
高丹草F2代农艺性状的数量遗传分析
2021-10-13 07:47:56董林洁包曙光康晨谢晓东罗峰孙守钧
天津农学院学报 2021年3期
董林洁,包曙光,康晨,谢晓东,罗峰,孙守钧
高丹草F2代农艺性状的数量遗传分析
董林洁,包曙光,康晨,谢晓东,罗峰通信作者,孙守钧通信作者
(天津农学院 农学与资源环境学院,天津 300392)
以粒用高粱‘忻粱52’和苏丹草‘美引-48’杂交构建的F2代群体为试验材料,对F2代的叶片数、茎节数、株高、茎粗和蜡粉等5个农艺性状进行测定,并利用主基因+多基因混合遗传模型分析得到各性状的最适遗传模型及遗传率。结果表明:叶片数、茎粗的最适模型为Model A_0,表明该性状不受主基因控制,为微效多基因控制的性状;茎节数和株高的最适模型为Model A_4,为负向完全显性混合遗传模型,受一对主基因遗传,主基因遗传率分别为91.13%和66.14%;蜡粉的最适模型为Model B_1,由两对主基因控制的加性-显性-上位性混合遗传模型,控制该性状的主基因遗传率为54.03%。
高丹草;农艺性状;主基因+多基因分析;数量遗传
高粱()和苏丹草()同为禾本科高粱属、一年生草本植物,二者的杂交种简称高丹草[1]。该品种不仅吸取了高粱茎秆粗、叶片宽大,苏丹草分蘖再生能力强等优点,还兼具两者耐旱、耐涝和耐盐碱等优点,表现出极强的抗逆性。其生物产量高,适口性较好,可多次刈割,也可在蜡熟期只刈割一次进行青贮、晒制干草等[2-4]。但饲草高粱茎秆表面附着明显的蜡粉,降低了牲畜对饲料的消化率[5],不利于营养物质的吸收,限制了饲草高粱的应用。目前国内外研究主要集中在某类作物蜡质合成与分泌过程中的相关基因方面[6-7],而对作物表皮蜡粉与植物农艺性状的研究较少。因此,本研究对高丹草F2代的叶片数、茎节数、株高、茎粗和蜡粉5个农艺性状进行数量遗传分析,旨在探究高丹草不同农艺性状的遗传规律,提高育种工作的预见性,减少盲目性[8],为下一步分子育种工作提供依据。
1 材料与方法
1.1 试验材料
本试验选取粒用高粱‘忻粱52’和苏丹草‘美引-48’杂交得到F2代群体400株。
1.2 试验设计
于2016年8月在天津市静海良种场进行亲本选择,同年12月在海南省乐东黎族自治县利国镇官村进行亲本杂交,获得杂交种F1代。2017年5月在静海良种场种植亲本及F1代,8月观察F1代正反交长势,经检验,F1代正反交长势相同,排除细胞质遗传,将F1套袋使其自花授粉以获得F2代杂交种。2020年5月将亲本及杂交种种植于天津市宁河区天津实验林场,播种亲本各1行、F2代50行,行长5m,行距0.5m,株距0.15m,田间管理同大田。在开花期对高粱茎秆蜡粉含量进行测定,在完熟期对高粱叶片数、茎节数、株高、茎粗进行测定。
1.3 测定方法
由于F2代发生性状分离,因此以下性状测量均为单株数据。
(1)叶片数:在完熟期,计算整株叶片数量;
(2)茎节数:在完熟期,计算整株茎节数;
(3)株高:在完熟期,用钢卷尺测量地面至穗顶部的距离;
(4)茎粗:在完熟期,用游标卡尺测量茎秆中部茎粗;
(5)蜡粉:在开花期,用色彩色差计测量茎秆中部蜡粉含量。
1.4 数据处理
基础数据采用MicrosoftExcel2010整理,用SPSS26.0进行相关性分析,并根据盖钧镒等[9]提出的主基因+多基因遗传分析对F2代分离群体进行分析,得到各模型的极大似然值及值,筛选出最适模型,并估算遗传参数,得到各性状的广义遗传率。
2 结果与分析
2.1 各农艺性状的相关性分析
由表1可知,叶片数与茎节数、株高、茎粗呈极显著正相关,相关程度最高的是茎节数,相关系数为0.560;茎节数与株高、茎粗呈极显著正相关,相关程度最高的是株高,相关系数为0.441;株高与茎粗呈显著正相关,相关系数为0.111;茎粗与蜡粉呈显著负相关,相关系数为-0.111。
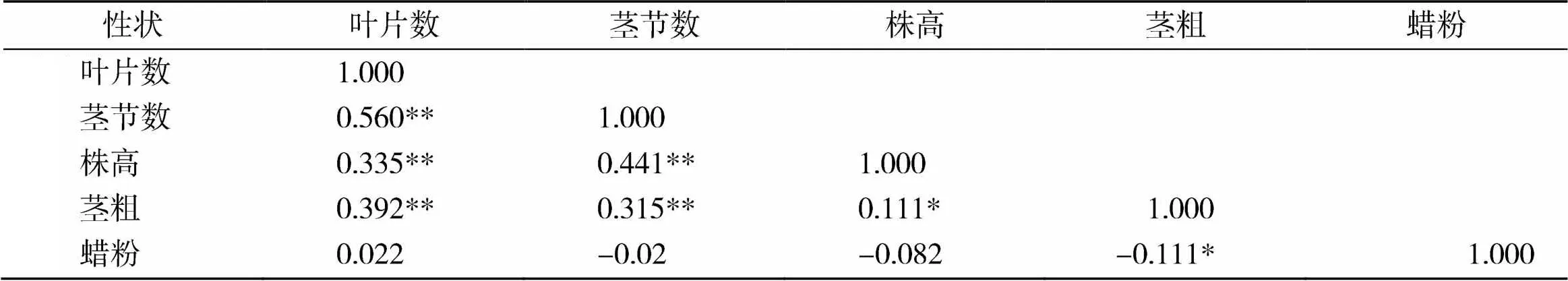
表1 F2群体各农艺性状间的相关系数
注:**在0.01水平(双侧)上极显著相关;*在0.05水平(双侧)上显著相关
2.2 各农艺性状的表型分析
由图1和表2可知,茎节数、株高和蜡粉均呈偏态单峰分布,符合数量性状遗传的特点,说明这些性状可能由主基因或多基因控制,而叶片数和茎粗呈标准正态分布,说明这些性状可能不存在主基因遗传,而受多基因遗传控制。
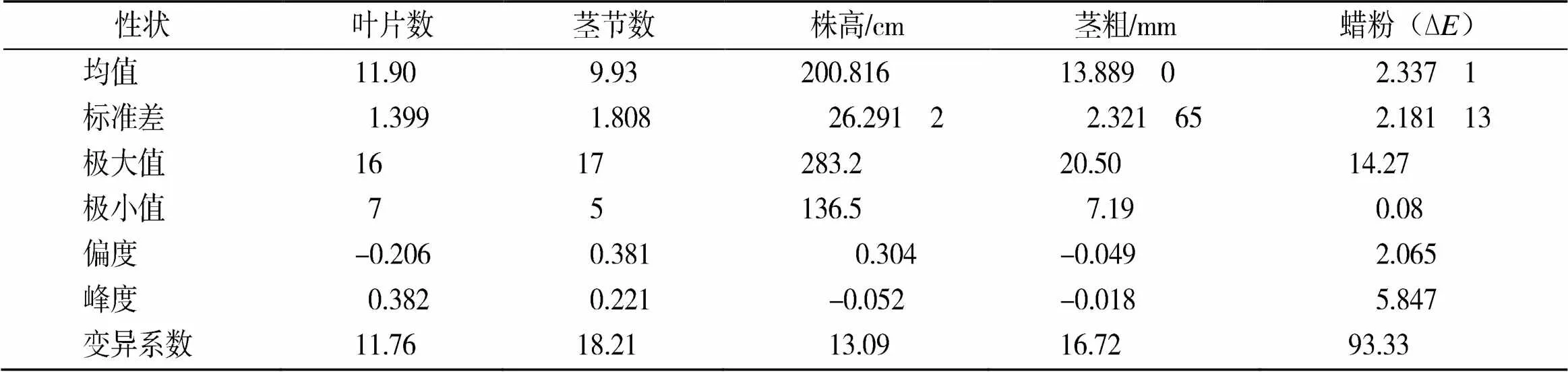
表2 F2群体各农艺性状的表型分析结果
2.3 遗传模型的选择分析
根据盖钧镒等[9-11]的主基因+多基因混合遗传模型的F2代单世代分离分析方法,对叶片数、茎节数、株高、茎粗、蜡粉5个性状进行遗传分析(表3)。首先根据值最小原则,选出值最小的4个模型作为该性状的备选模型,并对其进行适合性检验,以确定各性状的数量遗传模型。叶片数和茎粗的备选模型为ModelA_0、ModelA_2、ModelB_4和ModelB_6;茎节数和株高的备选模型为ModelA_0、ModelA_1、ModelA_4和ModelB_2;蜡粉的备选模型为ModelA_1、ModelA_4、ModelB_1和ModelB_2。
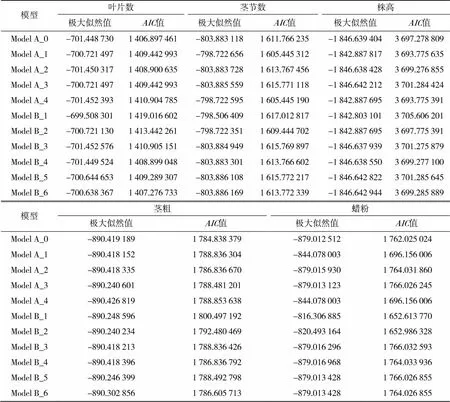
表3 叶片数、茎节数及株高的极大似然值和AIC值
2.4 备选模型的检验分析
根据12、22、32(均匀性检验)、2(Smirnov检验)、(Kolmogorov检验),对叶片数、茎节数、株高、茎粗和蜡粉5个农艺性状的4个备选模型进行适合性检验(表4),选出5个农艺性状的最适遗传模型。
(1)在叶片数4个遗传备选模型中,12、22、32、2和均未达到显著水平(<0.05),且ModelA_0的值最小,因此选择ModelA_0为叶片数的最适遗传模型。说明在该群体中叶片数不受主基因控制,而受微效多基因控制。(2)在茎节数4个遗传备选模型中,12、22、32、2和均未达到显著水平(<0.05),且ModelA_4的值最小,因此选择ModelA_4为茎节数的最适遗传模型。说明在该群体中,茎节数为负向完全显性混合遗传模型,受一对主基因控制。(3)在株高4个遗传备选模型中,除ModelA_0中2未达到显著水平(<0.05)外,2和其余模型均达到显著水平(<0.05),而12、22、32均未达到显著水平(<0.05),且ModelA_4的值最小,因此选择ModelA_4为株高的最适遗传模型。说明在该群体中,株高为负向完全显性混合遗传模型,受一对主基因控制。(4)在茎粗4个遗传备选模型中,2和均达显著水平 (<0.05),而12、22、32均未达到显著水平(<0.05),且相较于其他3个模型,ModelA_0的值最小,因此选择ModelA_0为茎粗的最适遗传模型。说明在该群体中,茎粗不受主基因控制,而受微效多基因控制。(5)在蜡粉4个遗传备选模型中,12、22、2和均未达到显著水平(<0.05),而ModelA_1和ModelA_4中32均达到显著水平(<0.05),且相较于其他3个模型,ModelB_1的值最小,因此选择ModelB_1为蜡粉的最适遗传模型。说明在该群体中,蜡粉由两对主基因控制的加性-显性-上位性混合遗传模型。
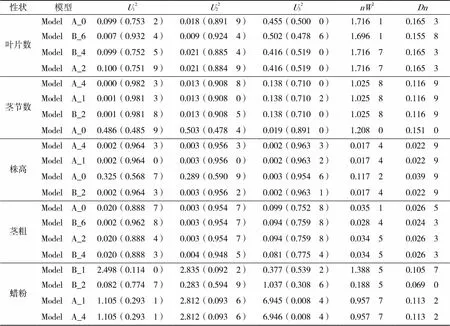
表4 各性状备选模型的适合性检验
2.5 农艺性状遗传参数估计
根据已确定的各个性状最优模型和IECM[12]估算方法,对F2群体农艺性状进行遗传参数估计,结果见表5。由于叶片数和茎粗都符合ModelA_0模型,不存在控制性状遗传的主基因,属于微效多基因控制的数量性状遗传,因此无法计算主基因遗传率。
在高丹草F2代群体中,茎节数的数量遗传模型符合ModelA_4,F2分离群体受一对主基因控制,为负向完全显性模型,即∶(+)=3∶1,主基因的加性效应为1.992 684,说明控制株高的主基因以加性效应为主,主基因遗传率为91.13%,遗传率较高,表明该性状受环境影响较小,可以在后代中稳定遗传;株高的数量遗传模型符合ModelA_4,F2分离群体受一对主基因控制,为负向完全显性模型,即∶(+)=3∶1,主基因的加性效应为24.689251,说明控制株高的主基因以加性效应为主,主基因遗传率为66.14%,遗传率较高,说明该性状受环境影响较小,可以在后代中稳定遗传;蜡粉符合ModelB_1模型,说明该群体中蜡粉符合两对主基因遗传的加性-显性-上位性混合遗传模型,两对主基因的加性效应分别为1.5827675和1.5816405,显性效应分别为-1.582167 0和-1.572 800 5,加性效应之和显著大于显性效应之和(d+d>h+h),即蜡粉的遗传是以两对主基因的加性效应为主,两对主基因的遗传率为54.03%,该群体蜡粉的遗传率较高,说明蜡粉在后代遗传中受环境因素影响较小。
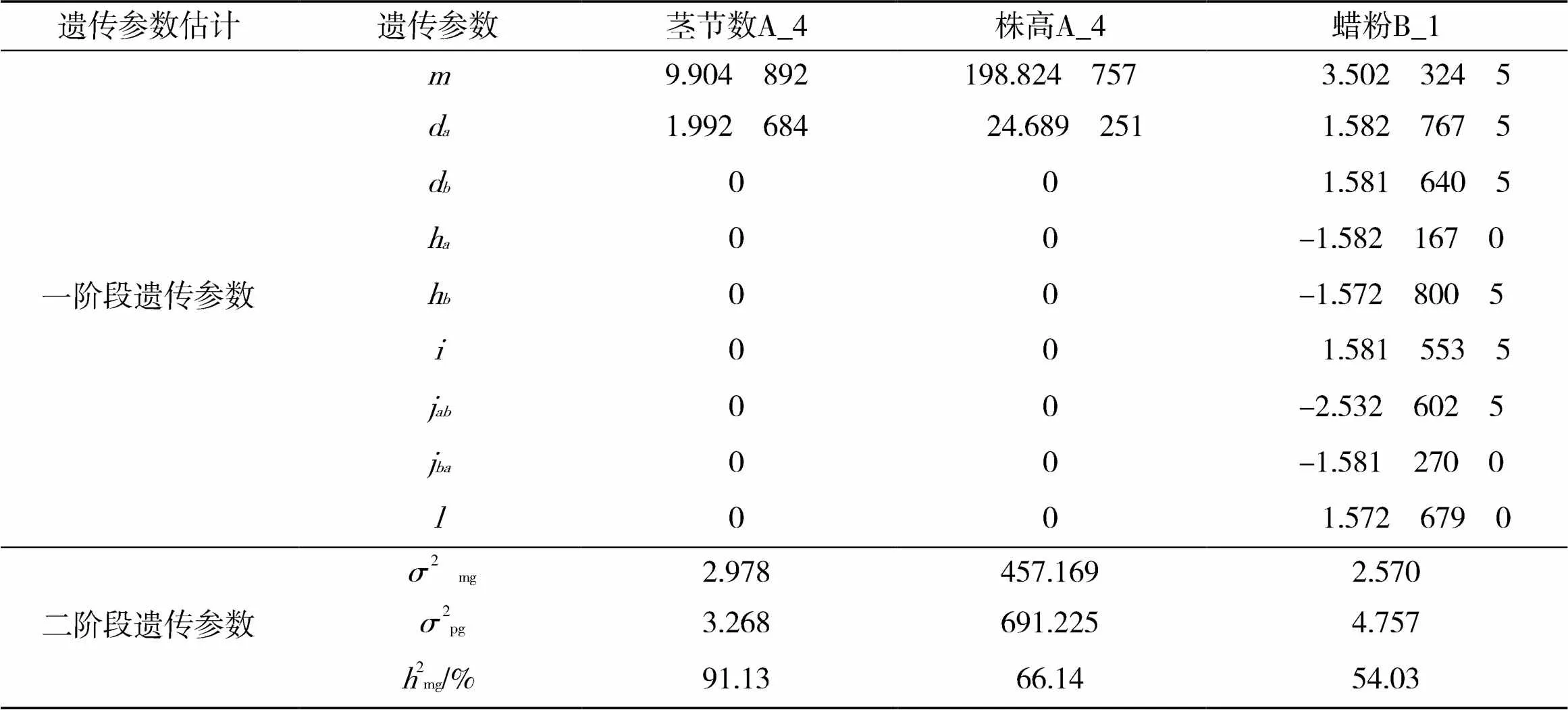
表5 各性状最适遗传模型的遗传参数估计
注:为群体均方;d为主基因的加性效应;d为主基因的加性效应;h为主基因的显性效应;h为主基因的显性效应;为2个主基因的加×加效应;j为加性()×显性()效应;j为加性()×显性()效应;为2个主基因的显×显效应;2mg为遗传方差;2pg为表型方差;2mg为主基因遗传率
3 结论
本研究通过主基因+多基因分析对高丹草F2代的叶片数、茎节数、株高、茎粗和蜡粉5个农艺性状进行遗传分析。结果表明:叶片数符合ModelA_0模型,该性状不存在主基因,为微效多基因控制,因此无法估算主基因遗传率,这与白晓倩等[13]的研究结果(ModelA_4)不一致,分析原因可能是试验群体种植的环境不同,由环境因素影响所造成的表型数据有差异;茎节数符合ModelA_4模型,为负向完全显性混合遗传模型,受一对主基因遗传,主基因遗传率为91.13%,这与白晓倩等[14]在高粱F2代中的研究结果(ModelA_1)不一致,造成结果不一致的原因可能是所选群体不一致,也可能受种植环境的影响;株高符合ModelA_4模型,为负向完全显性混合遗传模型,受一对主基因遗传,主基因遗传率为66.14%,这与白晓倩等[15]在高粱和苏丹草杂交F2代群体中的结果一致;茎粗符合ModelA_0模型,该性状不存在主基因,为微效多基因控制,因此无法估算主基因遗传率,这与卢华雨等[16]的研究结果(ModelB_6)不一致,其原因可能是群体大小有所差异;蜡粉符合ModelB_1模型,由两对主基因控制的加性-显性-上位性混合遗传模型,主基因遗传率为54.03%。
通过对‘忻粱52’和苏丹草‘美引-48’杂交F2代群体叶片性状的主基因+多基因分析,得到各性状的最适遗传模型及广义遗传率。在5个农艺性状中,茎节数、株高和蜡粉的遗传率相对较高,说明这3个性状的遗传受环境因素影响较小,可在育种早代进行选育。但大部分研究结果与前人研究有一定差异,说明数量性状不仅受遗传因子的控制,外界环境因素也对其遗传有较大影响。因此,在以后对杂交后代的遗传研究中,尤其应注意考虑环境对各性状的影响。
[1]杨小翠,詹秋文,刘言龙,等.苏丹草与高粱的判别分析和聚类分析[J].中国草地学报,2016,38(1):41-46.
[2]汪建飞,段立珍,罗自琴.杂交苏丹草中CN-含量的测定[J].草业学报,2002,11(1):43-46.
[3]ANTONOPOULOU G,GAVALA H N,SKIADAS I V,et al.Biofuels generation from sweet sorghum:Fermentative hydrogen production and anaerobic digestion of the remaining biomass[J].Bioresource Technology,2008,99(1):110-119.
[4]ZHAO Y L,DOLAT A,STEINBERGER Y,et al. Biomass yield and changes in chemical composition of sweet sorghum cultivars grown for biofuel[J]. Field Crops Research, 2009,111(1):55-64.
[5]穆艳芳.不同栽培措施对高粱品质性状的影响[J].天津农业科学,2016,22(2):106-108.
[6]GAN L,WANG X L,CHENG Z J,et al.encoding a β-ketoacyl-CoA reductase is involved in cuticular wax biosynthesis in rice[J]. Plant Cell Reports,2016,35(8):1687-1698.
[7]LEE S B,SUH M C. Advances in the understanding of cuticular waxes inand crop species[J].Plant Cell Reports,2015,34(4):557-572.
[8]陆虎华,胡加如,薛林,等.9个糯玉米自交系主要品质性状的配合力效应及遗传参数分析[J].天津农学院学报,2011,18(3):1-4.
[9]盖钧镒,王建康.利用回交或F2∶3家系世代鉴定数量性状主基因-多基因混合遗传模型[J].作物学报,1998,24(4):402-409.
[10]盖钧镒,章元明,王建康.QTL混合遗传模型扩展至2对主基因+多基因时的多世代联合分析[J].作物学报,2000,26(4):385-391.
[11]盖钧镒.植物数量性状遗传体系的分离分析方法研究[J].遗传,2005,27(1):130-136.
[12]盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003.
[13]白晓倩,于澎湃,卢华雨,等.高丹草叶部性状数量遗传分析[J].分子植物育种,2019,17(14):4808-4816.
[14]白晓倩,于澎湃,李延玲,等.粒用高粱F2群体农艺性状数量遗传分析[J].华北农学报,2019,34(1):107-114.
[15]白晓倩,于澎湃,卢华雨,等.高丹草主要株型性状遗传分析[J].分子植物育种,2019,17(9):2956-2964.
[16]卢华雨,白晓倩,于澎湃,等.饲用高粱4个主要株型性状的遗传分析[J].贵州农业科学,2019,47(1):5-9.
Genetic analysis of agronomic characters in F2population of×
Dong Linjie, Bao Shuguang, Kang Chen, Xie Xiaodong, Luo FengCorresponding Author, Sun ShoujunCorresponding Author
(College of Agronomy and Resource Environment, Tianjin Agricultural University, Tianjin 300392, China)
In this study, the F2generation population constructed by the cross between the grain‘Xinliang 52’ and‘Meiyin-48’ was used as the test material. Five agronomic characteristics of the F2generation including leaf numbers, stem numbers, plant height, stem diameter and wax powder were measured, and the optimal genetic model and heritability of each trait were obtained by principal-multiple gene analysis. The results showed that the most suitable model for the number of leaves and stem diameter was Model A_0. There was no major gene inheritance, but the quantitative trait inheritance controlled by micro-effect and multi-gene. The optimal model of stem numbers and plant height was Model A_4, a negative full-dominant model controlled by a pair of major genes, and the major gene heritability were 91.13% and 66.14%. The optimal model of wax powder was Model B_1, which belonged to the additive-dominant-epistatic mixed genetic model, controlled by two pairs of major genes with the heritability of the major gene of 54.03%.
×; agronomic characters; major-multi-gene genetic analysis; quantitative inheritance
1008-5394(2021)03-0006-06
10.19640/j.cnki.jtau.2021.03.002
S514
A
2020-11-11
农业农村部“引进国际先进农业科学技术”项目(2016-X09)
董林洁(1995-),女,硕士在读,主要从事饲用作物遗传改良研究。E-mail:donglj25@qq.com。
孙守钧(1961-),男,教授,博士,主要从事饲用作物遗传改良研究。E-mail:sunshoujun@tjau.edu.cn。
罗峰(1980-),男,副教授,硕士,主要从事饲用作物遗传改良研究。E-mail:luofeng1868@126.com。
责任编辑:宗淑萍
猜你喜欢
河北科技师范学院学报(2020年1期)2020-07-02 03:37:22
消费导刊(2017年24期)2018-01-31 01:29:31
华北农学报(2017年2期)2017-06-28 12:37:41
辽宁大学学报(哲学社会科学版)(2017年3期)2017-06-21 21:16:59
作文周刊(高考版)(2016年16期)2017-06-01 15:41:00
党建文汇·下(2016年5期)2016-05-14 11:34:41
奥秘(2016年4期)2016-04-21 17:31:28
造纸化学品(2016年5期)2016-02-07 03:36:57
中学语文(2015年27期)2015-03-01 03:53:28
植物营养与肥料学报(2014年1期)2014-03-11 20:25:15
