
东京野茉莉对重金属胁迫的响应特征研究
2021-11-28 11:10王丽艳吴巧花
南方林业科学 2021年5期
王丽艳,杨 桦,周 晨,吴巧花
(江西省林业科学院·南昌城市生态系统国家定位观测研究站,江西 南昌330013)
随着现代工业的迅猛发展,铜和镉被广泛应用于机械、化工、电镀、印染等部门,这些工业排放的废弃物成为环境中的重金属污染源。由于污染源的广阔性,使城市土壤中重金属已有不同程度的累积[1-2]。重金属对植物的伤害前人已做了很多研究,如铜能破坏植物体光合器官和叶绿体结构[3-4];镉能抑制叶绿素的生物合成[5],也会影响PSII反应中心和电子传递、卡尔文循环中有关酶的活性导致一系列生理代谢紊乱,从而减少生物量积累[6]。但这些研究主要集中在农业及草本植物上,土壤中的重金属是如何影响东京野茉莉(Styrax tonkinensis)的光合作用和叶绿素特性报道不多。本实验以东京野茉莉苗木为对象,研究不同浓度土壤铜和镉污染胁迫下其光合作用和叶绿素的变化,旨在分析铜和镉污染胁迫对光合作用影响的内在机制。
1 试验设计与方法
1.1 试验设计
对东京野茉莉分别进行铜(Cu)、镉(Cd)2种胁迫处理,并设置空白对照。空白处理1组共10盆,不处理,正常浇水。铜处理,共4组,每组10盆,铜胁迫用CuSO4溶液,每组浇灌浓度分别为:0 mmol·L-1、1 mmol·L-1、5 mmol·L-1、10 mmol·L-1、20 mmol·L-1。镉处理,共4组,每组10盆,镉胁迫用CdCl2溶液,每组浇灌浓度分别为:0 μmol·L-1、100 μmol·L-1、200 μmol·L-1、400 μmol·L-1、600 μmol·L-1。从2016年7月开始定期浇灌不同浓度的CuSO4和CdCl2溶液,浇灌的体积为100 mL,浇灌到11月底。浇灌时,将各浓度的重金属溶液用量筒量好,将溶液缓缓流入营养钵的土壤中,以防淋溶出盆。并在每次浇灌时(时间间隔7 d)记录1次每株东京野茉莉叶子的叶绿素(SPAD值),每浇灌2次记录1次每株东京野茉莉的株高和地径。
1.2 测定方法
1.2 .1 幼苗叶片叶绿素含量的测定
对4种不同浓度Cu、Cd的幼苗采摘一定量叶片供试。叶绿素含量参考《植物生理生化实验原理和技术》中的方法[7]。①在3个研钵中分别加入3份称取好了的剪碎的新鲜样品,在各研钵中加入95%乙醇2~3 mL和少量石英砂和碳酸钙粉,研成均浆,然后再加95%乙醇10 mL,继续研磨至组织变白。静置3~5 min。②将一张滤纸放入漏斗中,并且用乙醇润湿,将提取液沿玻璃棒倒入漏斗中,过滤到25 mL的棕色容量瓶中,提取液倒入容量瓶之后,再用少量乙醇冲洗研钵、研棒及残渣数次,最后连同残渣一起倒入漏斗中。③用塑料滴管吸取少量的乙醇,把滤纸上的叶绿体色素全部冲洗到容量瓶中。直至滤纸和残渣中无绿色为止。最后用乙醇定容至50 mL,摇匀。然后再将叶绿体色素提取液倒入直径1 cm的比色皿内。以95%乙醇为空白,在波长665 nm、649 nm、470 nm下测定吸光度。
1.2 .2 叶片光合参数的测定
试验初期选取各处理生长良好的4株苗木作为固定样株,测量时从顶叶向下第3~5片功能叶中选取叶片,利用Li-6400型便携式光合测定仪(Li-Cor,Inc,USA),在晴天的9:00-17:00进行光合参数测定,设定样本室温温度25℃,取自相对稳定的3 m高空,空气流速为400~410 μmol·s-1。选择长势良好、无病虫害和无机械损伤的幼苗,每株幼苗自上而下选取3片健康叶片作为测定叶,每个叶片待数据稳定后连续取3个数据,取平均值。仪器自动记录净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)等参数。
2 结果与分析
2.1 不同浓度Cu、Cd胁迫下东京野茉莉幼苗叶绿素含量的变化
2.1 .1 Cu胁迫下东京野茉莉幼苗叶绿素含量的变化
由表1可知,在Cu处理浓度为1 mmol·L-1时,东京野茉莉幼苗中叶绿素a、叶绿素b、类胡萝卜素含量及叶绿素总含量与对照组均不显著。但当Cu处理浓度为5 mmol·L-1、10 mmol·L-1、20 mmol·L-1时,叶绿素a、叶绿素b含量均显著低于对照组。类胡萝卜素在也受到了Cu胁迫影响,随Cu浓度增加类胡萝卜素含量先降低后上升,在10 mmol·L-1、20 mmol·L-1Cu浓度处理水平,类胡萝卜素含量显著高于对照组。随着Cu浓度增加叶绿素含量先升后降,与对照组差异不显著(P<0.05)。

表1 不同浓度Cu处理对幼苗叶绿素的影响Tab.1 Effects on chlorophyll of seedlings to different concentrations of Cu stress
2.1 .2 Cd胁迫下东京野茉莉幼苗叶绿素含量的变化
表2 表明,在Cd处理浓度为100 μmol·L-1、200 μmol·L-1时,东京野茉莉幼苗中叶绿素a、叶绿素b、类胡萝卜素含量及叶绿素总含量均显著高于对照,说明低浓度的Cd胁迫对东京野茉莉幼苗中叶绿素a、叶绿素b、类胡萝卜素含量及叶绿素总含量有一定的促进作用。随着Cd浓度的增加,东京野茉莉幼苗中叶绿素a、叶绿素b含量及叶绿素总含量呈下降趋势,叶绿素a、叶绿素b含量均与对照组差异显著,说明高浓度的Cd胁迫会对叶绿素含量变化起到抑制作用。而余东等[8]的研究结果表明低浓度和高浓度的Cd胁迫对枇杷叶绿素含量和光合作用均显示抑制作用,说明不同树种对Cd胁迫的响应表现不一致。本研究表明类胡萝卜素呈先降后升趋势,与对照组差异显著(P<0.05)。
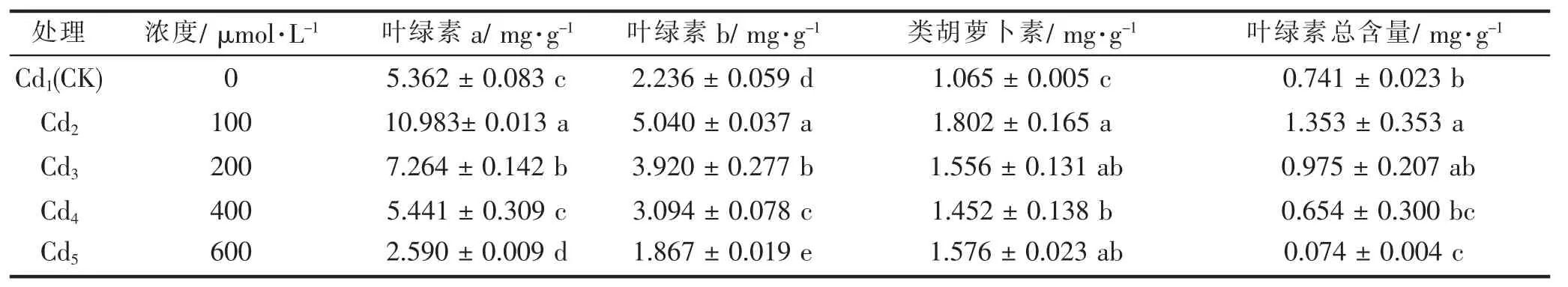
表2 不同浓度Cd处理对幼苗叶绿素的影响Tab.2 Effects on chlorophyll of seedlings to different concentrations of Cd stress
2.2 不同浓度的Cu、Cd胁迫对东京野茉莉的光合作用的影响
2.2 .1 Cu胁迫对东京野茉莉的光合作用的影响
根据表3可知,与对照组相比,不同Cu浓度处理的东京野茉莉幼苗净光合速率、气孔导度、蒸腾速率和胞间CO2浓度都有所下降。说明Cu对植物的光合作用有着抑制作用。根据多重比较结果,不同浓度Cu的处理对东京野茉莉净光合速率差异显著(Ρ<0.05)。由表3可知,1 mmol·L-1与5 mmol·L-1Cu浓度处理的数据结果无显著差异,与10 mmol·L-1、20 mmol·L-1Cu浓度处理存在显著差异;10 mmol·L-1与20 mmol·L-1Cu浓度处理相比较无显著差异,与0 mmol·L-1、1 mmol·L-1、5 mmol·L-1Cu浓度处理的结果存在显著差异。说明随着Cu处理浓度的升高,净光合速率呈下降趋势,不同浓度Cu处理均对东京野茉莉幼苗净光合速率起抑制作用。对于气孔导度、蒸腾速率、胞间CO2浓度,各组不同浓度Cu处理间均存在显著差异,随着浓度的增加呈先下降后上升再下降的趋势,均在浓度为5 mmol·L-1时达到最大值,说明在特定浓度下,Cu胁迫对东京野茉莉的气孔导度、蒸腾速率、胞间CO2浓度有增强作用。
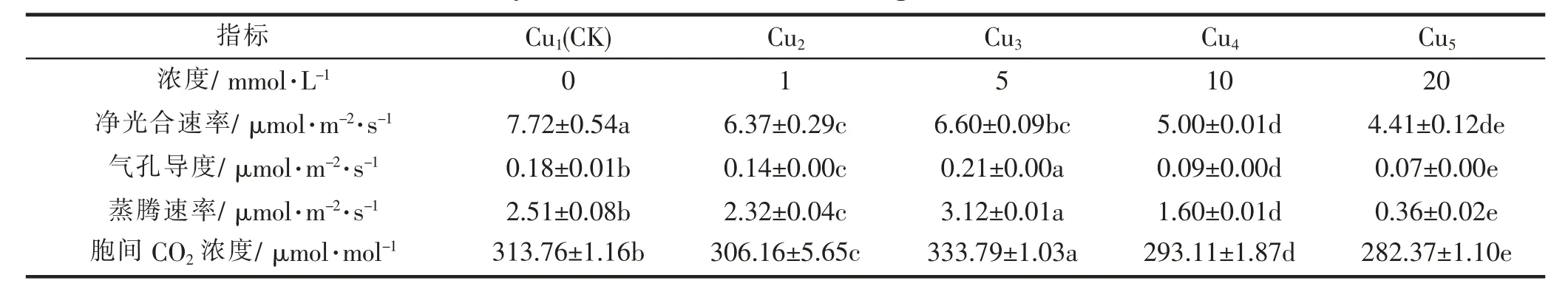
表3 不同浓度的Cu对东京野茉莉的光合作用的影响Tab.3 Effects on Photosynthesis of S.tonkinensis seedlings different concentrations of Cu stress
2.2 .2 Cd胁迫对东京野茉莉的光合作用的影响
根据表4可知,与对照组相比,100~400 μmol·L-1浓度Cd处理下,东京野茉莉幼苗净光合速率受到抑制作用,当浓度达到600 μmol·L-1时,其净光合速率高于对照组,说明一定高浓度的Cd处理,对东京野茉莉的幼苗净光合速率具有增强作用。田晓峰[9]的研究结果中,重金属Cd对金丝柳净光合作用速率有随重金属Cd浓度的升高而降低的趋势,与本文Cd对东京野茉莉的处理结果不太一致,说明不同树种对Cd胁迫的响应存在一定差异。

表4 不同浓度的Cd对东京野茉莉的光合作用的影响Tab.4 Effects on Photosynthesis of Styrax tonkinensis seedlings different concentrations of Cd stress
对于气孔导度和蒸腾速率而言,100 μmol·L-1和200 μmol·L-1浓度Cd处理与对照间无显著差异,当浓度为400 μmol·L-1时,东京野茉莉的气孔导度和蒸腾速率均产生较大幅度下降,而浓度达到600 μmol·L-1时,气孔导度和蒸腾速率均达到最大值,显著大于其它处理组和对照组。对于胞间CO2浓度而言,除了400 μmol·L-1浓度处理抑制了东京野茉莉的胞间CO2浓度,其它处理组均显著大于对照组的胞间CO2浓度,浓度为200 μmol·L-1处理下,达到最大值。
大多数研究表明,重金属Cd随着浓度的升高,对植物光合作用产生抑制现象,李亚藏等[8]研究Cd对茶条槭和五角槭光合作用影响表明:随着土壤中镉浓度的增加,茶条槭的净光合速率、气孔导度和蒸腾速率呈先升后降趋势,胞间CO2浓度则呈先降后升趋势。但在本试验中,东京野茉莉在Cd的胁迫下,净光合速率呈先降后升趋势,气孔导度、蒸腾速率及胞间CO2浓度呈先升后降再升的趋势。在秦天才等[9]研究Cd对植物光作用的影响中得出,Cd对植物光合速率的影响受不同植物种类、不同的发育阶段以及外界的各类条件而定。
3 结论与讨论
3.1 讨论
Cd是生物体中的非必需元素,Cu是生物生长所需要的微量元素。重金属胁迫能使植物光合作用减弱,破坏叶绿素结构引起叶绿素总量降低。朱宇林等[12]学者研究表明,在重金属Cd和Pb的胁迫下,银杏叶片随着重金属浓度的升高,光合作用和蒸腾速率下降,且叶绿素a/b值也下降,其原因是重金属胁迫导致非气孔限制。
孟丽[13]研究发现,重金属对光合特性的影响受胁迫时间和浓度的影响,当处理浓度较低时,光合作用增强,超过一定浓度,光合作用受到抑制。同理说明对植物生长发育也起到抑制作用。本试验发现,东京野茉莉光合作用参数值的变化趋势均在低浓度Cd处理下有明显的上升趋势,与周怡[14]的研究结果一致,这表明低浓度的Cd能促进东京野茉莉的光合作用,但在高浓度下又会出现一定的抑制作用,但抑制作用是有限的,可能是研究中设置的Cd浓度还没能达到严重抑制东京野茉莉光合作用的处理水平,说明在Cd胁迫下,东京野茉莉具有较强的光合适应性。
光合速率是反映在不同环境下植物光合作用运转状况的灵敏指标,净光合速率可直接体现光合系统功能,可作为衡量植株系统工作正常与否的重要指标[15-16]。有研究表明,高浓度Cu胁迫会严重影响大花萱草(Hemerocallis fulva)植株的正常生长,主要原因可能是是植物为了减少水分散失,缩小或关闭气孔,降低气孔导度,减少叶片的气体交换[17]。关梦茜[18]对2种萱草对重金属Cu、Cd胁迫的生理响应研究发现,随着Cu胁迫浓度的增大,净光合速率降低会导致吸碳、释氧量降低,蒸腾速率的降低会导致降温、增湿的生态效益降低。本试验发现Cu胁迫下,东京野茉莉光合作用减弱,与对照差异显著(P<0.05),说明Cu胁迫下东京野茉莉的光合作用受到抑制。
3.2 结论
东京野茉莉幼苗在Cd、Cu的胁迫下,随着处理浓度的升高,叶绿素含量先上升后下降。随着Cd浓度的升高,东京野茉莉植株的净光合速率由下降到上升,气孔导度、蒸腾速率、胞间CO2浓度都有所上升,总体来说东京野茉莉在不同浓度Cd胁迫下,光合特性受抑制规律不明显,Cu胁迫对东京野茉莉光合作用具有抑制作用。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
农业与技术(2020年12期)2020-07-04
中国农业科技导报(2020年12期)2020-03-15
新课程·下旬(2017年7期)2017-08-14
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
湖北农业科学(2014年13期)2014-08-28
