显示出与RFP/cRab5a(或RFP/cRab7b)共定位的黄色(图3A、B g)。以上结果表明,Ii、cIi(Cyt-Tra)及Ii突变体均可以与cRab5a或cRab7b在细胞内共定位,而CLIP与三聚体[cIi(CLIP-TRIM)]却不能。

图2 Ii功能域、Ii突变体、Rab5a和Rab7b分子真核表达蛋白的鉴定Fig.2 Identification of Ii domain,its mutants,Rab5a and Rab7b in eukaryotic expression
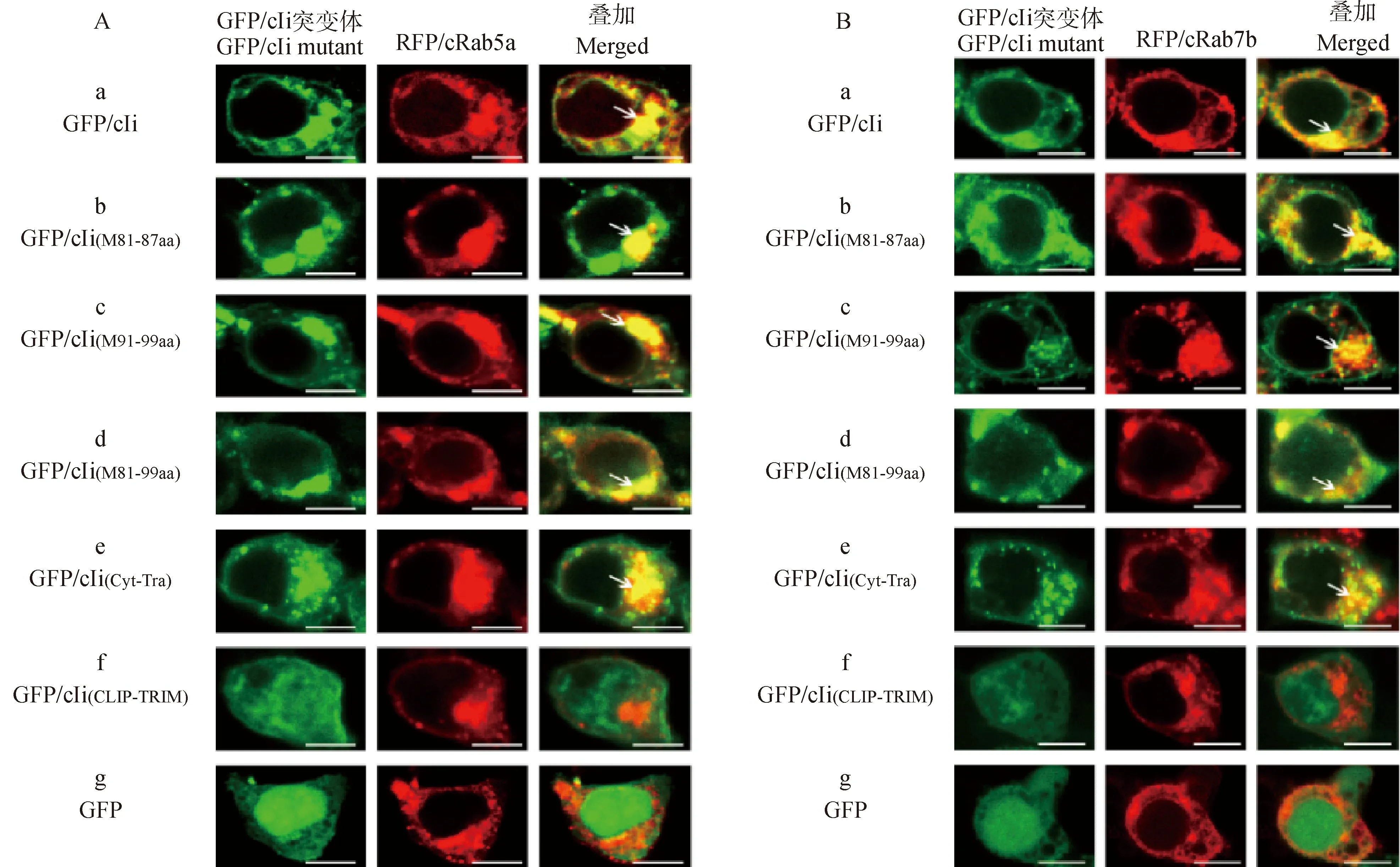
GFP.绿色荧光蛋白;RFP.红色荧光蛋白;标尺=50 μm;箭头所指为关联分子的共定位GFP.Green fluorescent protein;RFP.Red fluorescent protein;Bar=50 μm;the arrows show the co-localization of associated molecules图3 cIi突变体和功能域与Rab5a和Rab7b在真核细胞的共定位Fig.3 Colocalization of mutant and functional domain cIi with Rab5a and Rab7b in eukaryotic cells
2.3 原核表达的Ii结构域、Ii突变体重组质粒的蛋白表达与纯化
构建的重组菌Rosetta(DE3)pET-32a-cIi(M81-87aa)、Rosetta(DE3)pET-32a-cIi(M91-99aa)、Rosetta(DE3)pET-32a-cIi(M81-99aa)和空载体重组菌Rosetta(DE3)pET-32a进行蛋白的表达和纯化,如图所示,它们表达的蛋白His/cIi(M81-87aa)、His/cIi(M91-99aa)、His/cIi(M81-99aa)和His/cIi分子量相近分别为46.2 ku,而His蛋白大小为22 ku(图4A泳道 2、3、4、1和5)。而Rosetta(DE3)pET-32a-cIi(CLIP-TRIM)和Rosetta(DE3)pET-32a-cIi(Cyt-Tra)表达的蛋白大小分别为38.5和31.1 ku(图4B泳道 1和2),所表达的蛋白大小与预期相符合,这表明所构建的重组质粒可以正确表达目的蛋白。
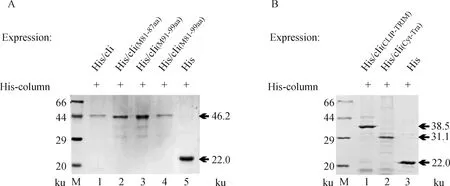
图4 cIi突变体与功能域原核表达蛋白的纯化Fig.4 Purification of prokaryotic expression protein of Ii mutant and functional domain
2.4 原核表达的Ii结构域、Ii突变体结合Rab5a和Rab7b特征的确定
为了解析Ii突变体[His/cIi(M81-87aa)、His/cIi(M91-99aa)和His/cIi(M81-99aa)]与cRab5a/7b的相互作用关系,应用Pull-down方法进行检测。GST/cRab5a、GST/cRab7b和GST分别作为锚定蛋白,GST或者带GST标签的蛋白可以与填料结合而留在柱内,当后加入的His标签蛋白的样品中存在与其结合的部分则随之被留在柱内,最后被解离液洗脱下来。如果没有与其结合的蛋白则最后解离液中只有单一的GST/cRab5a、GST/cRab7b或GST蛋白存在。
每组均以经证实的His/cIi与GST/cRab5a(或者GST/cRab7b)作为阳性对照,先检测His/cIi(M81-87aa)的结合特性。结果发现,当GST/cRab5a(或者GST/cRab7b)先结合在亲和层析柱时,后加入柱子中的His/cIi和His/cIi(M81-87aa)可以与GST/cRab5a(或者GST/cRab7b)结合而在解离液中出现两个条带,分别是His/cIi与GST/cRab5a(或者GST/cRab7b)和His/cIi(M81-87aa)与GST/cRab5a(或者GST/cRab7b)(图5A、B的泳道2、3),且解离条带与相应蛋白条带大小一致(图5A、B的泳道5、6),His空载体不与其相结合而出现单一条带(图5A、B的泳道4)。对照组的GST与His/cIi、His/cIi(M81-87aa)共同孵育时,仅出现GST标签蛋白的单一条带(图5A和B的泳道8、9)。进一步应用抗His的抗体对相应蛋白进行检测,结果发现,在相应泳道出现特异性条带(图5A、B的泳道2、3、5、6和7)。用相同的方法和步骤检测了His/cIi(M91-99aa)和His/cIi(M81-99aa)分别与GST/cRab5a(或者GST/cRab7b)的结合。结果发现,它们的解离液中均出现2条带,分别是GST/cRab5a(或者GST/cRab7b)和His/cIi(M91-99aa)(或者His/cIi(M81-99aa))(图6A、B的泳道2、3),且解离条带与相应蛋白条带大小一致(图6A、B的泳道4、5),而对照组均为标签蛋白GST的单一蛋白条带(图6A、B的泳道6、7)。

图5 His/cIi(M81-87aa)和GST/cRab5a与GST/cRab7b结合的鉴定Fig.5 Identification of binding of His/cIi(M81-87aa)with GST/cRab5a and GST/cRab7b
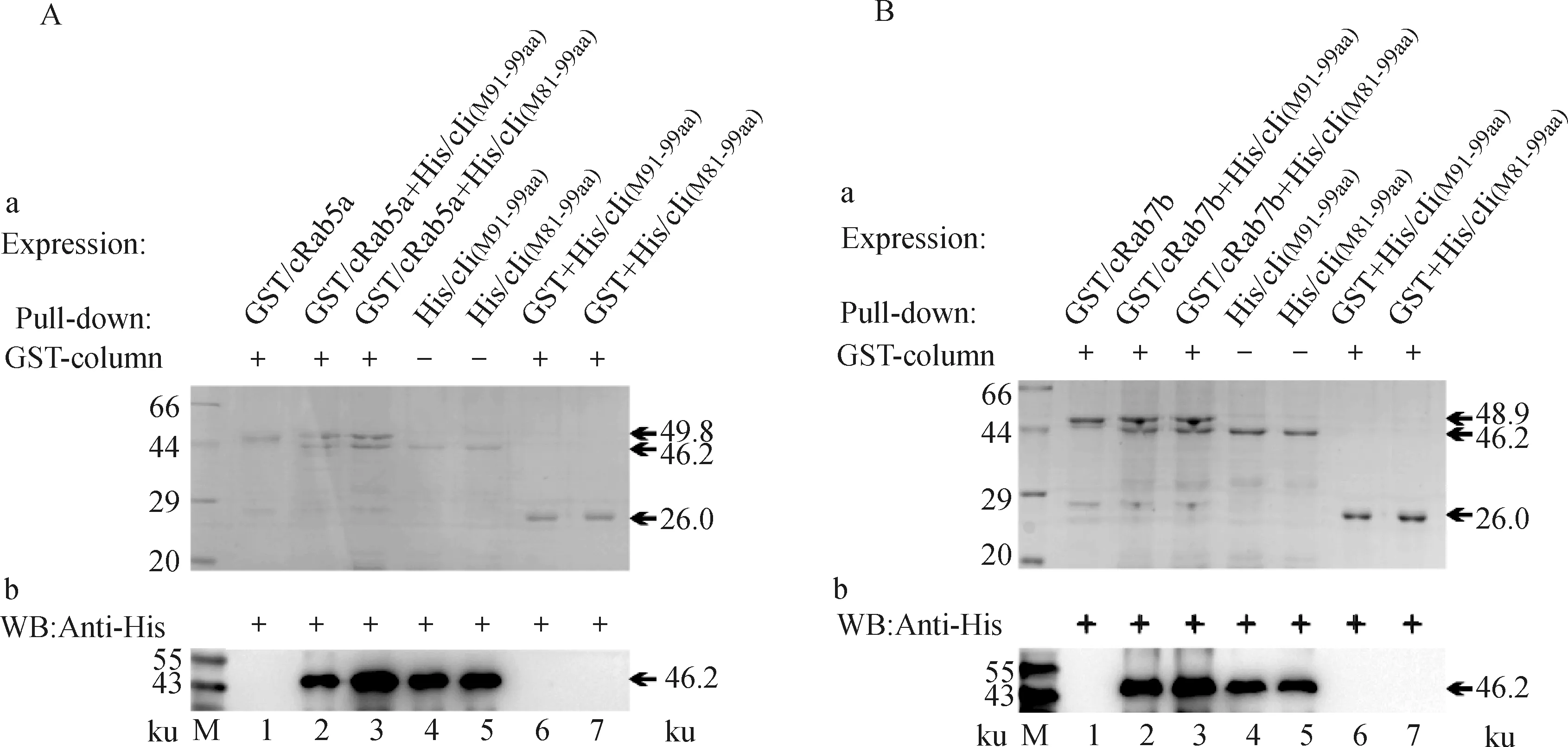
图6 His/cIi(M91-99aa)和His/cIi(M81-99aa)与GST/cRab5a和GST/cRab7b蛋白质相互作用的鉴定Fig.6 Identification of binding of His/cIi(M91-99aa)and His/cIi(M81-99aa)with GST/cRab5a and GST/cRab7b proteins
以上结果表明,Ii突变体均可以与GST/cRab5a(或者GST/cRab7b)结合,且这种结合不受标签蛋白的影响。这说明,Ii突变体也可以与Rab5a和Rab7b结合,Ii的CLIP区域不是其结合的主要结构域。
2.5 cIi结合Rab5a和Rab7b功能域的确定
进一步应用相同方法验证His/cIi(Cyt-Tra)和His/cIi(CLIP-TRIM)与GST/cRab5a(或者GST/cRab7b)结合。SDS-PAGE结果显示,在解离液中出现两个条带,分别是His/cIi(Cyt-Tra)和GST/cRab5a(或者GST/cRab7b)(图7A、B泳道2),与对应的蛋白条带大小一致(图7A、B泳道3),而His/cIi(CLIP-TRIM)不能与之结合,在电泳图中仅有一条条带(图7A、B泳道4),对照组中的GST亦不能与His/cIi(Cyt-Tra)或His/cIi(CLIP-TRIM)结合(图7A、B泳道6、7)。这说明Ii与Rab5a(或者Rab7b)结合的功能域是胞浆区和跨膜区,不是CLIP和三聚体区。
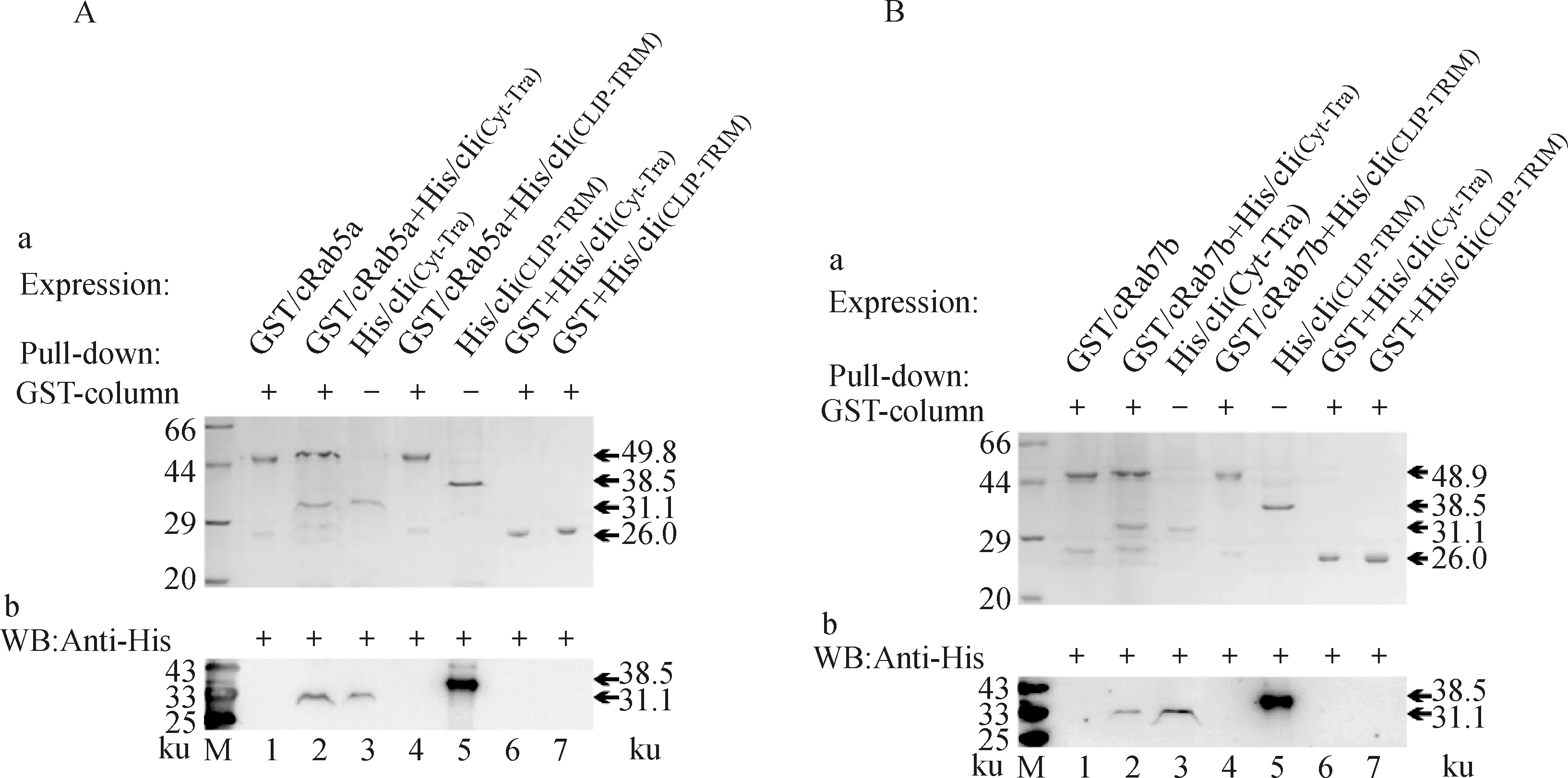
图7 His/cIi(Cyt-Tra)和His/cIi(CLIP-TRIM)与GST/cRab5a和GST/cRab7b结合的鉴定Fig.7 Identification of binding of His/cIi(Cyt-tra)and His/cIi(CLIP-TRIM)with GST/cRab5a and GST/cRab7b
3 讨 论
本研究应用拉下法检测了关联蛋白Ii结合Rab5a或Rab7b的分子特征。该法曾在课题组检测Ii与MHC分子结合中得到验证[7];还应用激光共聚焦显微镜观察了Ii与Rab分子在细胞内共定位的特征。课题组曾在前期工作中应用细胞器定位蛋白检测Rab5a或Rab7b在细胞内分布,发现它们均定位于细胞内吞体[20-21]。本研究结果(图3)表明,Ii(包括Ii胞浆区和跨膜区片段)能与Rab分子在内吞体共定位。总之,本研究获得了以下重要结果。
首先,Ii的胞浆区和跨膜区不仅能结合Rab分子,还是细胞内定位的功能结构域。鸡Ii由3个结构域组成,分别是胞浆区、跨膜区和内质网腔区。其中,内质网腔区由CLIP(81—107 aa)和三聚体区组成。项目组前期研究发现,Ii的跨膜区和胞浆区具有引导抗原肽进入内吞体的功能[7]。在本研究中,这两个结构域分别具有结合Rab5a和Rab7b的功能(图7);更重要的是,由于Ii只有在细胞内迁移中,才能发挥其从装配成熟MHC Ⅱ类分子到协助提呈外源性抗原肽不同功能,而Rab分子是胞内转运蛋白,所以,这两类分子的结合和在细胞内的共定位,不仅表明Rab分子参与Ii的胞内迁移,也提示Ii分解成胞浆区和跨膜区在迁移过程中可能存在其他重要功能,如研究发现,其可迁移到细胞核进而诱导细胞核转录因子的释放,并影响B细胞受体信号的转导[22]。此外,由于课题组在用细胞器定位的研究中已表明,Rab5a和Rab7b均定位于内吞体[20-21],所以,Ii与Ii的胞浆区和跨膜区片段分别与这两种分子共定位(图2),表明它们均可进入内吞体。
其次,本研究结果表明,Ii的CLIP尽管与结合Rab分子无关,但是对Ii分子在细胞定位是不可缺失的,它是保持空间结构的关键片段。Ii和MHC Ⅱ类分子在内质网中形成九聚体,期间Ii的CLIP占据MHC Ⅱ类分子凹槽,在迁移至内吞体后,Ii 被分解,抗原肽取代CLIP,与MHC Ⅱ类分子形成MHC-抗原肽复合物,最后被提呈给T细胞产生免疫应答。在此过程中,CLIP是重要结构域[23]。课题组前期研究发现,缺失部分CLIP片段的Ii[cIi(D81-87aa)和cIi(D91-99aa)],尽管还能维持结合Rab5a的特性,却改变了其在细胞内的定位,即缺失后的Ii分布于整个细胞,而不是细胞质[20],但是不清楚这种细胞定位的改变是否由于CLIP片段缺失导致的空间结构变化。由于CLIP分子量较小,难以在体外形成空间结构,故本研究通过突变CLIP的关键位点[cIi(M81-87aa)、cIi(M91-99aa)和cIi(M91-99aa)],探明上述问题。结果表明,突变CLIP后的Ii仍定位于细胞质,这提示Ii的CLIP对维持其在细胞内定位至关重要,由于Ii胞浆区和跨膜区具有结合Rab5a或Rab7b功能,故3个Ii突变体均维持与Rab分子结合。
4 结 论
本研究结果表明,Ii的胞浆区和跨膜区不仅能结合Rab5a或Rab7b分子,还是其在细胞内定位的功能结构域。Ii的CLIP与结合Rab5a或Rab7b分子无关,却是保持Ii分子空间结构和维持细胞内定位特性的关键片段。
猜你喜欢
数学大王·低年级(2020年8期)2020-08-14
山西地震(2019年1期)2019-03-20
生物工程学报(2019年1期)2019-01-30
上海农业学报(2017年3期)2017-04-10
安徽医科大学学报(2016年12期)2017-01-15
新农业(2016年23期)2016-08-16
小雪花·初中高分作文(2016年9期)2016-05-14
系统工程与电子技术(2016年2期)2016-04-16
天津医科大学学报(2015年2期)2015-12-22
中国光学(2015年1期)2015-06-06
