
小麦籽粒品质空间分布异质性及其形成机制研究进展
2021-11-19 04:54姜东仲迎鑫蔡剑周琴王笑戴廷波曹卫星
南京农业大学学报 2021年6期
姜东,仲迎鑫,蔡剑,周琴,王笑,戴廷波,曹卫星
(南京农业大学农业农村部小麦区域技术创新中心,江苏 南京 210095)
小麦是我国最主要的粮食作物之一,也是广受人们喜爱的主食品种。目前,我国小麦生产总量已完全满足国内需求,但品质提升不足,难以满足人们提高生活质量的需求,导致低质小麦大量压库,而优质小麦原料仍需大量进口。研究人员根据消费习惯将我国小麦分为强筋(面包类、蛋白质含量相对较高)、中筋(蒸煮类、蛋白质含量中等)和弱筋(饼干糕点类、蛋白质含量相对较低)3个品质专用类型。胡学旭等[1]分析了2006—2015期间我国小麦主产区742个品种的7 561份样品,发现选育和推广的强筋和弱筋小麦品种数量在不断增加,但优质强筋和弱筋小麦原粮达标率分别仅为4.8%和0.4%,这也导致我国优质小麦进口增长迅速。据中国产业信息网数据,2011—2013年我国进口小麦由125万t增加到553万t,2014—2019年优质小麦进口量维持在350万t左右(https://www.chyxx.com/industry/202004/851643.html)。而2020年新型冠状病毒肺炎疫情暴发后,很多国家主动关闭了粮食出口通道,完全依靠进口来解决我国优质小麦需求的风险极大。因此,小麦品质改良与产量提升成为我国小麦生产的重要任务之一。
我国小麦品质形成机制与调优技术研究始于20世纪80年代,远迟于美国、加拿大、澳大利亚及欧洲等主要小麦出口国。长期以来,我国小麦生产中需要施用足量的氮肥以实现较高的产量。在适量范围内增施氮肥确实可显著提高籽粒蛋白质含量,有利于强筋和中筋小麦品质的提高,但同时带来氮素利用效率下降及生态污染等问题[2]。弱筋小麦的产量和品质协调则面临更大的挑战,因为增施氮肥会导致籽粒蛋白质含量偏高,严重降低弱筋小麦品质[3]。在适于生产优质弱筋小麦的长江中下游麦区,小麦物化生产成本为 6 750~7 000元·hm-2,再加上土地租金和人工成本,产量要超过6 000 kg·hm-2才能有一定的收益,而在这个产量水平下,弱筋小麦的籽粒蛋白质含量极易超标,导致弱筋小麦产量与品质间的矛盾突出。因此,急需寻求新的研究思路,探究在控制氮肥投入的情况下,提高中强筋小麦面粉蛋白质含量,保证中强筋小麦品质;以及优化栽培技术措施实现弱筋小麦产量和品质协同提升。
决定小麦品质和产量最重要的化学组分——蛋白质和淀粉在籽粒不同部位的含量差异显著(图1),并且籽粒不同部位蛋白质含量和加工品质对施氮等栽培措施的响应也明显不同[4]。在常用的制粉工艺中,约占30%的果皮、种皮、糊粉层、胚、部分外胚乳等籽粒外层部位会作为麸皮被去除掉。这为通过栽培措施调控小麦籽粒产量和品质协调提供了新的思路:对于中强筋小麦,可以选择外层面粉蛋白质含量对施氮响应不敏感、氮素主要在籽粒内层积累的品种,在有效提高面粉蛋白含量同时提高氮肥利用效率;而对于弱筋小麦,可选择内层面粉蛋白质含量对施氮响应不敏感、氮素主要在籽粒外层积累的品种,这样可以在适当增施或控制氮肥用量实现高产的同时,稳定甚至降低内层胚乳蛋白质含量,保证弱筋小麦籽粒蛋白含量不超标,实现丰产提质同步。因此,本文综述了小麦籽粒主要化学组分和加工品质空间分布的异质性及其生理调节机制,品质空间分布异质性对栽培措施的差异响应等方面的研究进展,提出了未来需重点突破的方向,为进一步深入阐明小麦产量品质协同机制、创新优质小麦产量品质协同提升栽培技术提供新的思路。
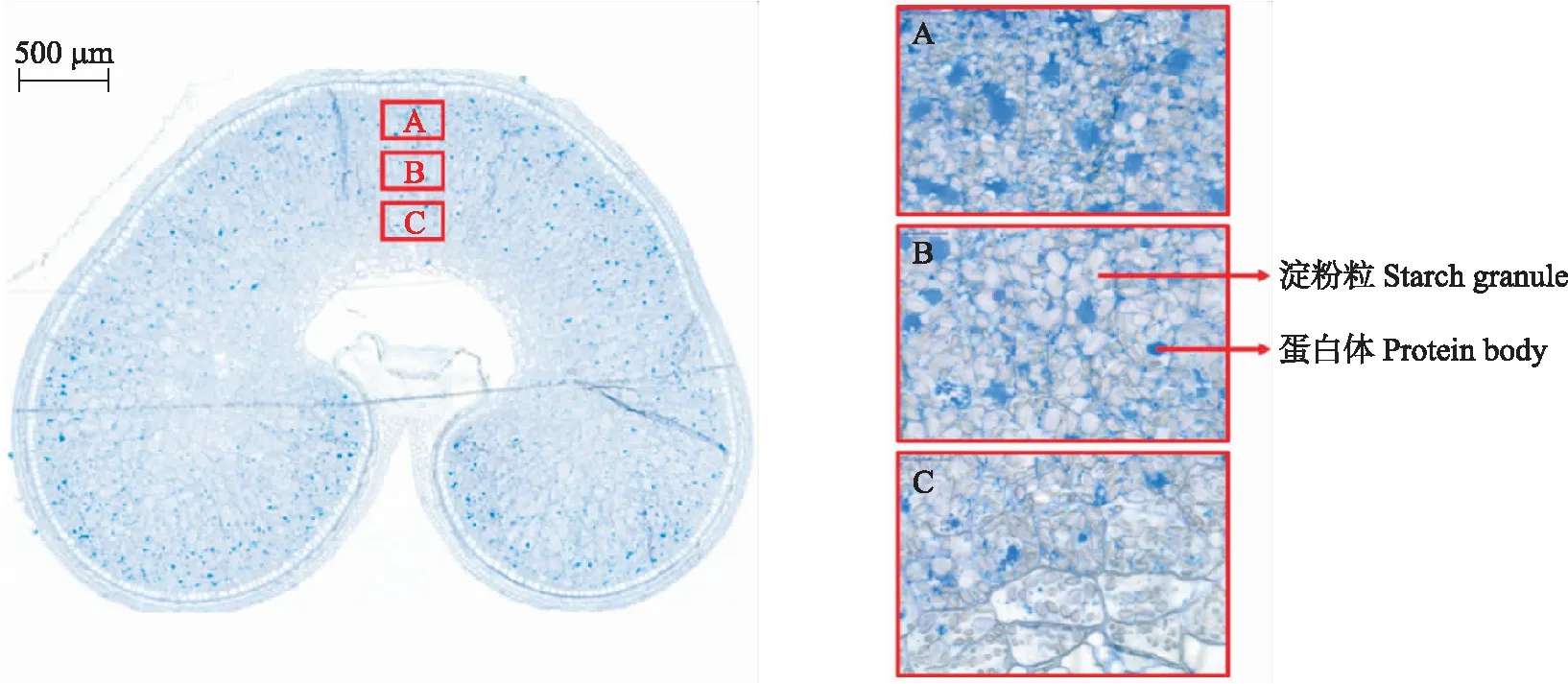
图1 甲苯胺蓝染色的灌浆期籽粒横切面Fig.1 Cross section of a developing grain stained with toluidine blue
1 小麦籽粒化学组分与加工品质空间分布异质性
小麦籽粒由胚乳、胚和麸皮组成,其中胚乳约占82%,胚约占3%,麸皮约占15%[5]。麸皮由外果皮、内果皮、外种皮、珠心表皮和糊粉层组成,其中糊粉层是包裹胚乳的最外层细胞,约占麸皮重的50%[6]。小麦籽粒含量最高的化学组分是淀粉,约占粒重的70%;其次为蛋白质,约占8%~20%[7],这2种组分在很大程度上决定了小麦籽粒的产量和品质。此外,籽粒中还包含很多对人体有益的营养物质,如膳食纤维、微量元素、植物生物活性物质及类胡萝卜素等[8],各组分在籽粒不同部分的分布均有显著差异。
1.1 蛋白质空间分布规律
关于籽粒不同部位蛋白含量的差异性早有报道,Kent[9]测定了亚糊粉层和内胚乳部位磨制面粉中的蛋白含量,发现亚糊粉层蛋白含量约为内胚乳的3倍。近年来,随着分层磨粉技术的发展,从外向内可大致获取麸皮(10%)、糊粉层(10%)、亚糊粉层(10%)、外胚乳(20%)、中胚乳(20%)、内胚乳(30%)部位的面粉[4]。对每一层的蛋白含量进行测定,发现蛋白含量呈现单峰分布,从麸皮层到内胚乳呈现先上升后下降的趋势,在糊粉层和外胚乳层的含量最高,不同品质类型小麦品种均表现出一致的规律[10]。
根据蛋白质在不同溶剂中的溶解度可进一步将其分为4类:溶于水的清蛋白、溶于100 g·L-1氯化钠溶液的球蛋白、溶于70%乙醇溶液的醇溶蛋白、溶于2 g·L-1氢氧化钠溶液的谷蛋白[11]。清蛋白和球蛋白被称为非面筋蛋白或者结构蛋白[12],主要作为酶参与各种代谢,对蛋白质加工品质影响较小;醇溶蛋白和谷蛋白被称为贮藏蛋白或面筋蛋白,可在淀粉粒周围形成连续的网络结构,构成面筋的主体[13]。关于不同层次面粉蛋白组分含量的差异已有较多报道。清蛋白和球蛋白的含量从外层到内层呈现逐层下降的趋势[4,14],结构蛋白主要存在于小麦籽粒的外层[12]。醇溶蛋白、麦谷蛋白这2种贮藏蛋白组分含量从外层到内层则呈先上升再下降的趋势,在籽粒外胚乳层达到峰值,内胚乳层最低[14-15]。
谷蛋白大分子聚合物(GMP)是不溶于十二烷基硫酸钠(SDS)的大型麦谷蛋白聚合物之一[16],与小麦面粉的烘焙性能高度相关。经聚丙烯酰胺凝胶电泳(SDS-PAGE)分离后,GMP被进一步分成高分子量麦谷蛋白亚基(HMW-GS)和低分子量麦谷蛋白亚基(LMW-GS)[8]。与面包强度呈正相关的HMW-GS、LMW-GS和GMP与麦谷蛋白的空间分布模式一致,均在外胚乳层含量最高[4]。
超薄切片染色结合显微镜观察可更为直观地观察灌浆期籽粒的蛋白体分布规律。Savill等[17]运用ArcGIS软件将籽粒横切面由外向内分成5层,并对不同层次的蛋白体面积进行量化,发现面积大于1 μm2的蛋白体在胚乳外层含量较高,而小于1 μm2的蛋白体则多集中在胚乳内部。Tosi等[15]采用蛋白特异抗体标记结合免疫电镜观察灌浆籽粒面筋蛋白空间分布,发现γ醇溶蛋白和HMW亚基在中胚乳中含量较高,而α醇溶蛋白、ω醇溶蛋白和LMW亚基则在外胚乳层中含量最高,与分层磨粉所得的结论基本一致。
1.2 淀粉空间分布规律
淀粉由2种葡萄糖聚合物构成,包括由D-葡萄糖基以α-(1,4)糖苷键连接的线性直链淀粉和聚合度较高且高度分支的支链淀粉。其中直链淀粉约占小麦总淀粉含量的18%~35%[18]。通过分层磨粉对不同层次淀粉及其组分含量测定发现,总淀粉及直链、支链淀粉含量从籽粒外层向内层呈增加趋势,在中胚乳层含量最高,再向内则基本保持不变。支链淀粉和直链淀粉比由麸皮层到外胚乳层迅速下降,而从外胚乳层向内则无显著差异[14]。
成熟期小麦籽粒主要包含A型(>10 μm)和B型(<10 μm)2种淀粉粒[19]。2种类型的淀粉粒在籽粒中分布亦不均匀,亚糊粉层细胞中的B型淀粉粒数较多,而中胚乳和内胚乳中的A型淀粉粒数目较多[20]。Zhou等[14]发现籽粒不同部位淀粉粒的平均粒径亦存在差异,从外层到内层A型淀粉粒的平均粒径呈上升趋势。Xiong等[21]报道,花后15 d的籽粒亚糊粉层的淀粉粒粒径为1.5~25.5 μm,B型和A型淀粉粒的峰值粒径分别为2.5和12.5 μm,淀粉粒多为形状不规则、粒径分布小的复合颗粒;内胚乳层的淀粉粒粒径为1.5~39.5 μm,B型和A型淀粉粒的峰值粒径分别为3.5和18.5 μm,形状多为规则的椭圆形或球形。Tosi等[22]发现花后20 d的籽粒糊粉层淀粉粒数和粒径均显著小于胚乳层,与成熟期淀粉的空间分布规律一致。
1.3 其他组分空间分布规律
细胞壁多糖是谷物产品中膳食纤维的主要来源,小麦籽粒含11.5%~15.5%的总膳食纤维(TDF),其中阿拉伯木聚糖(AX)5.53%~7.42%、纤维素1.67%~3.05%、β-葡聚糖0.51%~0.96%[23]。几乎所有的纤维素及大部分AX和β-葡聚糖均位于籽粒麸皮层,白面粉中仅含有2%~3%的细胞壁多糖。TDF含量由外向内逐层下降,内胚乳中的TDF、AX和β-葡聚糖含量为麸皮层的15.4%、24.2%和31.8%[24]。Velikovi等[25]通过组织切片的酶解试验结合MALDI质谱成像(MSI)鉴定了酶解释放的寡糖,观察不同部位细胞壁多糖分布规律,发现灌浆期AX和β-葡聚糖集中分布在籽粒外层细胞,并随籽粒的成熟逐步向内迁移。
脂肪在籽粒中所占比例较小,在面粉中仅占2.0%~2.5%[26]。大致分为极性脂质(包括细胞膜的结构成分磷脂和半乳糖脂)、三酰基甘油(贮存脂质)和游离脂肪酸3种类型。糊粉层和胚中的总脂质含量最高,而内胚乳中的总脂质含量最低。不饱和脂肪酸的比例表现出相似的趋势[24]。籽粒不同层次的脂质组成同样有差异。游离脂肪酸和三酰基甘油在不同层次面粉的比例均较高,而极性脂质的含量变化较大,面粉中溶血磷脂酰胆碱和二半乳糖基甘油二酸酯的含量较高,而麸皮和糊粉层中卵磷脂和甘油二酯的比例更高[27]。
小麦籽粒中的一些微量物质同样影响人体健康及面粉的加工品质,包括矿质元素、维生素B(主要是B9、B1、B2、B3和B6)和植化素[28]。小麦籽粒中铁和锌的含量由外层向内层逐渐下降,但可溶性铁和锌的比例逐层增加[29]。目前仅知道维生素或植化素在麸皮层和糊粉层的含量较高,其他微量物质由于含量过低,在胚乳中的空间分布规律报道较少[30]。
1.4 加工及烘焙品质空间分布规律
淀粉和蛋白等化学组分在籽粒中的空间分布异质性会导致不同层次面粉加工品质的空间差异。总淀粉和直链淀粉含量及A型淀粉粒比例由胚乳外层向内层上升趋势明显,面粉糊化参数如峰值黏度、低谷黏度、崩解值、最终黏度、回生值和糊化时间等均呈上升的趋势,而糊化温度呈逐渐下降的趋势。贮藏蛋白及其组分含量的空间异质性决定了不同部位面粉面筋的含量和强度显著不同[10]。例如,面粉的碱性水保持力、碳酸钠溶剂保持力、蔗糖溶剂保持力、去离子水溶剂保持力、乳酸溶剂保持力由外胚乳层向内胚乳层呈先上升后下降的趋势;外胚乳层HMW亚基和GMP含量均最高,面粉中的干/湿面筋含量最高,面筋指数较高,形成的面团强度较高[31]。
籽粒不同部位面粉烘焙品质也呈现显著的空间异质性。麸皮层及糊粉层中蛋白质、膳食纤维、钙铁锌等矿质元素含量均较高,但因淀粉含量过低,烘焙产品的感官评价均较差[14]。由麸皮层向内胚乳分层磨制面粉所烘焙出面包的体积、弹性和感官评分先上升后下降,由外胚乳层及中胚乳层磨制面粉制成的面包品质最优,内聚力和回复性数值相对较大,而硬度和咀嚼性数值则相对较小[4]。由外向内,不同层次面粉烘焙饼干的直径先减小后增加,厚度和硬度下降,比容和亮度先上升后下降,感官评分增加;其中,内胚乳层面粉制成的饼干直径、延展因子最大,感官评分最高,厚度和硬度值最小,饼干烘焙品质最好[14]。
2 籽粒品质空间分布差异的形成机制
Shewry等[32]和Evers[33]对软质小麦品种‘Caesar’及硬质小麦品种‘Svenno’和‘Hereward’的籽粒发育进行观察,发现在正常生长条件下,小麦籽粒发育可分为3个阶段:细胞分化和扩张期(花后0~14 d),籽粒不同部位结构基本形成,而蛋白与淀粉的合成处于起步阶段;籽粒充填期(花后14~28 d),面筋蛋白与淀粉的快速合成阶段;脱水期(花后28 d到成熟),籽粒灌浆速率减慢且含水量逐步下降[32-33]。一般在花后8 d左右,淀粉在造粉体中以由蔗糖转化形成的腺苷二磷酸葡萄糖(ADPG)为底物进行生物合成;面筋蛋白则以氨基酸为底物,在花后10 d左右开始在粗面内质网上合成[32]。因此,小麦籽粒品质异质性形成基因可能主要与以下几个过程有关:(1)籽粒由外到内淀粉含量逐层上升,内胚乳中较高的淀粉含量对蛋白产生稀释效应。然而,将小麦籽粒由外向内分成9层(每层约占10%左右),发现内部4层(约占50%)的淀粉含量基本不变,但是蛋白含量仍呈下降趋势[4,10,14],说明这一过程不成立;(2)籽粒不同部位蛋白或淀粉合成的起止时间存在差异。在玉米胚乳的程序性死亡由胚乳中心起始,逐层向外有序进行,因此外胚乳合成贮藏物质的时间更长。但Young等[34]采用伊文思蓝对小麦灌浆期籽粒染色,发现小麦胚乳细胞程序性死亡起始时间呈现无序性,表明籽粒不同部位同一贮藏物质合成时间基本相同,该过程亦不成立;(3)籽粒不同部位淀粉(蔗糖)或蛋白合成底物(氨基酸)的供应存在空间差异;(4)籽粒不同部位淀粉或蛋白合成能力的差异导致其空间分布的异质性。以下主要针对过程(3)和(4)进行综述。
2.1 蛋白质和淀粉合成底物的供应与籽粒品质空间异质性
Ugalde等[35-36]基于微型切片技术分析了花后20 d小麦籽粒游离氨基酸及糖类的组成,发现从胚乳腔运输至籽粒的氨基酸和蔗糖的浓度梯度与蛋白及淀粉的积累梯度不匹配,因此提出籽粒蛋白或淀粉积累异质性与底物的供应没有直接联系。但淀粉合成大致起始于花后8 d,贮藏蛋白合成大致起始于花后 10 d[32],并且淀粉及蛋白的空间异质性从花后14 d才开始形成[15]。因此,需进一步分析不同灌浆期胚乳不同部位游离氨基酸及糖类浓度和组成的变化,尤其是蛋白质及淀粉合成起始的灌浆早期空间分布变化。灌浆早期籽粒较小且含水量较高,获取不同部位样本的难度大,相关研究进展缓慢。Peukert等[37]通过质谱成像技术观察花后10~20 d的大麦胚乳,发现近胚乳腔处的二糖、三糖及己糖含量均高于外胚乳及糊粉层,与成熟籽粒淀粉空间分布一致。Zhong等[38]发现灌浆前期(花后7~11 d)游离氨基酸在籽粒中的分布模式与成熟期籽粒蛋白质分布趋势一致。因此,淀粉和蛋白质分布的空间异质性可能与灌浆早期底物的供应能力密切相关;但随着灌浆中后期淀粉和蛋白质迅速合成,底物消耗很快,尚缺乏直接的证据证明整个灌浆期间底物的供应能力导致了蛋白质和淀粉含量空间分布的异质性。而关于脂质、膳食纤维、微量元素等物质的空间分布差异与其合成底物的关系尚无报道。
灌浆底物供应能力在籽粒中的空间差异与灌浆物质的运输路径息息相关。王忠等[39]曾提出假说:通过小穗维管束运输来的灌浆物质先卸载到质外体(胚乳腔)后再运输给胚乳,这个过程可能存在2条途径:一条是传递细胞路径,通过胚乳腔周边的传递细胞直接进入内胚乳细胞并向外呈辐射状运输(图2-A);另外一条途径是糊粉层路径,通过种皮质外体运输到籽粒外周,然后穿过糊粉层由外向内运输至胚乳各层(图2-B)。目前,有关这2条运输路径的报道尚存在较大争议。Chen等[40]利用树脂半薄切片结合Image-Pro Plus软件观察了花后18和28 d籽粒中蛋白体的分布差异,发现蛋白体数量和相对面积从大到小依次为腹部近糊粉层、背部近糊粉层、腹部中胚乳层。这可能与蛋白质合成底物运输路径有关:氨基酸由胚乳腔经由籽粒外周,穿过外侧糊粉层运输,由于背部位于腹部的下游,从胚乳腔运输到背部的距离不仅比运到腹部长且需消耗更多的能量,进而推测氨基酸主要通过糊粉层路径进行运输。Xiong等[21]观察花后15 d小麦胚乳淀粉粒积累动态,推断出蔗糖也先在外胚乳积累,进而运输到内胚乳,最后运输至转移细胞,认为淀粉合成底物与蛋白质合成底物运输均通过糊粉层路径。然而,Peukert等[37]却发现在花后 10~20 d大麦胚乳腔周边细胞与糖类代谢相关基因的表达与代谢活性更强,并推断蔗糖主要通过转移细胞途径进行传递。与其结论一致的是,Moore等[41]用毛细管将15N标记的谷氨酰胺注入灌浆期麦穗中,采用纳米二次离子质谱技术(NanoSIMS)测定蛋白体中15N的富集程度,发现15N标记的谷氨酰胺从胚乳腔直接经由转移细胞运输至胚乳中,最后运输到亚糊粉层细胞。可见,灌浆底物运输的模式仍存在明显的争议,并且已有研究均通过观测糖类或蛋白体的空间分布情况来间接推测底物的转运路径,尚缺乏灌浆底物输入胚乳路径的直接证据。

图2 灌浆底物运输路径模式图(改编自王忠等[39])Fig.2 Proposed transport pathway of substrate for starch synthesis in endosperm(modified from Wang et al[39])A. 传递细胞路径,灌浆底物通过胚乳腔周边的传递细胞直接进入内胚乳细胞;B. 糊粉层路径,灌浆底物通过种皮质外体运输到籽粒外周,穿过糊粉层由外向内运输至胚乳。A. Transfer cell pathway,filling substrate transports into inner endosperm via radially across the tissue from the cavity and transfer cells;B. Aleurone pathway,filling substrate transports into inner endosperm via aleurone around the endosperm.
2.2 蛋白质及淀粉合成能力与籽粒品质空间异质性
籽粒淀粉合成主要在造粉体中进行,以蔗糖为起点,催化形成淀粉合成的直接前体ADPG,然后经一系列淀粉合成酶催化α-1,4糖苷键的延伸[42]。醇溶蛋白和麦谷蛋白为贮藏蛋白,在小麦胚乳蛋白体中积累,胚乳贮藏蛋白的合成、折叠、沉积发生在内膜系统[22]。蛋白体形成于小麦胚乳发育早期,以直径 2 mm 左右的单个小球形式出现,在胚乳发育中期扩大、聚集和融合,由球形转变为不规则的形状,后期单个蛋白体转化为蛋白质基质填充在淀粉颗粒间[43]。淀粉和蛋白质的合成均由多个基因共同参与调控。编码贮藏蛋白的基因主要是在转录水平上被调控的[44],因此母体组织可能存在特定的调节信号调控胚乳不同部位蛋白质合成编码基因的转录速率[15]。由于内胚乳距离母体组织更近,所以相较于糊粉层更易受到干扰,蛋白质合成编码基因的表达水平相对低。Ma等[45]将花后25 d的籽粒粗略分为蛋白质含量较低的内胚乳部位及蛋白质含量较高的其余胚乳部位,发现参与碳氮代谢的差异基因主要与氨基酸转换及氨基酸运输相关,直接编码蛋白合成的基因表达在两部位间不存在差异,表明蛋白合成能力可能与蛋白质含量的空间分布模式不相符。此外,Gillies等[46]对花后6、9、14 d籽粒的转录组数据进行综合分析,发现大多数参与淀粉合成和降解的基因在胚乳中表达上调,证明淀粉含量空间分布模式可能与编码淀粉合成相关基因的表达水平有关。但上述研究仅将灌浆期籽粒分成2个部分,很难实现不同层次间的精准比较。
近年来,激光捕获显微切割技术的发展为实现小麦籽粒不同部位微量样品的精准获取提供了可能。前述的分层磨粉技术可以将籽粒分为麸皮、糊粉层、外胚乳、中胚乳和内胚乳等不同层次面粉,但该技术仅能用于成熟籽粒的分层分析,无法获得灌浆期间新鲜籽粒的分层样品。激光捕获显微切割技术可通过显微镜观察并定位目标细胞,然后通过与显微镜相连的计算机显示屏对目标区域细胞划线标记,再经激光准确切割划线区域来提取目标细胞。激光显微切割技术可从新鲜样品组织中分离出纯细胞群甚至单一细胞,成功解决了细胞异质性问题[47]。如同步结合冷冻切片技术,还可提取高质量的RNA,用于转录组学等方面的研究。Zhan等[48]采用该技术,从分化中的玉米胚乳分离收集到5种类型的细胞进行转录组学分析,鉴定出母本与子代的差异表达基因,成功建立了不同类型胚乳细胞基因共表达模型。Zhong等[38]采用这一技术成功获取了不同灌浆时期小麦籽粒糊粉层、外胚乳、中胚乳和内胚乳细胞,以及转移细胞,并进行特定基因的表达水平分析,发现高分子量麦谷蛋白亚基(HMW-GS)及醇溶蛋白编码基因在中胚乳和内胚乳的表达水平较高,与成熟期籽粒蛋白空间分布模式不相符,因此确定蛋白质分布的空间差异与蛋白质合成能力无关。但由于蛋白质及淀粉合成过程涉及的基因数较多,有必要获取不同灌浆时期籽粒不同部位细胞时空差异基因,系统全面解析蛋白质和淀粉合成能力与籽粒品质空间异质性的关系。
3 栽培措施及环境对籽粒品质空间分布的调控效应
3.1 籽粒不同部位蛋白质和淀粉的积累对栽培措施的响应存在差异
肥料运筹会显著影响小麦籽粒品质的空间分布模式(表1)。He等[49]发现贫硫的ω-醇溶蛋白含量对氮肥增加的响应最为显著,在整个籽粒中同步增加;HMW-GS含量随施氮量增加而增加,但外胚乳层比内胚乳层增加效应更为明显,而富硫的醇溶蛋白则表现出几乎相反的趋势。氮素对淀粉品质的影响主要表现在中胚乳和内胚乳直链淀粉含量、直链淀粉含量/支链淀粉含量比、淀粉粒的分布及糊化性质的变化[10],且不同筋力品种对氮素的响应不同。例如,施氮对弱筋小麦品种‘扬麦15’籽粒不同部位的总淀粉、支链淀粉含量的调控能力均较弱,而对中筋品种‘扬麦16’和强筋品种‘徐麦32’籽粒的糊粉层和外胚乳层总淀粉含量及支链淀粉含量影响则较大[10]。追氮时期对籽粒品质空间分布同样具有调控效应,追氮时期由拔节期后移至孕穗期对各层面粉面筋蛋白含量均有提高作用,其中糊粉层和外胚乳层的增加最为显著,且对麦GMP和麦谷蛋白亚基的调控作用与面筋蛋白表现一致,而对面筋指数调控效果不显著;追氮时期前移至返青期则降低总蛋白及各蛋白组分含量,且外层的响应更为明显[4]。进而直接导致烘焙品质的空间分布变化:追氮后移提升了糊粉层与外胚乳层面粉烘焙面包的体积及感官评价,而对其余层次的烘焙品质调控不明显;追氮前移对面包烘焙品质(体积、外观及感官评价)的调控作用表现出相反趋势。此外,施用硫肥可增加籽粒每层面粉总蛋白及麦谷蛋白含量,对外胚乳和中胚乳中HMW-GS和外胚乳中醇溶蛋白含量的增加效果优于其余部位[50]。

表1 栽培措施对籽粒不同部位蛋白含量的调控效应Table 1 Regulation effects of cultivation practices on protein gradients in wheat grain
在生产上采取增密减氮栽培措施可实现对弱筋小麦籽粒品质的空间分布调控,增密减氮主要通过调控胚乳中蛋白体和面筋蛋白的空间分布以降低面粉沉降值,改善弱筋小麦的加工品质。籽粒蛋白品质对增密减氮的响应从大到小依次为外胚乳、中胚乳、内胚乳,因此,蛋白在外层的积累相对更多,可通过磨粉有效去除,降低面粉蛋白含量超标风险;同时,密度增加弥补了减氮带来的产量下降问题,故可以通过增密减氮的优化实现弱筋小麦高产优质同步[51]。
叶面喷施氨基酸同样可以对籽粒品质空间分布起到调控作用。李叶[52]在开花期叶面喷施氨基酸与尿素,发现2个中筋小麦品种不同层次A型淀粉粒数目、平均淀粉粒粒径,总蛋白、醇溶蛋白、麦谷蛋白与麦谷蛋白总亚基、氨基酸与面筋含量及沉降值都表现出不同程度的提高。2个品种籽粒蛋白及其组分和亚基含量的增幅由外至内呈先增加后降低的趋势,以中胚乳的增幅最大;而外胚乳层麦谷蛋白大聚合体与氨基酸含量的增幅大于内胚乳。
就微量元素而言,Shi等[53]发现施氮可以增加麸皮层Fe和Zn的含量,但面粉中仅有Zn含量增加。Xue等[54]发现全麦粉和麸皮中Fe含量均增加,而在面粉中甚至表现出下降的趋势,表明通过施氮很难实现面粉中Fe含量的增加。董明[55]研究发现叶面喷施锌肥可有效提升籽粒和面粉中Zn含量:花后5、15、25 d喷施锌肥均可增加籽粒不同层次Zn含量,且籽粒外层的增加量更多;但喷施锌肥会降低小麦籽粒各层面粉Fe和Mn的含量,籽粒外层矿质元素受影响较大,而内层矿质元素受影响较小。
3.2 环境对籽粒品质空间分布的调控效应
环境因素会对籽粒品质造成较大影响[56]。灌浆期高温提高了小麦籽粒蛋白含量[57],籽粒蛋白梯度差异会增大,即外胚乳层会积累更多的蛋白质。由于附着在糊粉层的外胚乳细胞在磨粉过程中很容易被去除,最终所得面粉的蛋白含量并不会改变,且蛋白含量会下降[17]。高温干旱会降低花后30 d籽粒内胚乳细胞壁中阿拉伯木聚糖的糖基化程度,而对籽粒外层细胞无显著影响[58]。有关于低温、渍水等逆境对籽粒品质空间分布的影响尚缺少报道,不同区域条件下小麦籽粒品质的空间异质性是否一致尚不明确。
4 籽粒品质空间分布异质性在面粉加工中的应用
籽粒品质空间分布的异质性为通过改进磨粉工艺来生产不同品质专用类型的面粉提供了可能。如表2 所示:中外胚乳位置的面粉HMW-GS含量及谷醇比较高,面团强度弹性较高且黏度较低,适于要求高面筋强度的面包烘焙[59]。相反,内胚乳层淀粉含量较高,谷醇比低,面团延展性好,适合烘焙饼干等产品。Shewry等[60]指出外胚乳面粉有足够的弹性和强度,可用于意大利面的制作;直链和支链淀粉含量比值的差异结合蛋白组分含量的差异使不同部位面粉在速冻食品(如馒头、面包、水饺等)制作中也有潜在的应用。

表2 小麦籽粒品质空间分布规律及加工应用前景Table 2 Spatial heterogeneity distribution of wheat grain quality and implications for processing within the wheat grain

续表2 Table 2 continued
因此,尽管从整个籽粒角度来看,其蛋白质或面筋含量并不符合传统的“强筋”或者“弱筋”的定义,但特定部位的面粉也可以满足最终烘焙产品的需求。富含膳食纤维、葡聚糖和微量元素等对人体健康有益成分的糊粉层,亦值得在健康面制品开发中予以重视。
基于不同层次面粉品质的差异性,还可通过不同层次面粉的混配生产特定麦制品加工所需的优质专用面粉。已有人以中胚乳层的面粉为基础,加入蛋白质和矿质元素含量较高的糊粉层面粉,所混配的面粉糊化参数下降,矿质元素含量上升,烘焙的面包色泽加深,体积、硬度、咀嚼性提高,但添加量超过20%时,面包品质和感官得分会出现明显的下降;以内胚乳面粉为基础添加麸皮层面粉,混配面粉黏度值下降,矿质元素上升,烘焙的饼干颜色加深,直径、延展因子和感官得分呈下降趋势;而将小麦籽粒第2层面粉以20%的添加比例与第5层面粉混配,或第1层以10%的添加比例与第9层混配,最大程度提升饼干的营养品质,并保证较好的口感[61]。
5 研究展望
综上所述,小麦籽粒品质空间分布的异质性已较为明确,但受分样技术等原因影响,这种空间分布异质性形成的机制研究缓慢。近年来,激光显微切割技术和单细胞测序技术的发展为弥补这一空白提供有力的技术支撑。未来可应用上述技术获取籽粒任意部位的细胞,结合多组学手段,明确籽粒品质空间异质性形成的关键调控过程、关键调控基因或基因网络,阐明其作用机制。Tauris等[62]用激光显微切割技术获取大麦籽粒糊粉层、珠心突起、胚和胚乳细胞,结合芯片技术明确锌在籽粒中的转运路线。随后,该课题组在胚乳细胞特异过表达1个锌转运蛋白基因(HvMTP1),成功提高了胚乳中锌的含量,促进锌从糊粉层向胚乳的运输和转移[63]。这为小麦籽粒品质空间分布的主动调控提供了借鉴,表明籽粒品质空间分布或许可通过操纵关键基因的定向表达进行改良。
周龙静[64]在田间条件下研究了叶面喷施氮肥对180个小麦重组自交系家系籽粒蛋白含量的空间分布,发现叶面喷施氮肥后小麦籽粒不同部位的总蛋白含量均有不同程度的提高,但不同家系不同层次面粉对喷施氮肥的响应敏感性不同,主要分为3种类型(图3):外层面粉响应更敏感(图3-A);内层面粉响应更敏感(图3-C);对氮素的响应由外向内呈先增强后减弱的趋势(图3-B)。这表明籽粒品质空间分布对氮肥的响应因基因型而异,而目前关于相关基因的探究较为匮乏。如果能明确调控对氮肥差异响应的关键基因,可以根据不同品种对施氮响应的敏感性差异,采用有针对性的配套适宜栽培技术,满足不同品质专用类型小麦的需求。

图3 不同品系蛋白含量空间分布对叶面施氮的差异响应(改编自周龙静[64])Fig.3 Different response of protein spatial distribution to foliar spraying nitrogen in different wheat lines(modified from Zhou[64])A. 品系197;B. 品系278;C. 品系250。P1—P5分别代表麸皮层、糊粉层、外胚乳、中胚乳和内胚乳。A. Line 193;B. Line 278;C. Line 250. P1-P5 represent bran,aleurone,outer endosperm,middle endosperm and inner endosperm,respectively.
猜你喜欢
农业技术与装备(2022年6期)2022-08-17
中国农学通报(2022年13期)2022-05-31
园艺与种苗(2021年9期)2021-11-25
辽宁农业科学(2021年1期)2021-03-17
祝您健康·文摘版(2021年3期)2021-03-09
小学生学习指导(高年级)(2019年5期)2019-11-27
金桥(2018年9期)2018-09-25
科学种养(2017年6期)2017-06-13
美食堂(2016年5期)2016-05-23
食品工业科技(2014年23期)2014-03-11
