
山黧豆β-腈基丙氨酸合成酶基因LsCAS的原核表达及蛋白聚合状态分析
2021-11-09 01:14:10贾海燕李辰浩宋瑶瑶刘凤娟焦成瑾徐全乐
西北植物学报 2021年9期
贾海燕,李辰浩,宋瑶瑶,刘凤娟,焦成瑾,徐全乐
(1 陇东学院 生命科学与技术学院,甘肃庆阳 745000;2 西北农林科技大学 生命科学学院,陕西杨陵 712100; 3 天水师范学院 生物工程与技术学院,甘肃天水 741000)
β-N-草酰-L-α,β-二氨基丙酸(β-N-oxalyl-L-α,β-diaminopropionic acid,β-ODAP)在山黧豆(Lathyrussativus)中首次获得分离和鉴定[1],并被发现广泛存在于三七(Panaxnotoginseng)、西洋参(Panaxquinquefolium)和人参(Panaxginseng)等物种[2-3]。研究表明,β-ODAP是传统中药材三七止血的活性成分,被命名为三七素(dencichine)[2],并在神经保护、糖尿病及肾病防治等过程中发挥重要作用[3-8]。因而,β-ODAP生物合成的调控是山黧豆、三七等作物品质形成机理研究的重要内容。
β-ODAP的生物合成在山黧豆种子萌发期和成熟期形成2个显著高峰[9]。对萌发期种子的代谢组学和转录组学分析表明,β-ODAP的生物合成与硫代谢等基础代谢途径密切关联[10-12]。其中,β-腈基丙氨酸合成酶(β-cyanoalanine synthase,β-CAS)负责催化异噁唑-5-酮与半胱氨酸(Cys)反应生成β-异噁唑-5-酮-丙氨酸[β-(isoxazolin-5-on-2-yl)alanine,BIA]。BIA进一步通过两步反应生成β-ODAP[13-14]。
在拟南芥和大豆等物种中的研究表明,β-CAS隶属于磷酸吡哆醛(pyridoxal phosphate,PLP)依赖的β基取代的丙氨酸合成酶家族(β-substituted alanine synthase,Bsas),可形成二聚体,分子量范围为50~70 kD[15-16]。在山黧豆中,将LsCAS的PLP附着关键位点Lys突变后导致突变体酶活丧失[14,17],说明LsCAS也是PLP依赖性蛋白酶。该蛋白可通过与丝氨酸乙酰基转移酶(serine acetyltransferase,SAT)互作起到β-ODAP生物合成调控分子开关的作用[14]。
植物硫代谢途径调控的一个典型特征是2个CS同源二聚体与2个SAT同源三聚体共同形成半胱氨酸调控复合物十聚体(cysteine regulatory complex,CRC),并通过聚合/解聚状态调节CS 与SAT酶活[18-20]。为进一步研究LsCAS功能及聚合状态,本研究克隆了LsCAS基因的CDS序列,构建了pGEX2T-LsCAS原核表达载体,通过蛋白纯化和Western-blot分析进行了LsCAS功能验证,并利用分子排阻初步解析山黧豆LsCAS蛋白的聚合状态,深入了解LsCAS的功能。
1 材料与方法
1.1 试验材料
试验所用山黧豆(Lathyrussativus)种子为西北农林科技大学生命学院实验室保存。种子在含腐殖土、蛭石、珍珠岩(1∶1∶1)的混和基质中于23 ℃萌发6 d,取其根部提取RNA。GST亲和柱购自碧云天生物技术公司,GST标签抗体购自美国Affinity公司,大豆CS 抗体为美国农业部Hari Krishnan教授惠赠,牛凝血酶购自上海翊圣生物科技有限公司,10 kD超滤管等购自Millipore。
1.2 试验方法
1.2.1LsCAS基因克隆采用Invitrogen公司的总 RNA 提取试剂盒(Trizol法)提取山黧豆6 d龄幼苗的根部RNA,利用TaKaRa公司的反转录试剂盒(PrimeScriptTMⅡ1st Strand cDNA Synthesis Kit)进行反转录合成cDNA。采用50 μL反应体系进行PCR扩增,其中5×PrimeSTAR 缓冲液 10 μL,dNTP 混合液(2.5 mmol/L)4 μL,模板2 μL,PrimeSTAR HS DNA 聚合酶(2.5 U/μL)0.5 μL;上下游引物(10 μmol/L)各1.5 μL。PCR程序为:98 ℃ 1 min;98 ℃ 10 s,57 ℃ 5 s,72 ℃ 1 min 15 s,30个循环;最后72 ℃ 延伸5 min。引物序列为F(ATCACCTTCCTCAAGCGAACCTC)和R(CTC-GGAACTCGTGGAAGAAAT)。PCR产物经1%琼脂糖凝胶电泳检测后,送北京擎科生物科技有限公司测序。
1.2.2 原核表达载体构建利用EcoR I和BamH Ⅰ对pGEX2T载体进行双酶切,经1%琼脂糖凝胶电泳检测后采用天根DNA纯化试剂盒(Tiangen)进行切胶回收,利用T4DNA连接酶(TaKaRa)和带有EcoR Ⅰ和BamH Ⅰ酶切位点的LsCAS片段连接。重组产物转化到DH5α后,经菌落PCR、双酶切和测序鉴定。
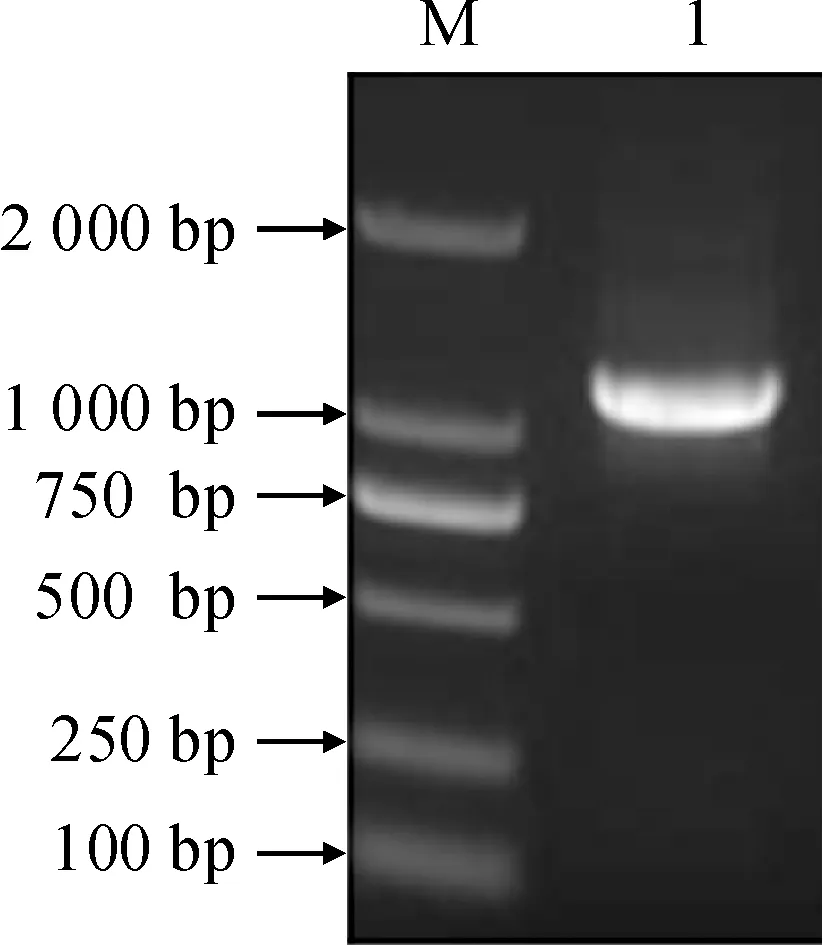
M. DL2000;1. RT-PCR图1 LsCAS基因的RT-PCR扩增结果Fig.1 RT-PCR amplification result of LsCAS
1.2.3 LsCAS蛋白纯化将pGEX2T-LsCAS重组质粒利用KCM法转化入感受态BL21(DE3)中。挑取单克隆于5 mL含Amp的LB液体培养基中,37 ℃、180 r/min培养12 h后;取2 mL菌液转接于200 mL含有Amp的LB液体培养基中培养至OD600为0.4~0.6,加入终浓度为0.2 mmol/L IPTG,25 ℃培养10~12 h。收集菌体后用PBS缓冲液重悬,超声破碎后收集上清液。使用碧云天GST亲和柱(BeyoGoldTMGST-tag Purification Resin)进行亲和层析,用PBS缓冲液平衡GST亲和柱并洗去未结合杂蛋白,用含有10 mmol/L GSH的GST洗脱缓冲液洗脱目的蛋白。通过SDS-PAGE对纯化蛋白进行鉴定。
1.2.4 Western-blot分析上述纯化蛋白使用12.5%的SDS-PAGE分离后,转移到NC 膜上,用GST标签抗体和大豆CS 抗体分别进行验证。
1.2.5 分子排阻将纯化的LsCAS用牛凝血酶切割GST标签,用10 kD超滤管浓缩至蛋白终浓度为3 mg/mL。取1 mL样品,使用AKTA蛋白纯化系统,Superdex 200 Increase 10/300 GL凝胶柱分析纯化的LsCAS分子量。检测条件为:柱温室温,流速0.5 mL/min,流动相0.1 mol/L Tris-HCl缓冲液(pH 7.5,含200 mmol/L NaCl),检测波长为280 nm和412 nm。
2 结果与分析
2.1 山黧豆LsCAS基因CDS序列克隆
利用RT-PCR从山黧豆萌发6 d的幼苗根部RNA中进行LsCAS基因CDS序列的特异性扩增。结果表明,PCR扩增产物在1 000 bp左右,与预期相符,且条带单一(图1)。PCR产物的测序结果与转录组数据序列完全一致,表明所获序列为β-腈基丙氨酸合成酶基因。LsCAS基因的CDS序列为1 035 bp,编码344个氨基酸。编码的蛋白质具有典型的CBS-like 蛋白功能结构域(Cystathionine beta-synthase,CBS)和半胱氨酸合成酶(CS),表明LsCAS基因克隆成功。
2.2 LsCAS表达载体构建
采用酶切连接法构建LsCAS基因的原核表达载体。经菌液PCR验证后(图2,A),挑取阳性克隆进行质粒提取及EcoR Ⅰ和BamH Ⅰ双酶切验证(图2,B)。琼脂糖凝胶电泳检测结果表明,重组质粒的电泳迁移率略小于pGEX2T空载体;重组质粒经双酶切后在5 kb和1 kb左右呈2条条带,其电泳迁移率分别与pGEX2T空载体和LsCAS基因扩增产物大小相当。这表明重组载体构建成功。经测序证实后,将其命名为pGEX2T-LsCAS。
2.3 LsCAS蛋白纯化
利用IPTG进行重组质粒诱导表达后,采用GST亲和层析法对LsCAS融合蛋白进行纯化。SDS-PAGE检测表明,所获融合蛋白条带单一,大小在64 kD左右,符合预期大小。进一步利用GST标签抗体和大豆CS抗体进行Western-blot分析,在诱导后的菌体蛋白和纯化后的重组蛋白中均能检测到特征条带,说明所获融合蛋白为山黧豆LsCAS蛋白(图3)。

A. 菌液PCR验证:1-8. 菌落PCR; B. BamH Ⅰ和EcoR Ⅰ双酶切验证:1. pGEX2T;2. pGEX2T-CAS;3. pGEX2T-CAS酶切产物;4. pGEX2T酶切产物;5. LsCAS基因PCR产物;M. DNA marker图2 重组质粒验证A. Colony PCR: 1-8. Colony PCR of randomly selected colonies; B. Double enzyme digestion by BamH Ⅰ and EcoR Ⅰ: 1. Plasmid pGEX2T; 2. Plasmid pGEX2T-LsCAS; 3. Enzyme digested fragments of pGEX2T-LsCAS; 4. Enzyme digested fragments of pGEX2T; 5. Cloned LsCAS gene via RT-PCR; M. DNA markerFig.2 Identification of recombinant plasmid
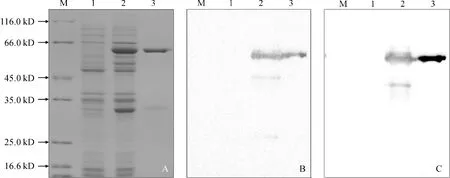
A. SDS-PAGE;B. 利用大豆CS抗体检测;C. 利用GST标签抗体进行检测;M.蛋白质分子量标准;1.IPTG诱导前总蛋白质;2.IPTG诱导后总蛋白质;3.纯化后的LsCAS蛋白图3 LsCAS蛋白纯化的 SDS-PAGE及Western-blot检测A. SDA-PAGE; B. CS antibody; C. GST antibody; M. Regular range protein marker; 1. Total protein before IPTG induction; 2. Total protein after IPTG induction; 3. The purified LsCASFig.3 SDA-PAGE and Western-blot analysis of purified LsCAS
2.4 分子排阻分析
使用Superdex 200 Increase 10/300 GL凝胶柱判断LsCAS在离体条件下的分子量。结果表明(图4),纯化的LsCAS在412 nm下具有特征吸收峰。由于412 nm是磷酸吡哆醛PLP的特征吸收波长[21],说明LsCAS属于PLP依赖的蛋白酶家族。在280 nm检测波长下,LsCAS在洗脱体积为14.56 mL处有最大吸收值。根据标准蛋白分子的排阻体积进行计算,对应的分子量为145 kD左右。由于LsCAS蛋白单体分子量为36 kD,说明LsCAS在离体条件下可能以四聚体方式存在。

图4 分子排阻验证LsCAS分子量及聚合方式Fig.4 Molecular weight analysis of LsCAS via size-exclusion chromatography
3 讨 论
在山黧豆中,LsCAS是β-ODAP生物合成途径的关键酶,负责催化异噁唑-5-酮与Cys合成关键中间化合物BIA[13-14]。研究表明,β-CAS隶属于磷酸吡哆醛PLP依赖的半胱氨酸合成酶CS家族[16,21]。因而,PLP附着的关键位点Lys(K)对CS功能的行使至关重要[21]。例如,拟南芥AtOASSK46A突变体表现为Cys合成酶活性丧失[22]。在山黧豆中,将PLP附着关键位点的Lys突变后,进行E.coliNK3功能互补分析和酶活检测均表明,LsCS及LsCAS突变体酶活丧失[14, 17]。本研究在分子排阻试验中,检测到LsCAS蛋白在412 nm波长下具有PLP的特征吸收峰,进一步证实了上述结论。
拟南芥、大豆等作物中的研究表明,植物硫代谢途径调控的一个典型特征是2个CS同源二聚体与2个SAT同源三聚体共同形成半胱氨酸调控复合物CRC,并通过复合物的聚合/解聚状态调节CS与SAT酶活[18-20]。当形成CRC复合物时,会降低CS的活性而升高SAT活性[23]。山黧豆LsCAS在发挥功能时,可通过与LsSAT2互作起到β-ODAP生物合成调控分子开关的作用[14],这暗示LsCAS与LsSAT2可能通过CRC复合物的形成调控二者酶活。将LsCAS与LsSAT2采用梯度摩尔比例混合后,在降低LsCAS酶活的同时确实会增加LsSAT2活性[14]。本研究利用分子排阻法证实,LsCAS可能以四聚体方式存在,说明LsCAS可能以两个二聚体方式参与CRC复合物的形成。
CRC的聚合与解聚状态由细胞内的硫含量决定[24]。在硫充足情况下,山黧豆LsCAS 和LsSAT2形成CRC调控复合物,增强了SAT活性;充足的硫供应保证了Cys的合成,进而在游离LsCAS作用下合成BIA及β-ODAP。在低硫情况下,OAS的积累促使CRC解聚,降低了SAT活性;低水平的Cys限制了以其为底物的BIA及β-ODAP合成。但积累的OAS可在CS催化下和异噁唑-5-酮以较低效率合成BIA[14]。
综上所述,本研究克隆了山黧豆LsCAS基因的CDS序列,通过原核表达获得了山黧豆LsCAS蛋白并利用Western-blot进行了验证。在此基础上,利用分子排阻试验证实了LsCAS为PLP依赖性蛋白酶,可能以四聚体方式发挥作用。在拟南芥中,AtCYS-C1编码的AtCAS蛋白定位于线粒体,主要参与氰基代谢和根毛发育[25-26]。那么,山黧豆LsCAS是否和拟南芥AtCAS具有相同的作用方式、其功能是否存在差异,有待进一步研究。
猜你喜欢
原子与分子物理学报(2021年2期)2021-03-29 07:30:46
中国科技纵横(2021年24期)2021-03-02 06:42:52
中成药(2018年7期)2018-08-04 06:04:18
中成药(2018年6期)2018-07-11 03:01:12
食品科学(2018年10期)2018-05-23 01:27:28
中成药(2018年3期)2018-05-07 13:34:18
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
西南军医(2015年6期)2015-01-23 01:25:50
山西大同大学学报(自然科学版)(2013年5期)2013-09-13 10:44:14
