
红心火龙果不同发育时期果肉呈色相关基因表达模式研究
2021-12-23 16:55赵国文贾瑞宗郭静远郭安平
热带作物学报 2021年11期
关键词:火龙果
赵国文 贾瑞宗 郭静远 郭安平
摘 要:火龍果属于仙人掌科量天尺属,是具有重要经济和营养价值的热带水果之一,其果实富含花青素和甜菜红素。在果实成熟后期甜菜红素的大量积累引起红心火龙果果实转色的现象。果肉色泽是评价火龙果果实品质的重要指标之一,然而其果实转色背后的基因表达调控模式并不清晰。深入研究火龙果果实发育过程中果肉颜色调控的分子机理有助于丰富火龙果品质形成的理论基础。本研究以‘美红一号’红心火龙果为实验材料,采集果实发育过程中9个不同时期的样品,通过比较转录组学共识别了96种差异表达的基因,最后分析发现15个与色素代谢通路相关的基因在果实发育不同阶段显著差异表达。GO功能富集分析发现差异表达基因主要富集在与结合活性、催化活性、转运活性和结构分子活性相关的功能分类上,且与色素代谢都有重要联系;KEGG代谢途径富集结果显示,次生代谢物合成途径、氨基酸和核酸代谢途径、酪氨酸代谢途径富集基因较多。利用实时荧光定量PCR技术对15个与色素相关的差异表达基因进行了验证,其结果与转录组分析结果一致。生理分析和转录组分析结果表明,红皮红肉火龙果发育过程中伴随大量甜菜色素的合成,且甜菜色素合成途径中DODA基因逐渐上调,在果实转色其中在P4时期明显的上升趋势,其它关键基因表达逐渐下调。本研究发掘火龙果中与果实转色相关的相关基因,为后续火龙果遗传改良提供了重要的目标基因和遗传基础。
关键词:火龙果;果实转色;转录组测序;基因挖掘;
中图分类号:S667.9 文献标识码:A
Abstract: Pitaya belongs to the genus of Cactaceae and is one of the tropical fruits with important economic and nutri-tional value. Its fruits are rich in anthocyanins and betaine. In the late stage of fruit maturity, the accumulation of beta red pigment causes the phenomenon of red-hearted dragon fruit fruit color change. The color of the pulp is one of the important indicators to evaluate the quality of the dragon fruit. However, the gene expression regulation mode behind the color change of the fruit is unclear. In-depth study of the molecular mechanism of the color regulation of the pitaya fruit during the development of the dragon fruit will help to enrich the theoretical basis for the formation of pitaya quality. In this study, ‘Meihong No. 1’ red-hearted dragon fruit was used as the experimental material. Nine samples from different stages of fruit development were collected. A total of 96 differentially expressed genes were identified through comparative transcriptomics. Finally, 15 genes were found to be related to pigments. Genes related to metabolic pathways were significantly differentially expressed at different stages of fruit development. GO functional enrichment analysis found that differentially expressed genes were mainly enriched in functional classifications related to binding activity, catalytic activity, transport activity and structural molecule activity, and were importantly related to pigment metabolism. The results of KEGG metabolic pathway enrichment showed that there were many enriched genes in the biometabolite synthesis pathway, amino acid and nucleic acid metabolism pathway, and tyrosine metabolism pathway. Real-time fluorescent quantitative PCR technology was used to verify 15 the differentially expressed genes related to pigments, and the results were consistent with the results of transcriptome analysis. The results of physiological analysis and transcriptome analysis showed that the development of red skin and red flesh dragon fruit was accompanied by the synthesis of a large number of beet pigments, and the DODA gene in the beet pigment synthesis pathway was gradually up-regulated. During the fruit color change, there was an obvious upward trend in the P4 period. The expression of key genes is gradually down-regulated. This study explored the related genes related to fruit color change in dragon fruit, and provided an important target gene and genetic basis for subsequent genetic improvement of dragon fruit.
Keywords: dragon fruit; fruit color conversion; transcriptome sequencing; gene mining
DOI: 10.3969/j.issn.1000-2561.2021.11.0012
火龙果起源中美洲热带雨林地区,为仙人掌科量天尺属,果实外形独特,香甜多汁,含有花青素和蛋白质等多种成分,营养价值较高深受消费者的喜爱[1-3]。火龙果原产地在中美洲的墨西哥、古巴等地区[4],在我国火龙果的种植主要在南方的广西、福建、云南、海南等地进行种植[5],为当地果农带来了良好的经济收益。红皮红肉(Hylocereus costaricensis)品种是一种新的改良品种[6],其果实呈近圆形,鳞片粗短,反卷似“莲花座”,是火龙果的主要栽培品种[7]。目前,火龙果研究不仅在种植栽培、生物学特性、储藏保鲜等方面进行;同时也在火龙果分子生物学、细胞生物学、色素提取[8-10]和植物病害[11-12]等方面取得了一定的进展。但火龙果基因组序列尚未报道,其分子生物学的研究仍受到较大限制。
火龙果根据其果皮果肉颜色分为红皮白肉、红皮红肉和黄皮白肉3种[13]。市场上常见的品种是红皮红肉和红皮白肉,红皮红肉更受到消费者喜爱。目前,火龙果色素生物合成途径已基本明确,代谢途径中的关键酶也已克隆分离,但对色素合成基因表达模式调控的研究还十分有限。因此,本研究以‘美红一号’红皮红肉火龙果为实验材料,进行转录组测序和后续分析,比较研究同一品种火龙果果肉在不同时期中基因的差异表达情况,对其进行相关的基因功能和代谢通路富集分析,筛选出差异表达基因,进一步对红肉火龙果中色素的代谢差异进行初步分析。通过对筛选出的基因开展功能验证实验,可以为甜菜色素合成代谢提供可靠信息。
1 材料与方法
1.1 材料
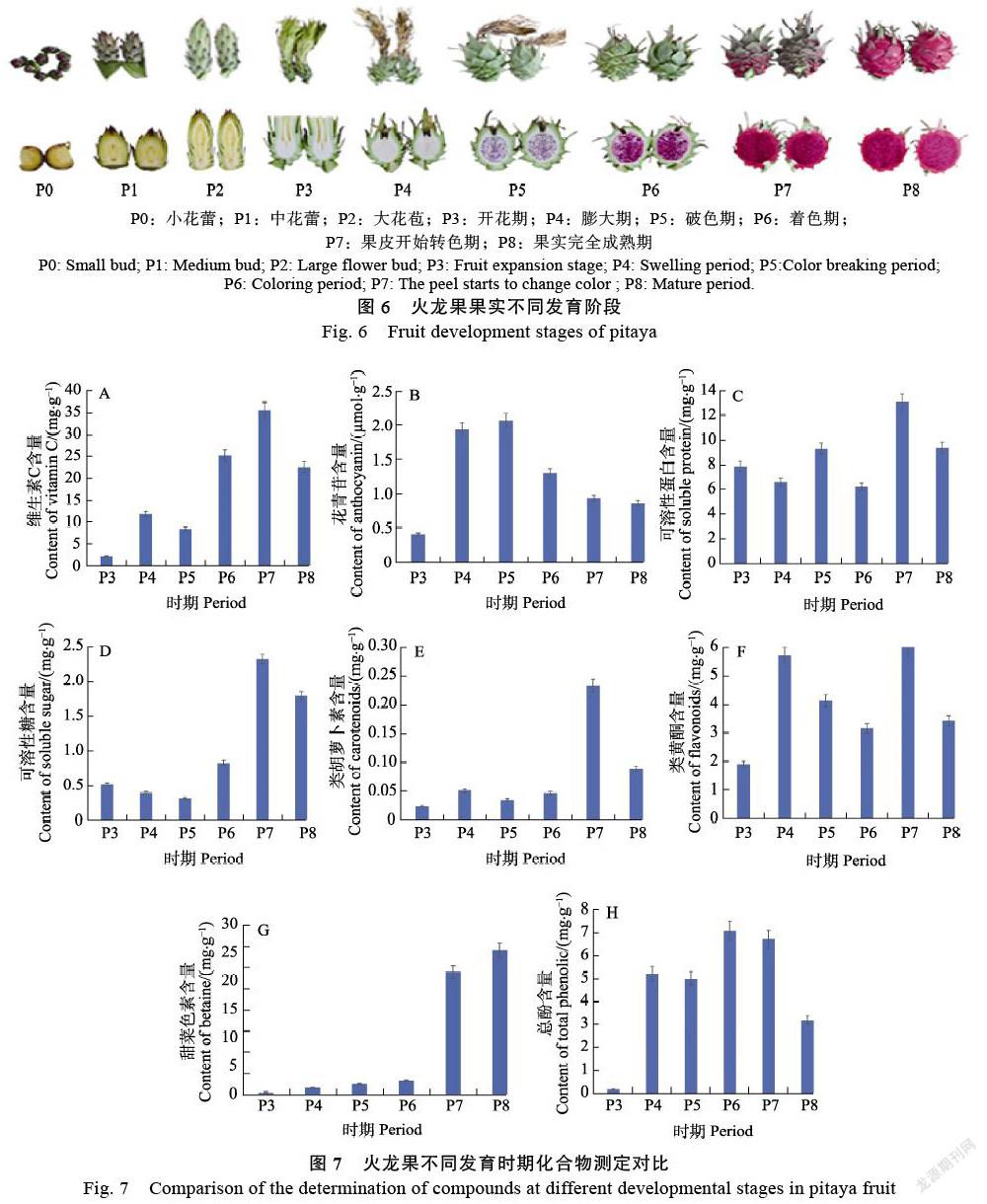
‘美红一号’红心火龙果(Hylocereus cos-taricensis)的P0:小花蕾;P1:中花蕾;P2:大花苞;P3:开花期;P4:膨大期;P5:破色期;P6:着色期;P7:果皮开始转色期;P8:果实完全成熟期9个发育时期的果肉样品的和根、茎、花、果皮和果肉5个不同组织的样品。实验材料来自海南省昌江市火龙果种植基地,现场取样并用液氮进行速冻处理,每个个体设计3个生物学重复,总共42份样品,存于–80 ℃冰箱。
1.2 方法
1.2.1 RNA提取与检测 将火龙果果实9个不同发育时期果肉组织和不同组织部位样品寄送至生工生物工程(上海)股份有限公司进行基于IlluminaHiSeqTM2000的转录组测序。样品放入含有液氮的研缽成粉末,转入放有裂解液的EP管中,采用CTAB法提取RNA[14]。取适量样品进行检测,于–80 ℃保存。RNA提取流程与检测均委托生工生物工程(上海)股份有限公司。
1.2.2 文库构建和测序 使用SMRT Link的Iso-Seq protocol进行全长转录本分析,将最终得到的高质量全长转录本用于后续分析。火龙果果肉样品在PacBio Sequel平台上测序,建立PacBio IsO-Seq文库,获得原始聚合酶read序列,去除read序列,使用SMRT分析套件进行reads of insert(ROI)、分类、聚类和校正,最终得到高质量的全长共有序列。将每个库的高质量全长序列组合在一起,在没有参考基因组的情况下,使用LoRDEC软件利用Illumina二代数据对一致的转录本序列进行纠错,去除聚集和纠错的转录本。在PacBio RS sequel测序平台是基于单分子实时(SMRT)的测序技术,环状测序得到polymerase read。polymerase read中含有很多无效数据会对后续分析带来严重干扰,如测序接头序列,建库长度的偏差,以及测序错误、低质量碱基、未测出的碱基等情况,因此需通过一些手段将上述无效数据过滤掉,以保证分析的正常进行。最终得到有效的subreads插入片段。
1.2.3 内参基因的选择 内参基因在各种组织和细胞中的表达量相对恒定,常作为检测基因表达水平变化的参考。其作用是校正上样量和上样过程中的实验误差,保证实验结果的准确性。通过检测每个样品的内参量,可用于校正上样误差,使定量结果更加可靠。一般应选择在加工因子条件下表达不发生变化的基因作为内参。本研究选择TBP2作为内参基因(表1)。
1.2.4 转录组数据的处理 测序后,对得到的所有原始数据(raw data)进行分析,过滤后得到clean data,然后去除无效的低质量序列。进行PacBio测序数据评估,最终得到有效的subreads inserts。使用SMRT Link的Iso-Seq protocol进行全长转录本分析,并使用最终的高质量全长转录本进行后续分析。基于Swissprot、Nr、KOG、GO和KEGG数据库,对所有转录本进行注释,包括表达注释和功能注释,最终富集分析。
1.2.5 差异分析 用TPM方法计算转录本的表达水平。对于没有生物学重复的样本,先用TMM对read计数数据进行标准化,然后用DEGseq进行差异分析。为了获得显著差异的基因,设置筛选条件为:qValue<0.05和倍数差异|FoldChange|>2。显着差异表达转录本的GO和Pathway富集分析。
1.2.6 qRT-PCR验证 选择火龙果TBP2基因作为内参基因,通过转录组数据分析,结合KEGG数据库,筛选出甜菜途径中色素相关转录因子、高表达基因和关键酶等15个unigenes进行qRT-PCR验证筛选。引物设计使用引物5.0(表1),使用TB Green(TliRNaseH Plus)(Takara)试剂盒,将样品放入实时荧光定量PCR仪(Mx3000P,Stratagene,美国)中完成反应,为防止技术失误,每个反应做3次重复,结果用相对定量法分析(2–CT)。
2 结果与分析
2.1 转录组测序及质量评估
为了解析火龙果果肉着色机制,以红心火龙果‘美红一号’为研究对象,采集了果实9个不同发育时期(P0:小花蕾;P1:中花蕾;P2:大花苞;P3:开花期;P4:膨大期;P5破色期;P6:着色期;P7:果皮开始转色期;P8:果实完全成熟期)的果肉样品,进行转录组测序。利用CTAB法提取高质量RNA后构建插入片段为21 317 bp的cDNA文库(3次生物学重复),并利用PacBio RS sequel測序平台是基于单分子实时(SMRT)测序技术进行转录组测序。使用Trimmomatic对原始下机数据进行处理,去除原始数据中低质量、接头污染以及未知碱基N含量过高的reads;质控后得到24 275条高质量reads,平均每个样品的长度为2381.01。质量评估显示用于后续转录组分析的数据质量Q30大于91.93%(表2),表明这批转录组数据质量值较高,能够满足后续数据分析需求。
对PacBio测序原始数据进行评估。使用SMRT Link的Iso-Seq protocol进行全长转录本分析,使用LoRDEC软件利用Illumina二代数据对一致性转录本序列进行错误校正。其中去除reads长度小于35 nt的reads本身及其配对reads,且Q30均高于(91%),从整体上看,低质量(Quality<30)的碱基比例较低,说明测序质量较好。
2.2 基因功能注释结果
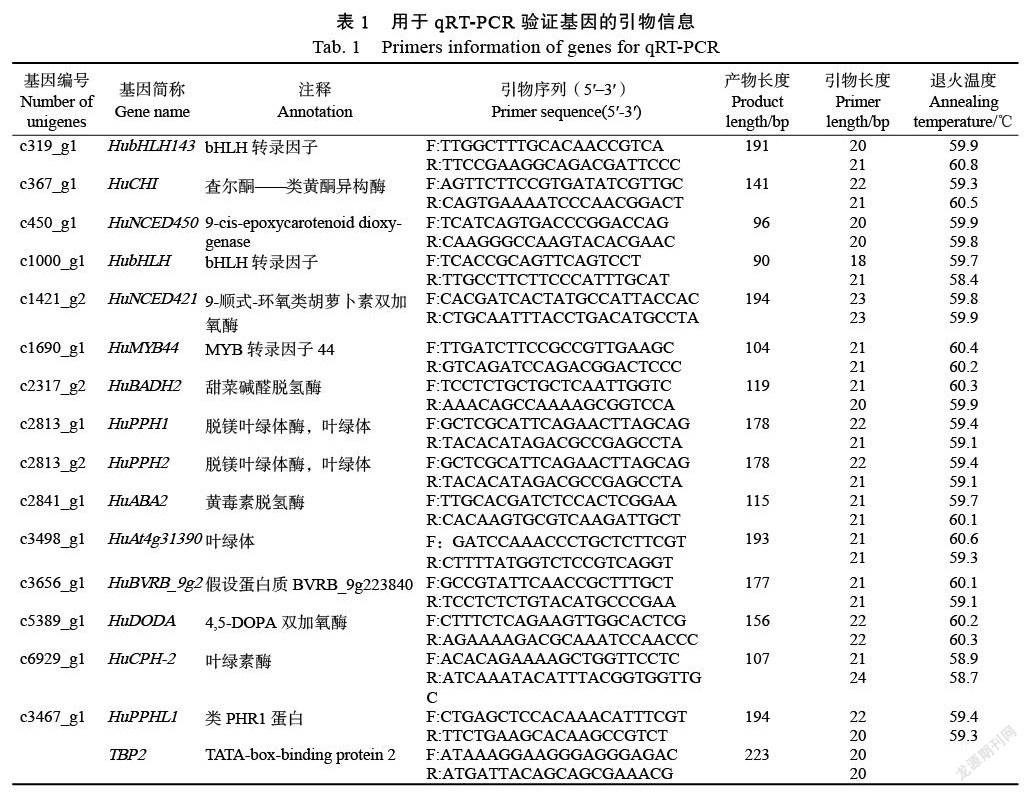
为了预测unigenes序列的功能信息,将24 275条unigenes序列分别在NR、SwissProt、KEGG和KOG数据库进行BLAST比对(图1)。根据NCBI Blast+,全长转录本模型与CDD、KOG、COG、NR、NT、PFAM、Swissprot、TrEMBL等多个数据库进行对比筛选后总共有4626条序列能够匹配到数据库中,占全部unigenes序列19%。其中,NR数据库4485条,Swissprot数据库4078条,KOG数据库3047条,KEGG数据库1592条,四大数据库同时注释1358条序列。
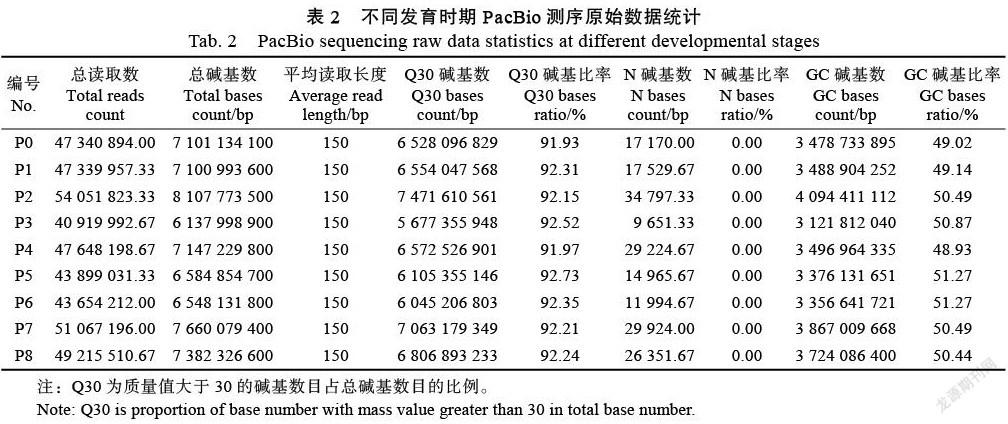
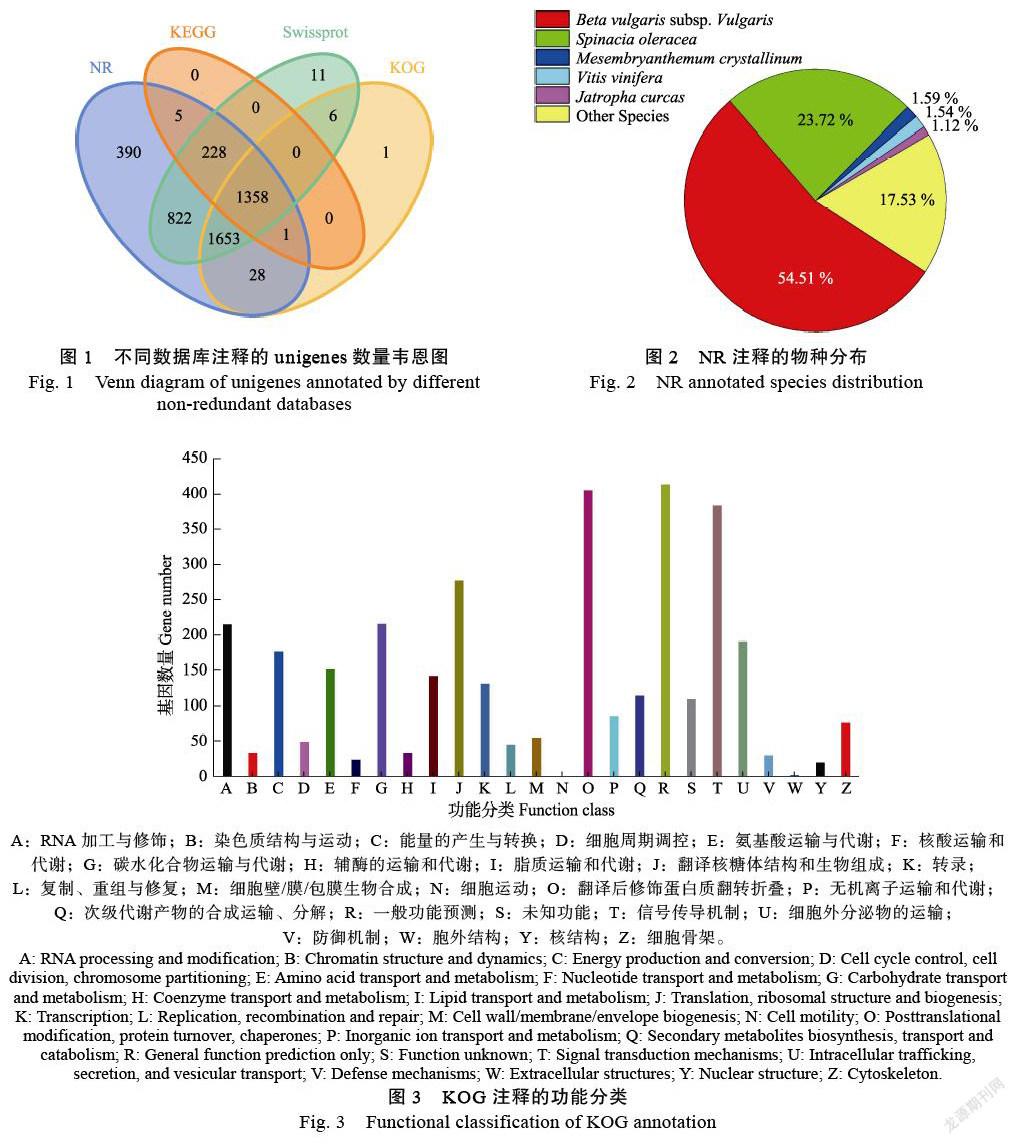
在NR数据库中进行物种比较分析(图2),共比较了132个物种。前五名是:甜菜2441条(Beta vulgaris subsp. Vulgaris:54.51%)、菠菜1062条(Spinacia oleracea:23.72%)、非洲冰草71条(Mesembryanthemum crystallinum:1.59%)和葡萄69(Vitis vinifera:1.54%)、桐油树50(Jatropha curcas:1.12%)。可以看出,火龙果和甜菜在基因序列上具有高度的同源性。KOG数据库中基因同源物的比较分析(图3),通过24个类别的比较,发现413个基因与一般功能预测相关,406个基因与翻译后修饰、蛋白质转运和分子伴侣相关十大类显示这些蛋白质主要与基因转录、翻译、蛋白质加工和细胞内转运相关,与次级代谢产物相关的基因共115个。
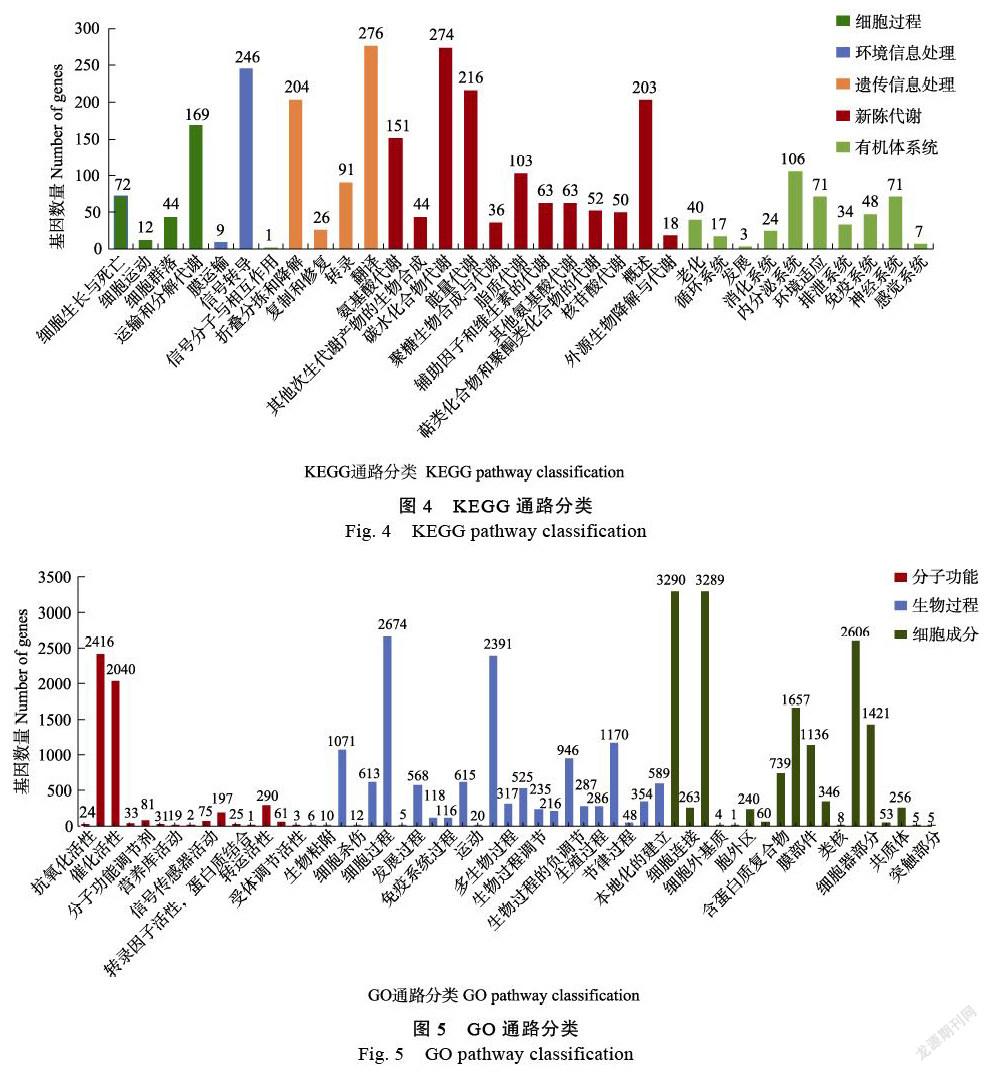
KEGG数据库共注释了2844个转录本。按代谢途径可分为5个分支,细胞过程297个、环境信息处理(environmental information processing)256个、遗传信息处理(genetic information processing)597个、代谢(metabolism)1273个、有机系统(organismal systems)421个碳水化合物代谢(274)和能量代谢(能量代谢)是代谢途径中涉及最多的途径,其次是概览图(246)。在KEGG代谢途径分析中(图4),更多的基因富集在次级代谢物合成途径、氨基酸和核酸代谢途径、酪氨酸代谢途径中。
GO(gene ontology)功能注释分析(图5)富集了所有样本的生物过程(biological process)、细胞成分(cellular component)和分子功能(molecular functon)三类基因,其中富集了更多的基因在细胞成分中,但分子功能较少。在生物过程中,2391个基因在代谢过程中富集,2674个基因在细胞过程中富集,1071个基因在生物调控和刺激反应中富集。这些过程与色素代谢密切相关。在细胞成分上,可以看出,细胞器或细胞器部分、细胞器或细胞器部分、膜或膜部分这3种类型的细胞或细胞器部分富含更多的基因;在分子功能方面,催化活性功能富集2040个基因,结合功能富集2416个,转运活性富集290个基因。此外,一些基因富含蛋白质和转录因子的结合活性。执行GO(基因本体)分析(图5)。在分子功能方面,4个部分明显富集到更多的基因,即结合、催化活性、转运活性、结构分子活性和色素代谢。
2.3 果实不同发育时期果肉组织代谢物含量与颜色变化
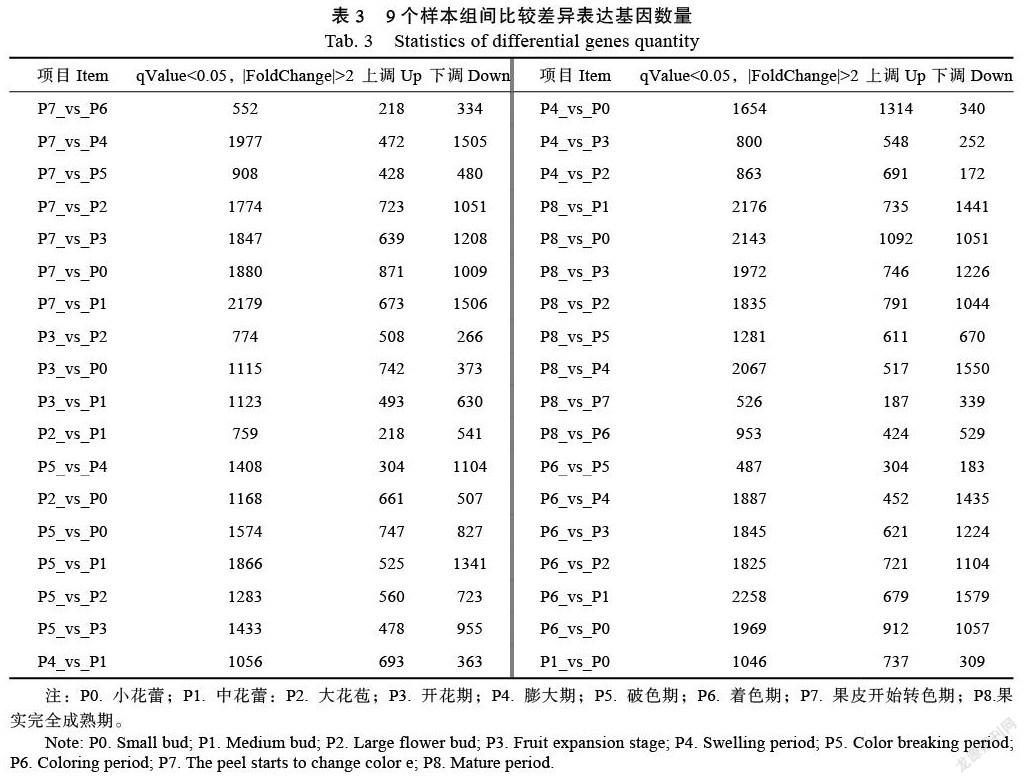
上述化合物中对火龙果果肉的颜色有影响的物质有花青苷含量、类胡萝卜素、类黄酮和甜菜色素,这些化合物都在火龙果不同的发育时期,其含量都有明显差异(图7),可以看出花色苷在P4和P5要明显高于其他发育时间,类胡萝卜素在P7的含量要明显高于其他发育时期。类黄酮在P4和P7的含量也要高于其他发育时期。甜菜色素在火龙果的P3~P6时期的含量较低,P7时期含量表达开始上调,P8时期含量最高。综上所述,火龙果果肉的颜色直观地由其所含的化学物质决定,不同的发育时期,火龙果果肉中的维生素C和甜菜色素均较高,导致果肉的颜色也在不同发育时期有差异(图6)。
2.4 果实不同发育时期果肉组织间差异表达基因(Differently expressed genes,DEGs)
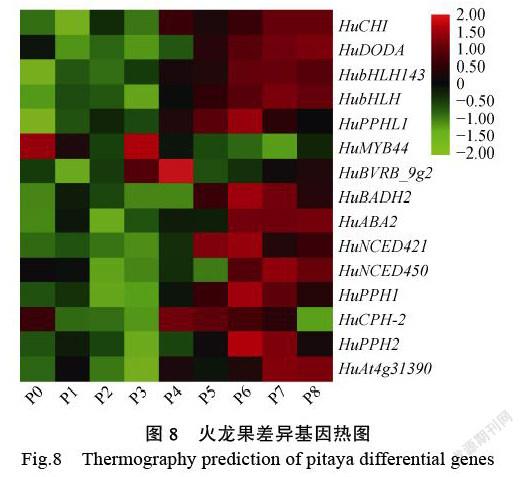
以qValue<0.05,|FoldChange|>2为标准,在9个样品中(表3),进行了成对比较筛选。可以看出,同一物种在不同时期中幼花幼果期(P0、P1、P2、P3)与成熟期(P6、P7、P8)比较的差异基因。结合果实不同发育时期化合物测定结果得出,在果实转色期(P4 vs P5)基因差异变化明显。在P4 vs P5果实转色期中,共有1408个基因显着表达,其中304个上调,1104个下调。DEG由GO(GeneOntology)分析。在分子功能上,富集到更多基因是在催化活性、连接、转运活性、核酸与转录因子结合活性、色素代谢等方面。在KEGG代谢途径分析中,更多的基因富集在次级代谢物合成途径、氨基酸和核酸代谢途径、酪氨酸代谢途径中。
2.5 差异表达基因qRT-PCR验证
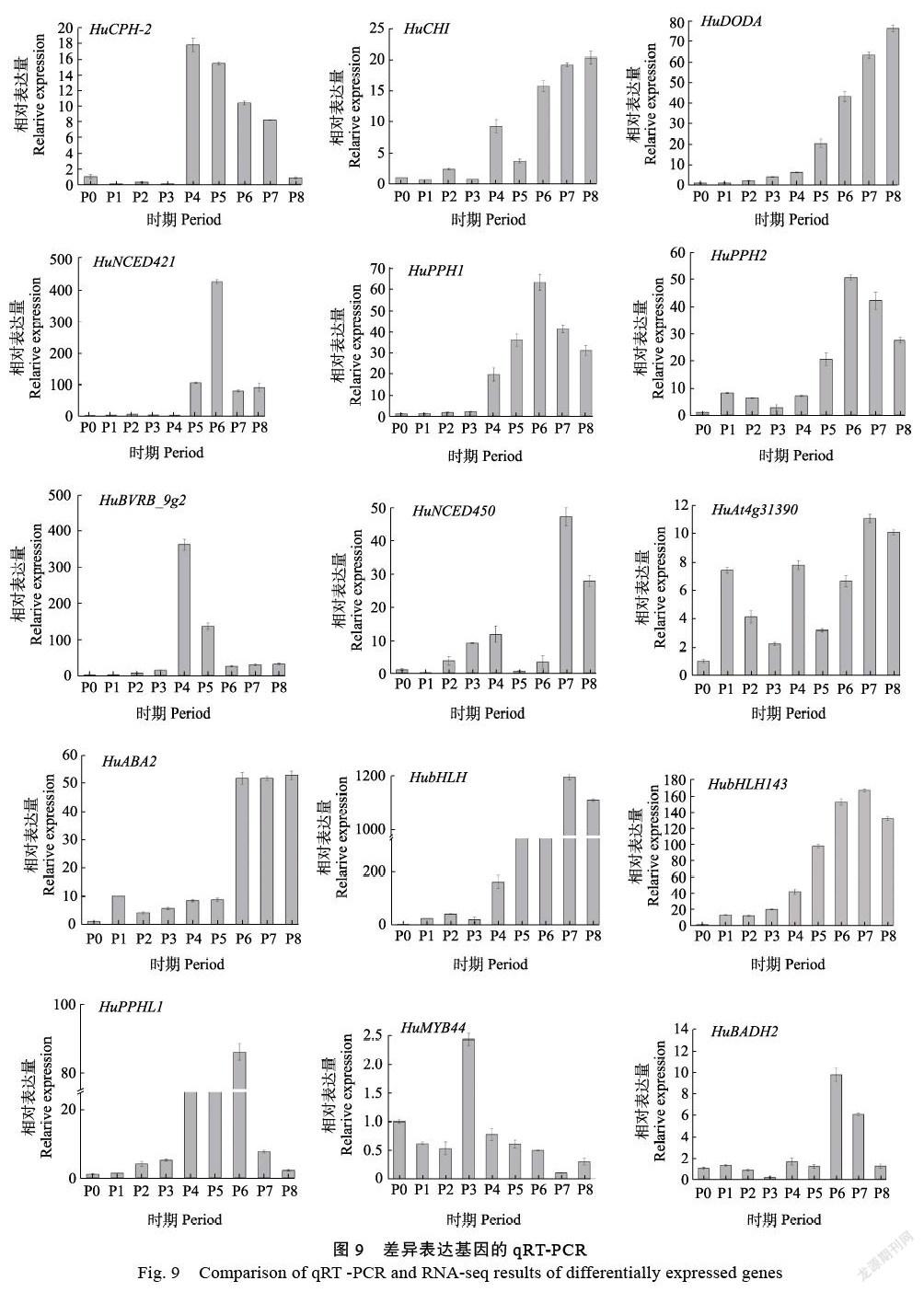
通过转录组分析结果分析,提取P0~P8全部发育时期的基因表达量绘制热图。根据(图8)中结果来看,这些基因在P0~P3时期表达量都较低,P4期果实中不同基因的表达量差异较大,P5~P8期基因普遍高表达。具体来说,HuCHI、HuNCED 450、HuABA2、HubHLH143和HubHLH在果实的整个发育后期P6~P8表达量开始上升;HuPPHL1、HuPPH1、HuPPH2、HuBADH2在果实发育后期P6~P7中表达量高;HuCPH-2在P4期高表达后又开始逐渐下降;HuAt4g31390在P7~P8期中表达量陡然增加;HuMYB44在果实发育前期表达量较高,到了后期表达量则下降,HuNCED421随着果实发育不断成熟,该基因的表达量在P5和P6时期出现高表达,果实成熟期又逐渐下降;HuDODA则随着时间累积,表达量一直增加。
为了转录组测序和分析结果的可靠性,进行了进一步的验证,使用荧光定量PCR对15个差异表达基因进行表达量验证(图9)。在P0~P8九个发育时期,这些基因均表现出不同程度的表达变化。其中HuBVRB_9g2在P4期表达量较高;HuNCED421在P6期出现了高表达;HuPPH1和HuPPH2在P6期出现高表达后,表达量又开始下降;HuDODA和HubHLH基因随着果实的发育表达量表现出明显上升趋势,尤其在P4时期表达量显著升高;HuABA2在P6~P8期表達量陡然增高。15个关键基因在9个发育时期的荧光定量结果与转录组分析结果一致,表明本研究分析和实验结果真实可靠。P4时期是果实转色前的关键时期,因此,找到火龙果果肉转色前后表达量显著升高的基因对研究果肉颜色形成的分子机制具有重要意义。
3 讨论
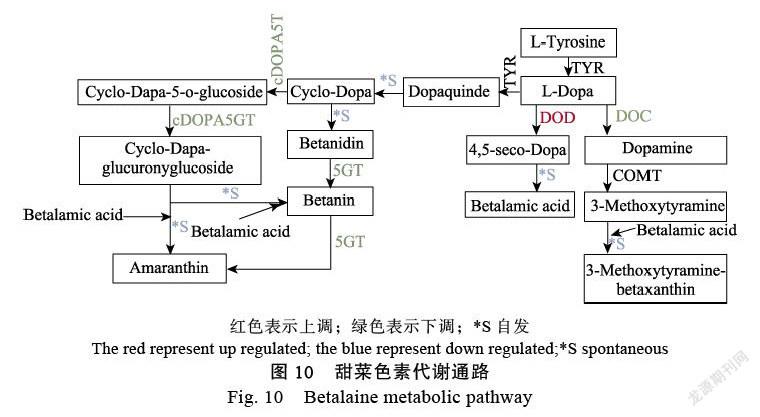
果肉的颜色变化是一个复杂的过程,需要各种基因、酶和转录因子的调控。还有许多外部环境影响,如温度、光线和其他自然因素。甜菜色素的合成前体在酪氨酸和其他酶的催化下形成甜菜醛氨酸(betalammic acid),其中共轭双键形成发色团[15]。另一方面,在细胞色素P450(CytP450)的催化下形成环多巴(cyclo-dopa)。环多巴自发形成甜菜碱。β苷元在GT等催化下与甜菜醛结合形成甜菜碱)[15-16],储存于液泡中[17](图10)。
本研究基于定量分析发现,结合转录组分析,DODA转录本HuDODA的结果呈现上调趋势。在甜菜色素的生物合成途径中,多巴(dopa)在4,5-多巴雌二醇双加氧酶等酶的催化下形成甜菜醛氨酸,其中共轭双键形成发色基团,而后通过自发反应形成甜菜色素[18]。1个HuCHI与黄酮类生物合成有关;3个(HuNCED450、HuABA2和HuNCED421)与类胡萝卜素生物合成有关;1个HuBADH2与甘氨酸、丝氨酸和苏氨酸代谢有关;1个(HuDODA)与DODA双加氧(DOD)甜菜生物合成相关,DODA是甜菜色素代谢途径中形成发色基团的关键酶,也是最有可能与甜菜色素关键基因表达相关的关键酶[16];3个(HuMYB44、HubHLH143和HubHLH)与色素相关的转录因子,4个(HuCPH-2、HuPPH1、HuAt4g31390和HuPPH2)卟啉与叶绿素代谢相关;2个(HuPPHL1和HuBVRB_9g2)预测蛋白类。在转录组数据代谢通路中发现(图10),3个关键酶的只有DODA表达一直处于上调趋势,并且该基因可能在果实临近采收时期高表达,其可能参与果实品质形成过程。
甜菜色素是一种水溶性天然植物色素,种类比较少。目前,已鉴定出75种甜菜色素。在不同pH下,植株呈现红、紫、黄、白等颜色,是火龙果颜色的重要组成部分[19-22]。据报道,在甜菜中,甜菜色素的合成需要4种酶的参与:酪氨酸酶[23-24]、4,5-DOPA双加氧酶[25]、细胞色素P450和葡萄糖碱基转移酶[26],火龙果中甜菜色素合成的分子调控机制尚未明确。
所有样品的GO分析显示,更多的基因富含催化活性、连接、转运活性、核酸和转录因子结合活性,这表明色素的代谢需要多种酶和调节因子。对9个样品进行配对筛选,发现同一品种不同时期(P8VSP1)(P6VSP1)的差异表达基因较多,说明火龙果肉的色素差异较大。与果实的发育阶段有关。在果实的生长发育过程中,一些与颜色相关的关键基因可能会继续发挥促进甜菜色素合成的作用。通过KEGG分析发现,大多数富集的酶与酶活性密切相关。同时,代谢途径和酪氨酸代谢途径中富含的酶较多。甜菜色素合成的第一个底物是L-酪氨酸合成的次级代谢产物[27-28],发育过程中可能存在与果实颜色发育相关的基因。赵思军的研究表明苯丙氨酸是酪氨酸合成的前体[29]。黄酮和黄酮醇的合成与黄酮色素的合成密切相关。火龙果果实颜色的成分较为复杂,不仅包括甜菜色素,还可能含有花青素等黄酮醇色素,但据报道,花青素和甜菜色素不能共存[30]。郭攀阳等[31]对2个不同品种火龙果的果肉颜色进行转录组测序,通过DEGs分析发现,同一品种不同时期火龙果的差异比同一时期不同品种间的更大。因此可以从中挑选差异倍数较高的,在P8样本中表达较为显著的基因进行深入研究。
参考文献
[1] 巴良杰, 罗冬兰, 戴卫军, 等. 红肉火龙果HpERF1/2/3转录因子的特性及对HpCytP450-like1的调控[J]. 保鲜与加工, 2018, 18(1): 32-40.
[2] 吴 琳. 火龙果种质资源评价及繁育研究[D]. 南宁: 广西大学, 2016.
[3] 蔡永强, 向青云, 陈家龙, 等. 火龙果的营养成分分析[J]. 经济林研究, 2008, 26(4): 53-56.
[4] 赵湖冰, 姚媛媛, 杨乐乐, 等. 火龙果研究现状与开发前景[J]. 山东化工, 2018, 47(14): 52-54.
[5] 申世辉, 马玉华, 蔡永强. 火龙果研究进展[J]. 中国热带农业, 2015(1): 48-52.
[6] Xi X, Zong Y, Li S, et al. Transcriptome analysis clarified genes involved in betalain biosynthesis in the fruit of red pitayas (Hylocereus costaricensis) [J]. Molecules, 2019, 24(3): 445.
[7] 刘小玲, 许时婴, 王 璋. 火龙果色素的基本性质及结构鉴定[J]. 无锡轻工大学学报, 2003(3): 62-66+75.
[8] 梁洁玲, 李芸瑛, 罗文娟, 等. 火龙果色素的提取工艺优化及色素稳定性研究[J]. 中国食品添加剂, 2018(11): 73-77.
[9] 段晓嫣, 田 艳, 邓放明. 火龙果色素生物活性及其提取纯化研究进展[J]. 食品与机械, 2017, 33(10): 214-219.
[10] 唐 霓, 徐 宁, 韦敏雪, 等. 火龙果色素的稳定性及应用研究[J]. 现代食品, 2017(9): 115-117.
[11] 许 哲, 杨丹娇, 李修辉, 等. 火龙果溃疡病防治药剂的筛选[J]. 植物医生, 2021, 34(01): 72-75.
[12] 陸 漫, 王小云, 郑霞林, 等. 我国火龙果病虫害绿色防控技术研究进展[J]. 广西植保, 2020, 33(01): 28-31.
[13] 曾灿彬. 红肉与白肉火龙果果实发育的色素比较与差异表达基因研究[D]. 海口: 海南大学, 2017.
[14] 刘春晓, 黄小庆, 刘自广, 等. 十字花科植物种子低分子RNA提取方法比较[J]. 基因组学与应用生物学, 2019, 38(3): 1236-1241.
[15] Polturak G, Heinig U, Grossman N, et al. Transcriptome and metabolic profiling provides insights into betalain biosynthesis and evolution in Mirabilis jalapa[J]. Molecular Plant, 2018, 11(1): 189-204.
[16] Wu Y, Xu J, He Y, et al. Metabolic profiling of pitaya (Hy-locereus polyrhizus) during fruit development and matura-tion[J]. Molecules, 2019, 24(6): 1114.
[17] Chen N, Yu Z H, Xiao X G. Cytosolic and nuclear co-localization of betalain biosynthetic enzymes in tobacco suggests that betalains are synthesized in the cytoplasm and/or nucleus of betalainic plant cells[J]. Frontiers in Plant Science, 2017, 8: 831.
[18] Fischer N, Dreiding A S. Biosynthesis of betalaines. on the cleavage of the aromatic ring during the enzymatic transformation of dopa into betalamic acid[J]. Helvetica Chimica Acta, 1972, 55(2): 649-658.
[19] Lee E J, An D, Nguyen C T T, et al. Betalain and betaine composition of greenhouse-or field-produced beetroot (Beta vulgaris L. ) and inhibition of HepG2 cell proliferation[J]. Journal of Agricultural and Food Chemistry, 2014, 62(6): 1324-1331.
[20] Clifford T, Howatson G, West D J, et al. The potential benefits of red beetroot supplementation in health and disease[J]. Nutrients, 2015, 7(4): 2801-2822.
[21] Martinez R M, Longhi-Balbinot D T, Zarpelon A C, et al. Anti-inflammatory activity of betalain-rich dye of Beta vul-garis: effect on edema, leukocyte recruitment, superoxide anion and cytokine production[J]. Archives of Pharmacal Research, 2015, 38(4): 494-504.
[22] Hua Q, Chen C, Chen Z, et al. Transcriptomic analysis reveals key genes related to betalain biosynthesis in pulp coloration of Hylocereus polyrhizus[J]. Frontiers in Plant Science, 2016, 6: 1179.
[23] Tanaka Y, Sasaki N, Ohmiya A. Biosynthesis of plant pig-ments: anthocyanins, betalains and carotenoids[J]. The Plant Journal, 2008, 54(4): 733-749.
[24] Sunnadeniya R, Bean A, Brown M, et al. Tyrosine hydrox-ylation in betalain pigment biosynthesis is performed by cytochrome P450 enzymes in beets (Beta vulgaris)[J]. PloS One, 2016, 11(2): e0149417.
[25] Hinz U G , Fivaz J , Girod P A , et al. The gene coding for the DOPA dioxygenase involved in betalain biosynthesis in Amanita muscaria and its regulation. [J]. Molecular & General Genetics Mgg, 1997, 256(1): 1-6.
[26] Fischer N, Dreiding A S. Biosynthesis of betalaines. on the cleavage of the aromatic ring during the enzymatic transformation of dopa into betalamic acid[J]. Helvetica Chimica Acta, 1972, 55(2): 649-658.
[27] Gao Z J, Han X H, Xiao X G. Purification and characterisa-tion of polyphenol oxidase from red Swiss chard (Beta vul-garis sub species cicla) leaves[J]. Food Chemistry, 2009, 117(2): 342-348.
[28] Wang C Q, Song H, Gong X Z, et al. Correlation of tyrosi-nase activity and betacyanin biosynthesis induced by dark in C3 halophyte Suaeda salsa seedlings[J]. Plant Science, 2007, 173(5): 487-494.
[29] 赵思俊, 赵晓喆, 向 欢, 等. 基于代谢通路调控的沙棘籽油抗抑郁作用机制研究[J]. 中草药, 2017, 48(13): 2682-2690.
[30] Fan R, Sun Q, Zeng J, et al. Contribution of anthocyanin pathways to fruit flesh coloration in pitayas[J]. BMC Plant Biology, 2020, 20(1): 1-12.
[31] 郭攀陽, 曾灿彬, 刘成立, 等. 红皮红肉和红皮白肉火龙果果肉呈色相关基因差异表达分析[J]. 分子植物育种, 2021, 19(13): 4311-4326.
责任编辑:沈德发
猜你喜欢
领导文萃(2021年8期)2021-06-08
语文周报·教研版(2020年8期)2020-05-13
农家参谋(2019年5期)2019-09-10
养生大世界(2019年9期)2019-09-05
世界热带农业信息(2018年4期)2018-10-25
少年文艺·开心阅读作文(2018年9期)2018-09-28
恋爱婚姻家庭·养生版(2018年8期)2018-08-18
世界热带农业信息(2016年7期)2016-08-23
故事作文·低年级(2016年3期)2016-04-08
世界热带农业信息(2016年3期)2016-04-05
