
RANKL基因在老年小鼠卵巢颗粒细胞中的作用特征
2021-11-04 08:17郭路刘晓程顾超李斌
老年医学与保健 2021年5期
郭路,刘晓程,顾超,李斌
复旦大学附属妇产科医院妇科/上海市女性生殖内分泌相关疾病重点实验室,上海200011
随着女性生育时间的延迟、三孩政策的开放以及人口老龄化的加剧,卵巢功能保护越来越受到社会关注。始基卵泡消耗加速、卵泡闭锁增多,导致卵巢储备功能耗竭,是卵巢衰老的病理生理特征,并伴随相应的围绝经症状、组织器官退化和代谢功能紊乱,严重危害女性身心健康[1]。RANKL 是肿瘤坏死因子超家族成员,参与多种生理病理过程,如骨重塑、淋巴结形成、胸腺发育、孕期乳房发育等[2]。RANKL参与调控破骨细胞分化和骨代谢过程,在老年女性骨质疏松发生中起着重要作用,并且氧化应激可能是其潜在作用机制[3-4]。近年来研究发现氧化应激相关基因在始基卵泡形成、卵泡发育、成熟、闭锁等多个环节中均发挥作用,可能对卵巢储备功能维持和卵巢衰老发展具有重要影响[5]。研究还发现RANKL 与月经初潮和更年期发生有关[6]。RANKL-/-小鼠生育能力降低,但RANKL 基因对卵巢功能的作用目前研究较少[7-8]。因此,本研究旨在通过RANKL-/-老年小鼠评估RANKL基因对卵巢功能影响及其可能作用机制,探索卵巢功能保护靶点,以期改善女性生活质量。
1 资料与方法
1.1 实验分组与取样 30 周龄SPF RANKL-/-雌鼠及同周龄野生型C57 雌鼠均购于上海南方模式生物科技股份有限公司,每组5 只,在标准的实验室条件下适应性喂养7 d。每天早9 时行阴道脱落细胞涂片检查,观察小鼠动情周期[9]。2 组鼠于喂养后1月处死,收集双侧卵巢及子宫,称定质量并计算子宫指数及卵巢指数。子宫指数=子宫湿重(mg) /体重(g)×100%;卵巢指数=卵巢湿重(mg) /体质量(g) ×100%[10]。10%甲醛溶液固定卵巢24 h,石蜡包埋,常规切片,HE染色后400 倍光镜下观察,分别对始基、初级、次级及闭锁卵泡进行计数[11]。
1.2 酶联免疫吸附测定(enzyme linked immunosorbent assay,ELISA)全血收集后4 ℃离心10 min,收集上清后,按照相应说明书测定血清抗苗勒管激素(anti-Mullerian hormone,AMH)、雌二醇(Estrogen,E2)浓度、超氧化物歧化酶(superoxide dismutase,SOD)、谷胱甘肽过氧化物酶(glutathione peroxidase,GSHPx)和丙二醛(malondialdehyde,MDA) 活性。
1.3 Ki-67 免疫组化 石蜡切片预热,梯度浓度乙醇脱蜡。95 ℃抗原修复30 min,3%双氧水避光孵育15 min,PBS 洗3 次,室温下封闭30 min,50 L Ki-67抗体(1:1 000,Abcam)4 ℃过夜。第2 天,50 L二抗孵育60 min,加入50 L DAB,显微镜下观察。Image Pro Plus 6.0 软件分析图像灰度积分光密度(IOD),用单位面积光密度(平均IOD)进行统计分析。
1.4 TUNEL 染色 二甲苯脱蜡10 min,2 次,梯度浓度乙醇脱蜡5 min,20 g/mL 蛋白酶K 作用30 min。PBS 洗涤3 次。加入TUNEL 溶液,在共聚焦显微镜下观察。
1.5 统计学分析 采用SPSS 21.0 软件对数据进行分析。数据符合正态分布且方差齐的计量资料以均数±标准差(±s)表示,用t检验;若方差不齐,用非参数检验。<0.05 表示差异有统计学意义。
2 结果
2.1 RANKL 基因对颗粒细胞增殖及凋亡作用 颗粒细胞为卵巢功能细胞,免疫组化检测显示RANKL-/-组小鼠卵巢组织颗粒细胞内增值相关标记因子Ki-67 表达显著降低,且TUNEL 染色提示RANKL-/-小鼠颗粒细胞凋亡显著增加。见图1~2。
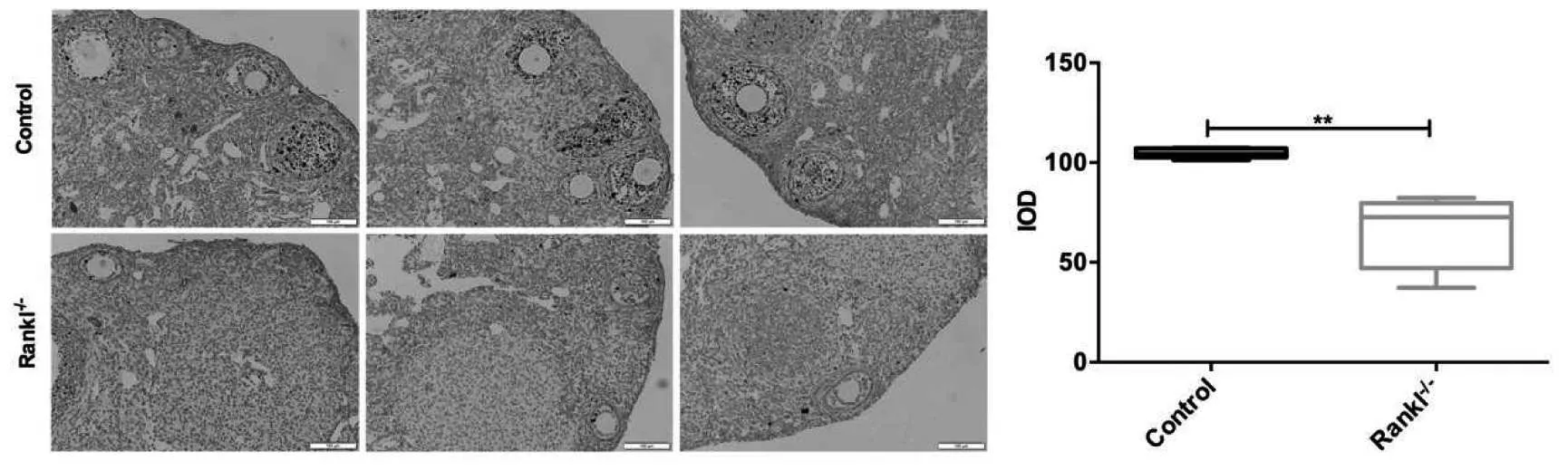
图1 RANKL-/-小鼠卵巢组织颗粒细胞Ki-67 表达
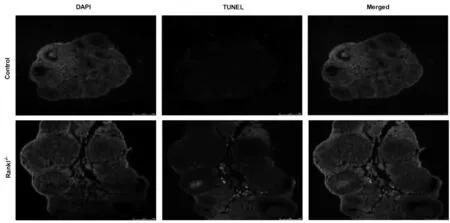
图2 RANKL-/-小鼠卵巢组织颗粒细胞凋亡情况
2.2 RANKL 基因对小鼠卵巢功能作用RANKL-/-小鼠体质量降低、动情频率及始基卵泡数量减少、闭锁卵泡数量增加,血清AMH 水平降低,但与对照组相比差异无统计学意义。RANKL 基因对卵巢指数无明显影响,但子宫指数显著升高。此外,实验发现RANKL-/-小鼠血清E2 水平显着降低。以上结果表明RANKL基因敲除影响卵巢颗粒细胞数量及E2合成能力,但其对卵巢储备功能作用不明显,其影响机制仍需后续进一步研究。见图3。

图3 RANKL 基因对卵巢功能作用
2.3 氧化应激可能参与RANKL 基因作用颗粒细胞过程 与对照组相比,RANKL-/-小鼠血清抗氧化相关因子SOD 抑制率和GSH-Px 活性显著降低,氧化相关因子MDA 活性显着升高。见图4。
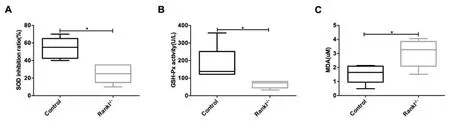
图4 RANKL-/-小鼠氧化应激水平改变
3 讨论
RANKL基因主要表达于淋巴结、胸腺、T 细胞、树突状细胞以及卵巢等组织中,是骨保护蛋白的配体,是破骨细胞分化和活化的关键因子。T 细胞活化可诱导该基因的表达,导致破骨细胞生成和骨丢失的加剧,进而引发骨质疏松以及其他骨疾病[12]。研究发现RANKL基因在乳腺发育、调节激素波动和卵巢周期中起着重要作用[13]。然而,RANKL 基因对卵巢功能作用目前仍待研究,氧化应激是否参与这一过程尚不明确。
RANKL 和RANK 相互作用可诱导Ki67 和抗凋亡蛋白Bcl-2 的表达,在蜕膜基质细胞生长中起重要作用[14]。在本实验中,RANKL-/-小鼠卵巢颗粒细胞Ki-67 表达降低,细胞凋亡增加,说明RANKL 基因通过影响卵巢颗粒细胞增殖与凋亡影响颗粒细胞数量。卵巢颗粒细胞是合成及分泌雌激素的重要功能细胞,颗粒细胞增殖及分化是卵泡发育的重要特征之一,与卵细胞进行交互对话,营养和支持卵细胞的增长发育,在卵泡的激活、选择及排卵等过程中均具有重要作用,影响卵巢储备功能。本实验发现RANKL-/-小鼠雌激素合成能力显著降低,在体质量、动情频率、血清AMH 水平和始基卵泡数量方面有下降趋势,闭锁卵泡数量有增加趋势,但差异均无统计学意义,提示RANKL 基因可以影响老年小鼠颗粒细胞雌激素合成能力,但卵巢储备功能无显著改变。随着女性年龄的增加,卵巢功能逐渐衰退,主要表现为高促性腺激素及低雌激素水平,可造成骨密度降低、认知记忆功能低下、性功能障碍和心血管疾病等远期并发症,严重影响女性身心健康。RANKL 基因的基础研究为寻找改善更年期女性生活质量靶点提供理论依据。此外,本实验还发现RANKL-/-小鼠卵巢指数未见明显改变,但是子宫体积增大,子宫指数显著增加,这可能与RANKL 基因影响子宫腺体发育相关,但其具体机制尚不清楚。RANKL 主要作用于骨细胞,并且其作用机制已得到共识,近年来,越来越多的研究表明RANKL 和骨保护素的相互作用受氧化应激调节[15]。氧化应激参与卵母细胞成熟、颗粒细胞增殖和分化以及类固醇激素分泌,进而诱导卵巢功能衰竭[16]。实验发现RANKL-/-小鼠血清SOD 抑制率和GSH-Px 活性降低,血清MDA 活性增加,提示RANKL-/-小鼠氧化应激水平升高,可能参与RANKL 基因对颗粒细胞作用过程,但目前结果尚不能提示氧化应激是其作用机制,仍需后续进一步实验设计。
综上所述,RANKL 基因可以通过影响颗粒细胞增殖和凋亡影响雌激素合成能力,进而诱导女性潮热、盗汗、骨质疏松等更年期症状。尽管目前实验结果未提示卵巢功能显著改善,但这可能与实验存在的局限性相关。首先实验采用的是RANKL 基因敲除鼠,RANKL 基因对卵巢的作用是非特异性的,其对其他组织器官的影响可能削弱对卵巢的作用,后续应增加RANKL基因条件性敲除鼠相关研究,或者在RANKL-/-小鼠中增加RANKL 基因的补救实验,检测小鼠卵巢功能改变;其次,实验采用30 周龄的老年鼠,此年龄段野生型小鼠卵巢储备已显著下降;此外,RANKL-/-小鼠氧化应激水平升高,氧化应激与年龄密切相关,随着年龄的增加,氧化应激改变应该更加显著,在后续的实验中应进一步延长实验小鼠的周龄。最后,RANKL 与氧化应激的关系目前已得到共识,但RANKL 是否通过氧化应激影响卵巢颗粒细胞作用及其机制还需要更深入的基础研究。
猜你喜欢
保健文汇(2022年8期)2022-11-10
临床肝胆病杂志(2022年10期)2022-10-19
昆明医科大学学报(2022年4期)2022-05-23
昆明医科大学学报(2022年2期)2022-03-29
保健与生活(2020年18期)2020-09-26
家庭百事通·健康一点通(2019年7期)2019-08-16
家庭科学·新健康(2017年12期)2018-01-09
家庭百事通·健康一点通(2017年5期)2017-05-19
风湿病与关节炎(2016年12期)2017-01-14
妇女之友(2016年10期)2016-12-27
