
抗真菌乳酸菌的筛选及其在酸奶发酵中的应用
2021-10-22 00:21张颖刘同杰公丕民张兰威王淑梅易华西
食品与发酵工业 2021年19期
张颖,刘同杰,公丕民,张兰威,王淑梅,易华西*
1(中国海洋大学 食品科学与工程学院,山东 青岛,266000) 2(哈尔滨学院 食品工程学院,黑龙江 哈尔滨,150086)
酸奶在加工、包装以及运输和贮藏过程中,极易受到耐酸真菌的污染,造成安全危害、经济损失以及品牌形象恶化[1]。其间的主要污染微生物是酵母菌和霉菌,包括青霉、曲霉、毛霉、乳酸克鲁维酵母、毕赤酵母、马克思克鲁维酵母等[2-3]。乳品企业通常采用物理或化学技术控制酸奶真菌污染,物理方法诸如冷藏、辐照等,已被普遍采用;化学防腐剂经常与物理防腐技术结合使用来控制真菌的污染[4]。但是与酸奶变质相关的酵母菌等真菌,由于细胞内pH值低,对山梨酸等弱酸防腐剂具有较强抵抗力[5],保鲜效果并不理想。此外,大量使用化学防腐剂会导致真菌的抗药能力增强,某些菌株的抗性机制已被证实具有遗传能力[6]。因此开发安全、高效的生物防腐剂来预防和控制酸奶中的真菌污染对保障酸奶微生物安全,延长酸奶保质期具有重要意义。
乳酸菌被公认为是安全的食品级微生物[7-8]。有的乳酸菌对导致食品腐败变质的微生物具有抑制活性,可代替化学防腐剂,具有生物防腐剂的作用[9]。然而,将这类乳酸菌作为辅助菌株用于酸奶生产,可能会对酸奶本身的感官和理化性质产生不良影响。理想情况下,被选择的乳酸菌在保持其抗真菌性能的同时,对酸奶的稳定性和感官质量不应产生负面影响[10]。目前国内市售的商业酸奶防腐保鲜菌主要依赖进口,开发具有我国自主知识产权的酸奶防腐保鲜菌对保障和提升我国酸奶的品质与安全具有重大意义。针对这一行业问题,本研究以酸奶中常见腐败真菌为指示菌筛选具有抗真菌活性的乳酸菌,然后将其作为辅助发酵剂用于酸奶生产,通过对应用效果进行评价,探究其作为酸奶防腐保鲜菌的潜力。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株
140株乳酸菌:分离自人体肠道及乳制品,保存于中国海洋大学食品科学与工程学院功能性乳制品与益生菌工程研究室。指示菌:食品腐败真菌(马克思克鲁维酵母、季也蒙毕赤酵母、黄曲霉)、食品腐败细菌(金黄色葡萄球菌、大肠杆菌、沙门氏菌、李斯特菌)、酸奶发酵剂YF-L907(保加利亚杆菌和嗜热链球菌)。酸奶发酵剂,北京多爱特生物科技有限公司;黄曲霉保存于中国海洋大学应用微生物实验室;马克思克鲁维酵母、季也蒙毕赤酵母、娄地青霉,中国普通微生物菌种保藏中心。
1.1.2 培养基
MRS培养基、M17培养基、PDA培养基、LB培养基、BHI培养基、脱脂乳培养基,青岛海博生物技术有限公司。
1.1.3 主要实验材料
全脂乳粉、白砂糖。
1.2 仪器与设备
洁净工作台(SW-CJ-1D),苏州净化设备有限公司;乳品发酵监控仪(CINAC),法国Alliance-AMS公司;高速冷冻离心机(TG20KR-D),长沙东旺实验仪器有限公司;真空冷冻干燥机(Alpha 1-4),德国Marin Christ公司;全自动灭菌锅(LDZX-75KBS),上海申安医疗器械厂。
1.3 实验方法
1.3.1 抗真菌乳酸菌的筛选
乳酸菌按2%接种量接种至MRS液体培养基中,37 ℃培养,每24 h活化3次,发酵液经离心(8 000 r/min,4 ℃,5 min)后,收集上清液,冻干浓缩10倍,用0.22 μm无菌滤膜过滤收集备用。
采用琼脂平板扩散法[11]评估乳酸菌对腐败真菌的抑制作用。在培养皿中放入无菌牛津杯,倒入20 mL含指示菌浓度为105CFU/mL的半固体培养基,待培养基完全凝固后拔出牛津杯,往孔中加入180 μL浓缩发酵上清液,室温下扩散5 h,28 ℃培养24 h,对抑菌圈进行测量。
有机酸排除实验:用5 mol/L NaOH溶液调乳酸菌发酵上清液pH值至6.0,排除酸干扰,测定抑菌活性。
蛋白酶敏感实验:向上述上清液中依次加入1 mg/mL木瓜蛋白酶、胰蛋白酶以及胃蛋白酶,37 ℃水浴2 h后,测定抑菌活性。
1.3.2 抑菌谱测定
使用乳品中常见腐败真菌和细菌以及基础发酵剂菌株作为指示菌,利用琼脂扩散法测定抑菌活性。
1.3.3 菌株鉴定
1.3.3.1 形态学观察
目标菌株在固体培养基上培养至菌落形成,挑取菌落稀释成菌液,在玻璃片上固定后,经革兰氏染色后在显微镜下观察菌体形态。
1.3.3.2 16S rDNA分析
使用细菌试剂盒提取总DNA,使用引物1(5′-AGAGTTTGATCCTGGCTCAG-3′)、引物2(5′-CTAC-GGCTACCTTGTTACGA-3′)进行PCR扩增。PCR 反应扩增条件:94 ℃预变性5 min,94 ℃变性1 min,55 ℃退火 30 s,72 ℃延伸90 s,30次循环,72 ℃延伸10 min,PCR产物的测序由上海生工生物工程有限公司完成。测序结果于NCBI的GenBank数据库进行BLAST比对,采用MEGA 7.0构建系统发育树,确定分离得到的乳酸菌种属。
1.3.4 酸奶制备
全脂复原乳(全脂乳粉120 g/L,白砂糖60 g/L)加热至60 ℃水浴30 min,均质(65 ℃,20 MPa)后迅速分装至锥形瓶中,在105 ℃下高温杀菌5 min后,迅速冰浴冷却至42 ℃接种发酵剂,恒温发酵至pH 4.5左右,置于4 ℃下冷藏24 h后熟。
使用酸奶发酵剂YF-L907(保加利亚杆菌:108CFU/mL,嗜热链球菌:108CFU/mL)与F32-2复配发酵,酸奶分为3组,F32-2 添加量分别为0、1×106和1×107CFU/mL。酸奶冷藏3 d后进行感官评价,贮藏期间于第1、7、14、21、28天测量相关理化指标和微生物指标。
1.3.5 感官评价
参照中国乳制品工业行业规范RHB 103—2004《酸牛乳感官质量评鉴细则》,选取10名评价员从色泽(10分)、组织状态(10分)、口感与气味(10分)等3个方面对酸奶进行感官评价。
1.3.6 酸奶理化指标测定
pH值测定:发酵期间酸奶pH使用CINAC乳品发酵监控仪跟踪记录,贮藏期间酸奶pH变化使用pH计测量。
可滴定酸度测定:参照GB 5009.239—2016《食品安全国家标准 食品酸度的测定》。
1.3.7 酸奶微生物指标测定
参照GB 4789.2—2016《食品安全国家标准 食品微生物学检验 菌落总数测定》,其中乳球菌计数使用M17培养基,乳杆菌计数使用MRS 培养基。
1.3.8 抑制酸奶中腐败真菌活性测定
霉菌抑制活性测定:将样品(各3杯平行样)置于28 ℃下恒温放置4周,定期观察有无霉菌生长并予以记录[12]。如发现有1杯样品中有霉菌生长,则观察结果记录为1。
酵母菌抑制活性测定:酸奶发酵结束后添加马克思克鲁维酵母(100±20)个/mL,置于4 ℃下冷藏4周后进行平板计数,采用孟加拉红培养基,按照公式(1)计算酵母菌的抑制率[13]。
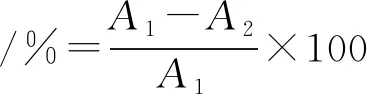
(1)
式中:A1,对照组酵母菌活菌数;A2,实验组酵母菌活菌数。
1.4 数据分析
采用SPSS 22.0软件对结果进行差异显著性分析(P<0.05),采用Origin 8.5软件作图。
2 结果与分析
2.1 抗真菌乳酸菌的筛选及其抑菌谱研究
以马克思克鲁维酵母、季也蒙毕赤酵母、黄曲霉为指示菌,从140株乳酸菌筛选出具有明显抑菌效果的菌株F32-2,其对马克思克鲁维酵母的抑菌效果最明显。将马克思克鲁维酵母作为指示菌进一步对菌株F32-2的抑菌物质进行了初步摸索。经过酸排除实验后,菌株F32-2的发酵上清液保留了大部分抑菌活性,但在经过不同蛋白酶处理后抑菌活性消失,初步确定主要抑菌物质为蛋白类或肽类。
进一步测定了菌株F32-2的抑菌谱,结果如图1所示。菌株F32-2不仅对3株腐败真菌有抑制作用,对金黄色葡萄球菌、大肠杆菌、沙门氏菌、李斯特菌也表现出明显的抑制效果。另外,菌株 F32-2对酸奶发酵剂中的主体菌株保加利亚杆菌抑制作用较小,对嗜热链球菌基本无抑制,具备酸奶辅助发酵剂和酸奶保鲜菌的潜质。据此,以菌株F32-2为研究对象,开展其在酸奶保鲜中的应用研究。

图1 菌株F32-2对季也蒙毕赤酵母 (a),马克思克鲁维酵母 (b),黄曲霉(c),大肠杆菌(d),金黄色葡萄球菌(e),李斯特菌(f),沙门氏菌(g),保加利亚杆菌(h)和嗜热链球菌(i)的抑制效果Fig.1 Inhibition of strain F32-2 on Meyerozyma guilliermondii (a),Kluyveromyces marxianus (b),Aspergillus plarus (c),Escherichia coli (d),Staphyloccocus aureus (e),Listeria monocytogenes (f),Salmonella paratyphi (g), Lactobacillus bulgaricus (h),and Streptococcus thermophilus (i)
2.2 菌株鉴定
2.2.1 菌体形态观察
对菌株F32-2进行革兰氏染色,在显微镜下观察菌体形态如图2所示,菌体呈短杆状,F32-2为革兰氏阳性杆菌。

图2 F32-2菌体显微镜观察形态(×1 000)Fig.2 Microscopic observation of F32-2 (×1 000)
2.2.2 16S rDNA分析鉴定
将菌株F32-2的16S rDNA鉴定结果在NCBI的GenBank数据库进行BLAST比对,其与鼠李糖乳杆菌的相似度达到99%,使用MEGA 7.0软件构建系统发育树如图3所示,菌株F32-2与鼠李糖乳杆菌为同一分支,鉴定其为鼠李糖乳杆菌,命名为鼠李糖乳杆菌F32-2(LactobacillusrhamnosusF32-2)。

图3 菌株F32-2的系统发育树Fig.3 The phylogenetic tree of strain F32-2
2.3 L.rhamnosus F32-2对酸奶发酵时间的影响
按1.3.4中分组制备了酸奶样品,使用CINAC乳品发酵监控仪跟踪记录酸奶发酵过程的pH变化,pH随发酵时间的动态变化曲线如图4所示。仅接种酸奶发酵剂YF-L907的酸奶在3.8 h到达发酵终点(pH=4.5),辅助接种L.rhamnosusF32-2的2组酸奶在发酵后期pH均低于对照组。L.rhamnosusF32-2接种量为1×106和1×107CFU/mL的酸奶发酵时间分别缩短了0.3和0.5 h,表明L.rhamnosusF32-2在酸奶发酵过程中没有对保加利亚杆菌和嗜热链球菌产生拮抗作用,且本身也发酵产生乳酸,使酸奶提前达到发酵终点,缩短了发酵时间。

图4 L.rhamnosus F32-2对酸奶发酵时间的影响Fig.4 Effect of L.rhamnosus F32-2 on yogurt fermentation
2.4 L.rhamnosus F32-2对酸奶感官品质的影响
感官品质是消费者接受或拒绝食品最重要的标准[14]。基于1.3.5的评分标准,从酸奶的色泽、组织状态、口感与气味等3方面进行感官评价,评分结果如图5所示。不同组酸奶的3个指标评分均在8~9分,组间差异不显著(P>0.05),表明L.rhamnosusF32-2对酸奶的感官品质无显著影响。
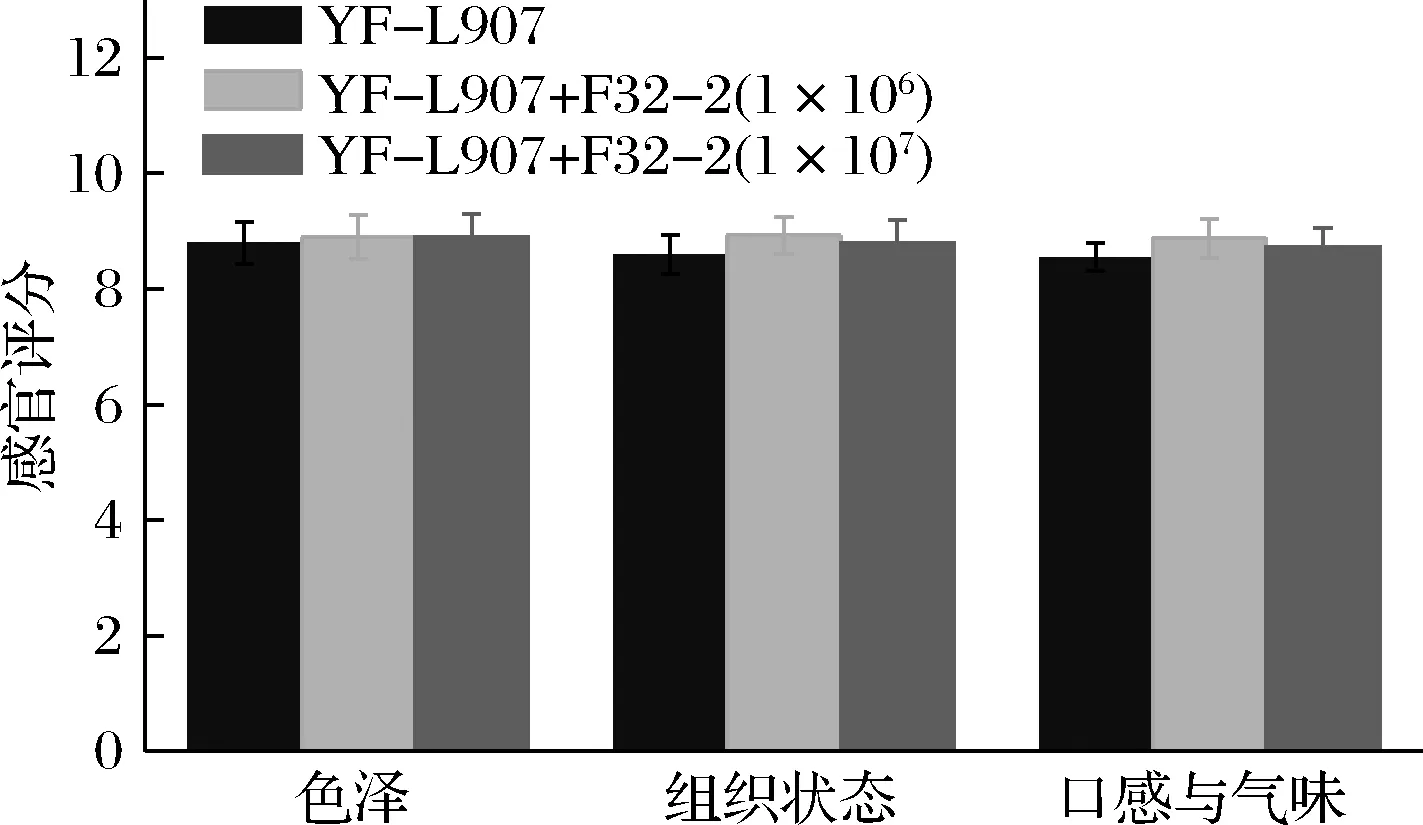
图5 酸奶感官评价结果Fig.5 Sensory evaluation results of yogurt
2.5 L.rhamnosus F32-2对酸奶冷藏期间pH值和可滴定酸度的影响
在酸奶发酵结束后,乳酸菌继续将乳糖转变为乳酸,改变酸奶pH值和酸度[15]。图6显示了酸奶在4 ℃贮藏期间pH值和可滴定酸度的变化。当L.rhamnosusF32-2接种量为1×106CFU/mL,pH值和可滴定酸度与对照组酸奶无显著性差异(P>0.05);当酸奶L.rhamnosusF32-2的接种量达到1×107CFU/mL,贮藏至第21天后,酸奶pH值显著高于对照组(P<0.05),可滴定酸度显著低于对照组(P<0.05)。
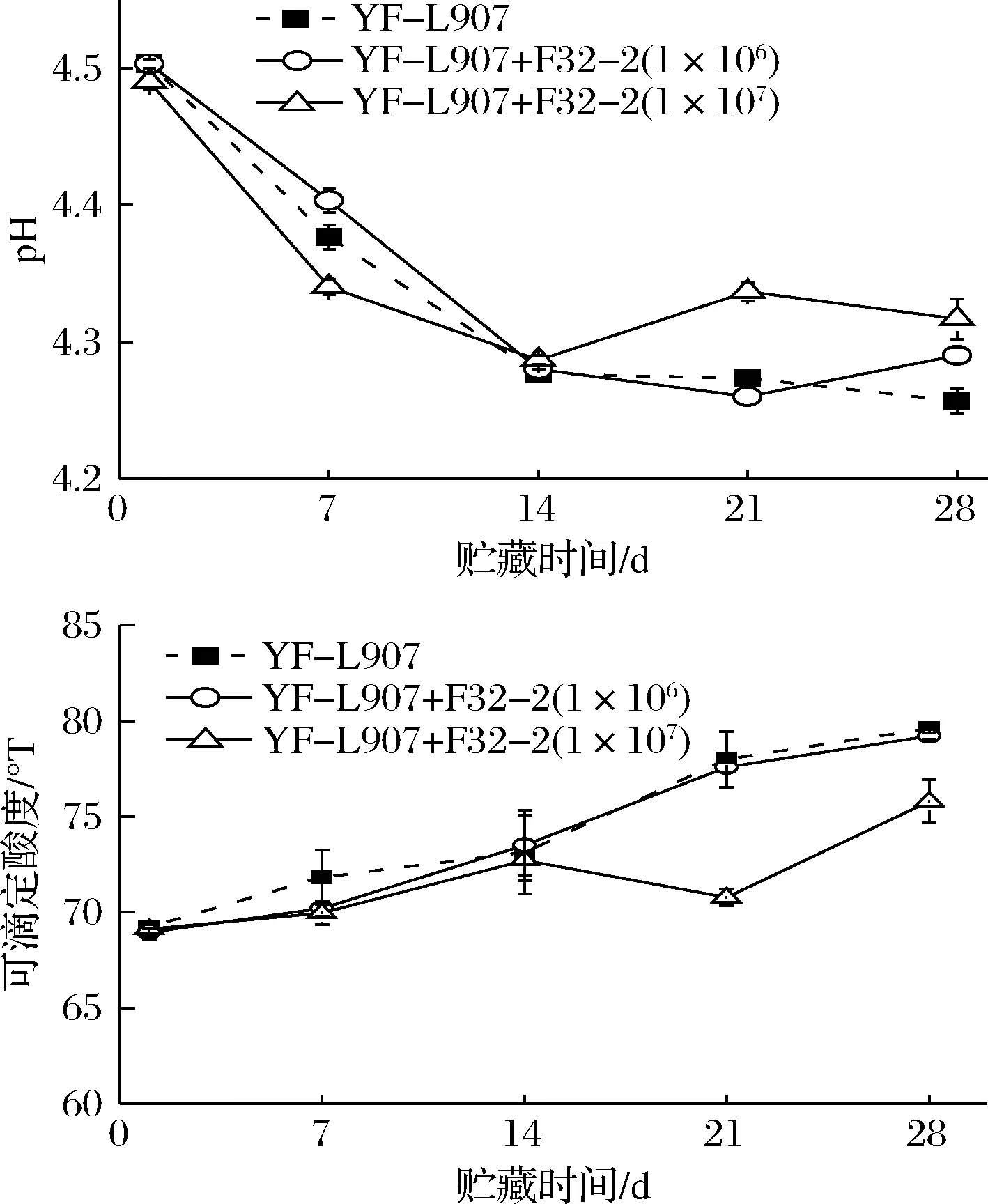
图6 4 ℃下酸奶贮藏期间pH值和可滴定酸度的变化Fig.6 Changes of pH and TA of yogurt during storage at 4 ℃
贮藏时间和L.rhamnosusF32-2接种量均能影响酸奶的酸度,加大L.rhamnosusF32-2接种量,能在贮藏后期延缓酸奶的后酸化,这与酸奶贮藏期间乳酸菌的种类、数量和发酵活性有关。保加利亚杆菌是导致酸奶后酸化的主要菌株[16]。贮藏后期,L.rhamnosusF32-2增殖成为酸奶中的优势菌,抑制了保加利亚杆菌的生长,从而一定程度上缓解了酸奶的后酸化。
2.6 L.rhamnosus F32-2对酸奶冷藏期间活菌数的影响
酸奶冷藏期间乳球菌和乳杆菌总数的变化如图7所示。各组酸奶的乳球菌数随着时间的延长而降低,仅接种发酵剂YF-907的对照组酸奶乳球菌总数最多。辅助接种L.rhamnosusF32-2后,酸奶中的乳球菌总数都略微减少,表明L.rhamnosusF32-2在酸奶冷藏期抑制了嗜热链球菌的生长,但无显著性差异(P>0.05)。3组酸奶中乳杆菌总数的变化趋势一致,均先下降后上升,且始终保持在108CFU/mL之上。接种L.rhamnosusF32-2的2组酸奶的乳杆菌总数均显著增加(P<0.05),且随时间变化的波动幅度更小,表明L.rhamnosusF32-2的添加提高了酸奶中的乳杆菌总数。在酸奶贮藏后期,L.rhamnosusF32-2可一定程度上抑制保加利亚杆菌的生长,成为酸奶中的优势菌群,从而使乳杆菌总数回升。LI等[12]发现干酪乳杆菌AST18的添加会提高酸奶中的乳杆菌总数,同时抑制乳球菌的生长,与实验结果一致。

图7 4 ℃下酸奶贮藏期间活菌数的变化Fig.7 Changes of viable count of yogurt during storage at 4 ℃
2.7 L.rhamnosus F32-2对酸奶中腐败真菌的抑制效果
前期研究中L.rhamnosusF32-2对霉菌的抑制实验是在培养基体系中进行,为验证L.rhamnosusF32-2在酸奶体系中对霉菌的抑制效果,将酸奶置于常温下贮藏。结果如表1所示,在第21天,仅接种发酵剂YF-L907的酸奶出现明显的霉菌污染,L.rhamnosusF32-2接种量为1×106CFU/mL的酸奶在第28天出现污染,L.rhamnosusF32-2接种量为1×107CFU/mL的酸奶在贮藏期内没有出现污染。
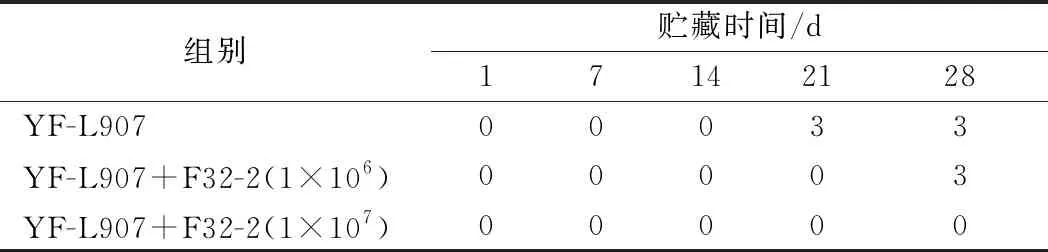
表1 L.rhamnosus F32-2 对酸奶中霉菌的抑制效果Table 1 Inhibitory effect of L.rhamnosus F32-2 on mold in yogurt
贮藏28 d后的酸奶如图8所示,YF-L907组酸奶表面长出了白色霉菌菌落,且伴随着严重的乳清析出;YF-L907+F32-2(1×107)组酸奶表面保持光滑,质地均匀无明显乳清析出,表明L.rhamnosusF32-2可有效抑制酸奶中霉菌的生长。L.rhamnosusF32-2的抑菌效果体现出剂量依赖性,当添加量达到1×107CFU/mL时,酸奶在常温下的贮藏期能延长到28 d以上。
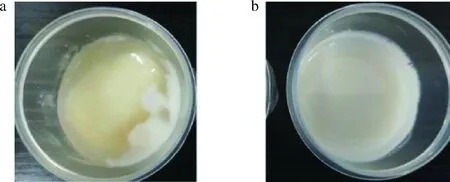
a-YF-L907组酸奶;b-YF-L907+F32-2(1×107)组酸奶图8 酸奶的货架期观察实验Fig.8 Shelf life saving experiment of yogurt
由于无法使用直接观察法研究酸奶中酵母菌的生长情况,于是设置了酵母菌污染酸奶实验。在酸奶发酵结束后接种马克思克鲁维酵母,4 ℃下贮藏28 d后进行酵母菌计数,计算酵母菌的抑制率,结果如表2所示。当L.rhamnosusF32-2 的添加量为1×106和1×107CFU/mL时,马克思克鲁维酵母菌的抑制率分别达到了39.28%和66.09%,与对照组相比酵母存活数均显著降低,且具有剂量效应。本研究结果与郭建林等[17]的研究结果相似,酵母菌污染后能长期存活在酸奶中,保鲜菌接种量越高,贮藏结束后酵母菌数越少。DELAVENNE等[13]同样发现,哈尔滨乳杆菌K.V9.3.1 Np对解脂耶氏酵母的抑制作用具有明显的剂量依赖性。关于L.rhamnosusF32-2对马克思克鲁维酵母的抑制机理有待进一步的深入研究。
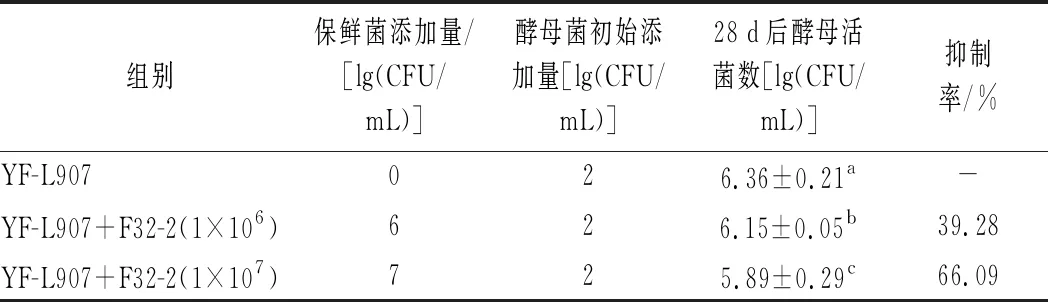
表2 L.rhamnosus F32-2对酸奶中酵母菌的抑制效果Table 2 Inhibitory effect of L.rhamnosus F32-2 on yeast in yogurt
3 结论
以导致酸奶腐败变质的典型真菌为指示菌,筛选出1株具有明显抑制真菌活性的L.rhamnosusF32-2,主要抑菌产物为蛋白类物质。将其作为辅助发酵剂用于酸奶发酵,对酸奶的发酵时间、感官品质、活菌总数均无负面影响,且在酸奶贮藏后期,能有效改善酸奶后酸化现象,在酸奶体系中能有效抑制霉菌和酵母菌的生长,延长酸奶的货架期,具有酸奶辅助发酵剂和保鲜菌的开发前景。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
食品与发酵工业(2022年2期)2022-02-22
河南畜牧兽医(2021年9期)2021-12-10
中国土壤与肥料(2021年5期)2021-12-02
生物学通报(2021年5期)2021-03-16
军事文摘·科学少年(2021年1期)2021-02-04
中国酿造(2020年10期)2020-11-04
当代水产(2019年3期)2019-05-14
食品界(2018年10期)2018-11-23
小学阅读指南·低年级版(2016年10期)2016-09-10
