
刺槐多倍体诱导及叶片解剖结构分析
2021-10-13 07:38张丞慧孟冰超杨茂磊马洪峥胡宝全
天津农学院学报 2021年3期
张丞慧,孟冰超,杨茂磊,马洪峥,胡宝全
刺槐多倍体诱导及叶片解剖结构分析
张丞慧,孟冰超,杨茂磊,马洪峥,胡宝全通信作者
(天津农学院 园艺园林学院,天津 300392)
四倍体刺槐因其具有适应性强、生物量大等特点,被广泛推广种植。本研究采用展开子叶的刺槐幼苗为材料,通过5种浓度的秋水仙素处理茎尖,诱导刺槐四倍体,并研究诱导后四倍体叶片解剖结构的变化。研究发现0.05%的秋水仙素处理幼苗1 d能取得较好的诱导效果。四倍体刺槐的叶片解剖结构中,上表皮厚度、栅栏组织、海绵组织和叶片厚度等都显著高于二倍体,四倍体叶片紧实度显著提高,表明刺槐四倍体化后有较强的抗逆性。研究结果可为刺槐多倍体育种提供可靠的技术支持,并为刺槐的倍性育种工作提供参考和理论支持,同时为阐述四倍体刺槐抗逆性形成原理提供形态解剖学依据。
刺槐;四倍体;秋水仙素;叶片;解剖结构
多倍体化是植物界的一种普遍现象,广泛存在于被子植物中,是植物进化和新物种形成的重要途径。因多倍体植物普遍具有营养器官巨大、抗逆性高和生活力强的优点,被作为一种重要的育种途径而广泛使用。秋水仙素是从百合科植物秋水仙中提取出的一种能抑制纺锤丝的化学诱变剂,被广泛用于植物多倍体的人工诱导中,常见的诱导方法包括浸渍法、喷涂法、注射法和培养基诱导法等。
刺槐是蝶形花科二倍体植物,适应性、耐盐性强[1],用途广泛,是重要的生态防护树种。四倍体刺槐最早由韩国诱导成功,具有生长迅速、生物量大、抗逆性强等特点[2]。引入我国的四倍体刺槐,作为用材树种其木质坚韧细致,作为青饲料来源其营养价值高,被广泛引种到我国北部地 区[3]。前人对四倍体刺槐的栽培、组织培养和生理生化特性进行了大量研究,但四倍体刺槐的种质创新工作进展仍为缓慢[2]。
研究人员通过对刺槐人工诱导,获得了新的四倍体刺槐种质资源,如罗晓丹以刺槐愈伤组织和刺槐种子为材料,结合秋水仙碱浸渍法与低温处理法,诱导得到刺槐四倍体,其中60mg/L的秋水仙素浸渍法处理刺槐种子2 d后诱导率最高,达25.71%[4]。姜丹使用0.3%的秋水仙素浸泡刺槐初露白尖的种子3 d后获得诱导四倍体[5]。冯玥等通过秋水仙素诱导及继代培养检测稳定性,认为0.4%的秋水仙素浸渍处理刺槐种子2d,诱导多倍体效率最高[6]。刺槐多倍体的诱导虽然取得了一些进展,但组培繁琐耗时,且以往的研究缺乏对获得的四倍体刺槐的解剖特征的进一步分析,而叶片特征的变化能够更直接反映植物对环境的适应能力,其抗逆性的强弱可从解剖结构的变化中分析得出。
本试验采用改进的方法诱导刺槐四倍体,并对诱导后的刺槐四倍体的叶片解剖结构进行分析,探究四倍体化对刺槐叶片结构的影响,以期为探索四倍体刺槐优势的形成提供理论基础。
1 材料与方法
1.1 试验材料
试验以刚展开子叶的刺槐幼苗为材料,种子采自天津农学院校园经鉴定为二倍体的成年刺槐。刺槐种子经60℃温水浸种,自然冷却到室温后浸泡过夜,置于培养皿中覆盖纸巾,于22 ℃温箱中催芽,保持湿润,待种子展开子叶未抽真叶时进行处理。
1.2 四倍体刺槐诱导
以5种不同浓度的秋水仙素(0.40 %、0.20 %、0.10 %、0.05%、0.01%)溶液,22 ℃下浸泡刺槐幼苗茎尖生长点24h,蒸馏水清洗后,种植于基质(草碳∶蛭石∶珍珠岩=1∶1∶1)中培养,每个处理梯度20~25株,对照5株,重复3次。
1.3 染色体鉴定
取茎尖于饱和对二氯苯溶液中,20℃浸泡 3h,取出茎尖经蒸馏水清洗后,固定于卡诺固定液中,固定24h后取出,蒸馏水清洗3次,于1mol/L盐酸中55 ℃酸解10min,蒸馏水清洗3次后,置于卡宝品红溶液中染色1h。取染色后的茎尖分生区组织,于载玻片上经盖玻片压片后,显微镜镜检,选取5个视野对染色体计数。
1.4 气孔及保卫细胞测量
撕取刺槐叶片表皮于载玻片上,于显微镜下测量气孔及保卫细胞长宽,每个植株选取5片叶片,选取5个视野,每个视野下统计5个气孔及保卫细胞的长宽,未处理植株作为对照,数据使用SPSS19.0,采用方差分析进行显著性检验。
1.5 叶片及茎的解剖结构观察
每株刺槐取健康叶片,以主脉为对称轴,切成宽度为5mm的小段,于FAA固定液中处理24h后,梯度乙醇脱水,二甲苯透明,石蜡包埋,于LeicaRM2235切片机上切片,番红固绿染色,中性树胶封片,LeicaDM4000B显微镜观察,LAS4.2软件测量,每株叶片样本分别测量10次。
2 结果与分析
2.1 秋水仙素对刺槐幼苗的抑制作用
不同浓度秋水仙素处理刺槐幼苗后,存活幼苗见表1,发现秋水仙素浓度越高,植株死亡率越高,0.40%秋水仙素处理幼苗存活率仅为8.3%,0.01%处理存活率为60.0%,明显观察到秋水仙素对刺槐幼苗的抑制作用,且抑制程度与秋水仙素浓度成正比。

表1 秋水仙素处理结果 %
2.2 刺槐多倍体诱导结果
通过刺槐对照株与变异株的观察,变异植株,叶片明显增大,发现同一生长阶段的变异株叶片气孔长度、宽度,保卫细胞的长度、宽度与对照株相比差异均达到极显著水平,且变异株叶片气孔和保卫细胞长度、宽度相较于对照株大一倍左右(表2)。对照株和变异株刺槐分别统计10个分裂中期细胞,经统计计数后发现,对照株染色体数为2=22,变异株的染色体数为4=44。通过气孔分析结果和染色体计数,确定变异株为四倍体(图1)。
统计诱导成功的刺槐发现,0.05%和0.01%的秋水仙素处理刺槐幼苗,成功率为30.7%和29.3%,但0.01%诱导后,植株存活率高,0.05%处理组诱导成功的占存活刺槐的75.9%且存活率较高,因此浸泡法诱导刺槐幼苗多倍体的最佳秋水仙素浓度为0.05%。
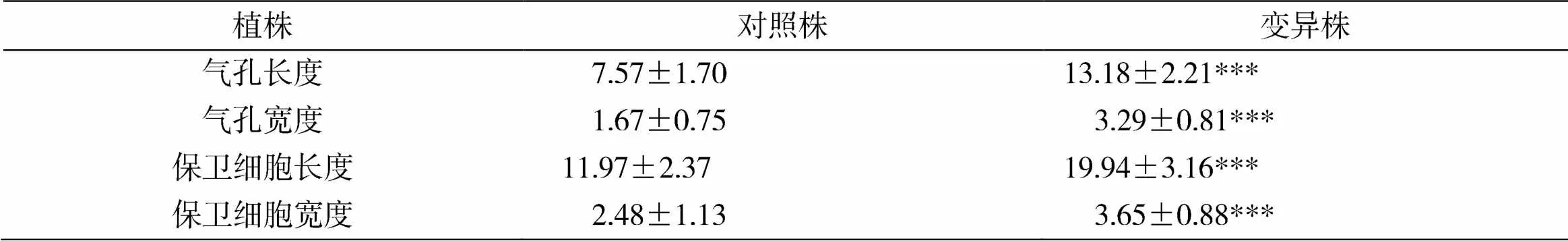
表2 刺槐对照株与变异株气孔及保卫细胞大小比较μm
注:***表示在<0.001水平上差异显著
注:A为四倍体刺槐;B为对照二倍体
2.3 叶片解剖结构比较
对二倍体刺槐和四倍体刺槐的叶片解剖结构分析发现(表3),四倍体刺槐除下表皮厚度与二倍体植株差异不显著外,上表皮厚度、栅栏组织、海绵组织、叶肉厚度、叶片厚度、维管束面积和最大导管面积均显著高于对照二倍体(<0.05),其中栅栏组织、叶肉厚度、叶片厚度和维管束面积达到极显著水平(<0.001)。内含大量叶绿体的栅栏组织变厚,表明四倍体刺槐可能含有更多的叶绿体,提高了单位面积叶片的光合作用效率,积累生物量的速度加快。而四倍体刺槐叶片维管束面积的极显著增加,则表明四倍体刺槐在水分和无机盐的运输上可能比二倍体效率 更高。

表3 不同倍性刺槐叶片细胞指标差异分析 μm
注:<0.05、<0.01和<0.001分别表示为*、**、***。下同
进一步比较叶片组织结构的紧密度、结构疏松度和上下表皮相对厚度(图2),结果表明,四倍体刺槐只有叶片紧密度显著高于二倍体,其他3个指标均差异不显著(表4)。这表明四倍体叶片变厚的过程中,组织变得紧密,这种紧密的组织排列可能是多倍体抗逆性提高的体现。
注:A为对照二倍体;B为四倍体刺槐

表4 不同倍性刺槐叶片细胞相关指标差异分析 %
3 讨论
倍性育种是林木遗传育种工作中一种重要的种植创新手段,如通过多倍体诱导结合杂交育种,我国选育出了速生优质三倍体毛白杨。在多倍体诱导中,秋水仙素作为最有效的细胞有丝分裂阻断剂,被广泛用于林木多倍体的诱导。但秋水仙素处理植物材料,本身对植物存在毒害作用,处理后造成幼苗生长缓慢、器官畸形等问题[7],甚至发育停止并死亡。因此在林木多倍体的诱导中,选取适宜的秋水仙素的浓度和处理时间,成为影响多倍体诱导效果的重要因素。本试验获取四倍体刺槐的过程中,观测到经过秋水仙素处理后的部分幼苗中,存在明显的生长抑制现象,且抑制作用和致死率与秋水仙素的浓度呈正相关。
以往研究中,对刺槐多倍体的诱导多采用组织培养,在培养基中添加秋水仙素诱导四倍体[3],步骤繁琐耗时。而由于刺槐易萌发,浸泡种子诱导操作简便,是研究人员诱导刺槐四倍体的主要方式[5-6]。但以往诱导种子的时期均为种子初露白时,进行秋水仙素浸泡处理,需要秋水仙素的浓度高,需要0.3%或0.4%的秋水仙素处理2~3 d才能取得较好的诱导效果[5-6]。本试验采用初展开子叶的刺槐为处理材料,秋水仙素的处理浓度和时间明显减少,0.05%处理1 d即可较好的获得加倍的刺槐,提高了诱导效率,诱导方法简单且时间短。造成这种现象的原因,可能与种子种皮对秋水仙素渗透速度的影响有关,去掉种皮的初萌动种子或子叶展开的幼苗,秋水仙素更容易渗透到分生组织。
植物叶片是对环境变化最敏感的器官[8],对植物叶片的解剖结构分析可反映植物对环境的适应能力。植物多倍化能引起植物的叶片厚度等方面发生明显变化[9-10]。这与本试验中发现的结果一致,刺槐四倍体化后,上表皮厚度、栅栏组织、海绵组织和叶片厚度等都显著高于二倍体,这表明四倍体刺槐叶片的结构发生了巨大变化。进一步比较发现,刺槐四倍体的紧实度显著高于二倍体,而疏松度、上表皮相对厚度和下表皮相对厚度都变化不显著,这说明刺槐四倍体叶片器官巨大化过程中基本是同比例变化。这种变化也可能与基因组加倍后的复杂性和个体差异有关。有研究认为多倍化不一定导致所有植物叶片解剖结构指标发生显著变化[11],如杨桂英等对不同倍性的山茶叶片解剖结构的分析发现,六倍体山茶叶片厚度和海绵组织厚度与二倍体比较变化不显著[12]。邱芬等分析多倍体化滇杨发现个别多倍体材料叶片紧实度和上表皮相关对厚度等指标并不显著高于二倍体[11],说明多倍化对叶片的巨大化受个体本身基因组差异的影响,多倍体诱导育种过程需要进行较细致的性状测定比较。
植物的叶片长期与环境接触,环境胁迫的压力会影响叶片的解剖结构,因此,研究叶片的解剖结构可以反映其抗逆性的强弱[13]。如不同倍性的滇山茶的上下表皮厚度可作为评价耐寒性的指标[14]。而四倍体刺槐的栅栏组织极显著高与二倍体,强化的栅栏组织被认为可提高叶片的紧实程度,试验中也检测到四倍体刺槐叶片的紧实度显著高于二倍体。这种紧密的叶片结构在低温环境下为叶片提供更好的防护[15]。因此,分析认为,四倍体刺槐的抗逆性应强于二倍体,尤其在抗寒性等方面优于二倍体。本研究获得的刺槐四倍体加倍成功的是地上部分,要获得完整的四倍体尚需经过扦插生根过程。
4 结论
本试验采用展开子叶的刺槐幼苗为材料,通过不同浓度秋水仙素的处理,诱导刺槐四倍体,并研究诱导后四倍体叶片解剖结构的变化,发现0.05%的秋水仙素处理幼苗1 d能取得较好的诱导效果,四倍体刺槐的叶片解剖结构,上表皮厚度、栅栏组织、海绵组织和叶片厚度等都显著高于二倍体,四倍体叶片紧实度显著提高,表明其有较强的抗逆性。结果可为刺槐的倍性育种工作提供参考和理论支持。
[1]张瑞芳,侯澄辉,刘子敬,等.混合盐胁迫对刺槐和绒毛白蜡根系生长及其解剖结构的影响[J].天津农学院学报,2018,25(1):14-18,24.
[2]聂琳,彭杰,常军,等.四倍体刺槐研究现状及进展[J]中国农学通报,2013,29(4):1-4.
[3]张乃锋,刁其玉,韩凤宝,等.四倍体刺槐对奶牛产奶性能及乳成分的影响[J]饲料研究,2009(1):61-63.
[4]罗晓丹. 四倍体刺槐诱导及其体外植株再生的研究[D].长沙:中南林业科技大学,2009.
[5]姜丹. 刺槐子叶不定芽再生及多倍体诱导的研究[D]. 北京:北京林业大学,2012.
[6]冯玥,任云辉,李秀宇,等.人工诱导多倍体刺槐及其倍性鉴定[J]分子植物育种,2018,16(21):254-261.
[7]EWALD D,ULRICH K,NAUJOKS G,et al. Induction of tetraploid poplar and black locust plants using colchicine:chloroplast number as an early marker for selecting polyploids in vitro[J]. Plant Cell Tissue & Organ Culture,2009, 99(3): 353-357.
[8]康萨如拉,牛建明,张庆,等.短花针茅叶片解剖结构及与气候因子的关系[J]草业学报,2013,22(1):77-86.
[9]李凤兰,张志毅,张民侠.白杨染色体加倍技术研究及三倍体育种(Ⅲ)──加倍体某些形态特征的观察[J]北京林业大学学报,1994(2):15-18.
[10]唐军荣,李斌,朱丽娜,等.滇杨多倍体苗期叶片形态及光合生理比较分析[J]林业科学研究,2016,29(1):103-109.
[11]邱芬,辛亚龙,唐军荣,等.不同倍性滇杨叶片解剖结构差异分析[J]中南林业科技大学学报,2017,37(6):56-59,69.
[12]杨桂英,王兵益,何瀚,等.从叶片解剖结构探讨云南山茶不同倍性的耐旱潜力[J]西南农业学报,2015,28(6):2714-2719.
[13]陈雪峰,景晨娟,赵习平,等.植物叶片组织结构在抗逆研究中的应用进展[J]河北农业科学,2018,22(3):50-53.
[14]杨桂英,何瀚,曹子林,等.3种不同倍性滇山茶的耐寒性研究[J].云南大学学报(自然科学版),2016,38(2):335-343.
[15]马英姿,梁文斌,陈建华.经济植物的抗寒性研究进展[J]经济林研究,2005(4):89-94.
Induction of polyploidL.and analysis of anatomical structure of leaves
Zhang Chenghui, Meng Binchao, Yang Maolei, Ma Hongzheng, Hu BaoquanCorresponding Author
(College of Horticulture and Landscape, Tianjin Agricultural University, Tianjin 300392, China)
TetraploidL. has been widely planted because of its strong adaptability and large biomass. In this study, the seedlings ofwere treated with 5 different concentrations colchicine to induce tetraploid, and the changes of the anatomical structure of the tetraploid leaves after induction were studied. It was found that 0.05% colchicine could achieve better induction effect in one day. The thickness of the upper epidermis, the palisade tissue, sponge tissue and leaf thickness of the tetraploidleaves were significantly higher than those of diploid, and the cell tense ratio of tetraploid ones were significantly improved, which indicated thathad strong resistance to stress after tetraploidization. The results can provide reliable technical support for the polyploid breeding of,provide reference and theoretical support for the ploidy breeding of,and provide morphological and anatomical basis for explaining the formation principle of the resistance of tetraploid.
L.; tetraploid; colchicine; leaves; anatomical structure
1008-5394(2021)03-0018-04
10.19640/j.cnki.jtau.2021.03.004
Q944.52;S722.3
A
2020-06-12
国家自然科学基金(3180011064);天津市大学生创新训练计划项目(201810061251)
张丞慧(1998—),男,本科在读,主要从事林木倍性育种方面的研究。E-mail:18149296066@126.com。
胡宝全(1982—),男,助理研究员,博士,主要从事林木遗传育种方面的研究。E-mail:baoquan-hu@126.com。
责任编辑:杨霞
猜你喜欢
西北农业学报(2022年5期)2022-06-07
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年12期)2022-01-14
山西农业科学(2021年3期)2021-12-06
东北林业大学学报(2021年12期)2021-02-10
诊断学(理论与实践)(2020年6期)2020-12-09
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
江苏农业科学(2016年9期)2016-11-28
江苏农业科学(2014年9期)2014-11-15
