
黔中喀斯特9种木质藤本叶功能性状研究
2021-10-13 11:17王梦洁容丽李婷婷王琪叶天木
热带亚热带植物学报 2021年5期
王梦洁, 容丽*, 李婷婷, 王琪, 叶天木
黔中喀斯特9种木质藤本叶功能性状研究
王梦洁1,2, 容丽1,2*, 李婷婷1,2, 王琪1,2, 叶天木1,2
(1. 贵州师范大学地理与环境科学学院(喀斯特研究院),贵阳 550025;2. 中国科学院普定喀斯特生态系统观测研究站,贵州 普定 562100)
为揭示喀斯特生境中藤本植物的生态策略,对中国科学院普定喀斯特生态系统观测研究站的陈旗流域中9种木质藤本的叶片功能性状及其相关性进行了研究。结果表明,叶面积、叶厚度、叶绿素含量、比叶面积、叶组织密度和叶干物质含量6个叶功能性状均存在不同程度的变异,性状的种间变异为9.24%~98.18%,种内变异为0.64%~39.71%。落叶植物性状的种内变异系数低于常绿植物;相较于喀斯特乔灌木,藤本植物具有较低的叶厚度、叶组织密度和较高的叶面积、叶干物质含量;藤本植物叶功能性状间关系紧密,比叶面积与叶组织密度、叶干物质含量呈极显著负相关,叶组织密度与叶干物质含量呈极显著正相关。叶绿素含量与叶干物质含量呈极显著负相关。叶厚度与叶绿素含量呈显著正相关,与叶组织密度呈显著负相关。除叶厚度外,叶面积与其余性状均存在显著或极显著相关性。结合叶经济谱理论分析,刺葡萄(var.)、野葛()和野葡萄()倾向于“快速偿还”型,白木通(、红毛悬钩子()、小果蔷薇()和崖豆藤()倾向于“慢速偿还”型,插田泡()和勾儿茶()的功能分异明显,介于两者之间。可见,为适应干旱贫瘠的喀斯特环境,藤本植物通过权衡叶功能性状关系来采取不同的适应策略。
喀斯特;木质藤本;叶片;功能性状;变异;叶经济谱
藤本植物是一类不能自由直立,必须依附于支持物攀援的层间植物[1–2],主要分布于热带、亚热带地区。由于受支持物的限制,藤本植物受环境因子的影响,形成了独特的形态和适应特征[3]。该类植物通常在林下或林冠攀援、匍匐,与共存树木进行地上和地下的资源竞争,进而影响植物功能性状[1]。
叶片是植物碳收支和水分平衡的主要场所,且对环境变化有高度敏感性,而叶片功能性状与植物对资源的获取及利用密切相关[4],对阐明植物的环境适应性具有重要意义。叶厚度与植物资源获取、水分保存及同化有关[5]。叶绿素含量反映了植物对光的获取能力[6]。比叶面积和叶干物质含量综合反映植物利用资源的能力,处于贫瘠干旱区的植物通过减小比叶面积,增加叶干物质含量,降低植物内部水分散失,提高对养分元素的保有能力[7]。叶组织密度则反映叶片中生物量的累积状况[8]。这些性状具有易测定、可塑性强的特点,对于种间比较和综合反映植物对环境的适应能力的效果明显[9]。
叶经济谱是通过一系列功能性状指标的组合来衡量,比叶面积、叶绿素含量、叶干物质含量等特征是植物资源利用分类轴划分上的最佳变量之一[10]。在叶经济谱的一端是“快速偿还”型(quick- return type),植物具有比叶面积高、叶绿素含量高、叶干物质含量低、寿命短、光合速率高等特征,另一端是“慢速偿还”型(slow-return type),具有比叶面积低、叶绿素含量低、叶干物质含量高、寿命长、光合速率低等特征[11]。植物经济谱数据库已涵盖了全球大部分地区,但在国内研究仍偏少,尤其是关于特殊环境的研究。
植物功能性状在种间、种内的变异是群落构建、物种共存的重要前提[12–13]。通常认为种间变异对群落构建的贡献更大,然而大量研究表明,种内变异同样不能忽略。唐青青等[13]的研究表明植物功能性状的变异有34.6%来源于种内差异,郭志文等[14]的研究表明植物的形态结构性状变异主要来源于物种个体水平(高于50%)。因此,只有结合种内和种间性状变异才可能真实反映物种在群落构建过程中对环境变化和资源竞争的响应[15]。我国喀斯特多分布于亚热带地区,暖湿气候与高度异质性生境演化出了复杂多样的生物种类,孕育了丰富的藤本植物资源[16–17],南方喀斯特地区共有藤本植物1 463种, 隶属67科241属[18]。由于该类植物具有适应性强、生长速度快,覆盖面积较大等独特优势,可作为生态脆弱区植被恢复和重建的优先选择。目前关于喀斯特植物功能性状的研究大多集中在乔灌木[19],涉及层间藤本植物的偏少,且对于喀斯特藤本的研究以物种组成或者从光合生理角度探讨其对干旱胁迫的适应性等为主[17,20],缺乏叶片结构性状等对环境的适应性研究。为此,本文选取贵州省中部普定县典型喀斯特地区的藤本植物为研究对象,通过测定9种藤本植物的叶片性状,分析喀斯特地区藤本植物叶性状特征、种间种内变异程度以及叶片性状间的关系,揭示该地区藤本植物对生境的适应性特征,以期为喀斯特植被恢复和石漠化治理提供科学依据。
1 材料和方法
1.1 研究区概况
研究样地位于中国科学院普定喀斯特生态系统观测研究站监测研究区的陈旗流域(105°43ʹ30ʹʹ~ 105°44ʹ43ʹʹ E,26°15ʹ36ʹʹ~ 26°15ʹ56ʹʹ N),地跨陈旗堡和赵家田两村。平均海拔为1 403 m。该地年均气温15.2℃,1月均温5.2℃,7月均温23.0℃,年均降雨量1 341 mm,集中在5-9月。年均相对湿度80%,年日照时数仅1 189 h,属于中亚热带季风温暖湿润气候区。以石灰土为主,黄壤次之。土壤偏中性,有效磷(6.27 mg/kg)和有机质含量(10.67%)偏低,而有效氮(440.53 mg/kg)含量偏高。受人类活动影响,植被类型仅以次生性、次顶级的常绿落叶阔叶混交林存在,藤刺灌丛和灌草丛则占据绝对优势。该地区群落层间植物十分丰富,主要物种有白木通()、勾儿茶()、崖豆藤()、野葛()、小果蔷薇()、插田泡()等。该类植物生态适应性强,覆盖范围较广,部分植物如野葛、野葡萄的高度可达10 m以上,对于改善喀斯特脆弱的生态环境具有重要意义,且这些植物有着较高的经济价值,如白木通、野葛、野葡萄等可作为特种经济植物。
1.2 功能性状的选取和测定
2019年8-10月,在研究区(面积约1 hm2)内设置6个面积为30 m×20 m的样地,位置处于山地的中坡且以半阳坡面为主,样地间隔距离较小。采用典型取样法进行调查,乔木样方面积为20 m×20 m,灌木样方面积为5 m×5 m,调查并记录样方中乔木、灌木及藤本的名称、株数、株高、冠幅、胸径/基径等相关指标。
根据群落调查的结果,对每个样地中的藤本优势种进行采集(表1)。每种随机采取生长良好、个体大小一致的5株作为研究对象。在每株外冠层的东南西北4个方向,采集完全展开、未受病虫害的成熟叶片(每株不少于5片),分别用便捷式叶面积扫描仪AM-300和便携式叶绿素仪CCM-200测定叶面积和叶绿素含量。选用数显千分尺(精确度为0.001 mm)测量叶厚度,测量时应尽量避开主叶脉,一次测量5枚叶片厚度(LT),重复3次取平均值。用电子天秤称量叶片鲜重后放入烘箱,105℃杀青20 min,之后在80℃烘箱内烘48 h,取出后称其干质量。参照Cornelissen等[21]的方法计算叶片功能性状,比叶面积(SLA)=叶面积(cm2)/叶干质量(g);叶干物质含量(LDMC)=叶干重(g)/叶鲜重(g);叶组织密度(LTD)=叶干质量(g)/叶体积(cm3);叶体积=叶面积(cm2)×叶厚度(cm)。
1.3 数据统计与分析
首先对植物功能性状数据进行初步整理,对偏离正态分布的数据进行对数转换,计算各功能性状的变异系数(CV),通过方差分析比较物种性状间的差异,采用Personal相关性检验分析植物功能性状间的相关性,利用主成分分析(PCA)对不同物种叶片功能性状进行主成分排序分析。数据分析与图表绘制均在SPSS 23、Origin 2018和CANOCO 4.5中完成。
2 结果和分析
2.1 功能性状的变异
植物叶片功能性状在种内和种间水平上均存在不同程度的变异。由图1可见,从种内变异系数来看,变异系数最大的分别为:插田泡的叶面积(25.32%)、野葛的叶厚度(36.33%)、野葡萄的叶绿素含量(5.90%)、刺葡萄的比叶面积(25.69%)、勾儿茶的叶组织密度(39.71%)、红毛悬钩子的叶干物质含量(5.87%);变异系数最小的分别为:小果蔷薇的叶面积(9.02%)和叶厚度(9.29%)、红毛悬钩子的叶绿素含量(1.91%)、野葛的比叶面积(6.51%)、插田泡的叶组织密度(6.97%)、勾儿茶的叶干物质含量(0.64%)。从植物的生活型来看,落叶植物叶片功能性状的种内变异系数普遍低于常绿植物。叶片性状的种间变异系数为9.24%~98.18%,表现为叶绿素含量<干物质含量<厚度<比叶面积<组织密度<面积。
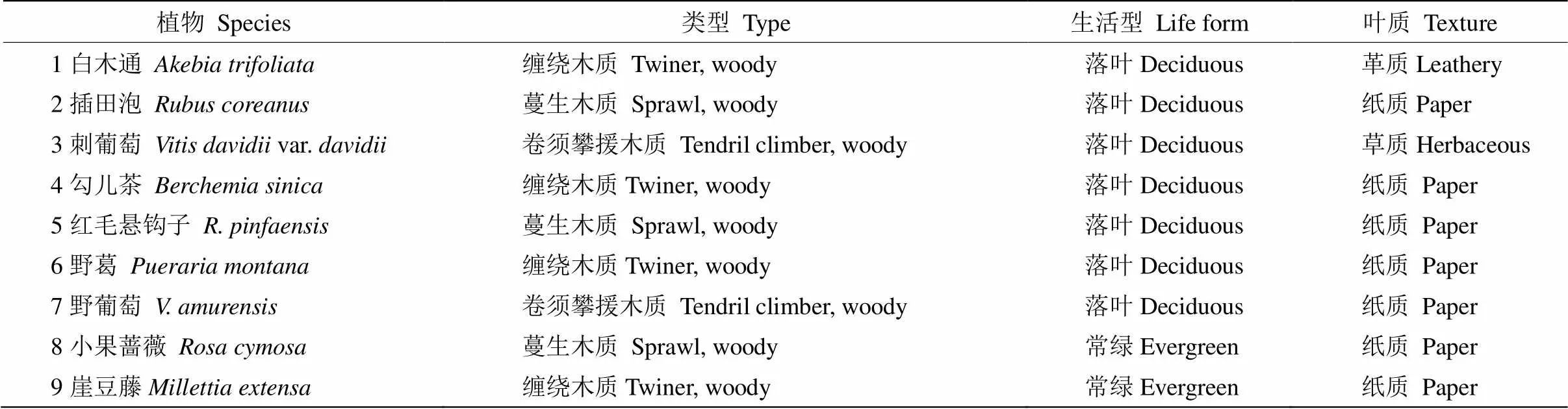
表1 9种优势藤本植物
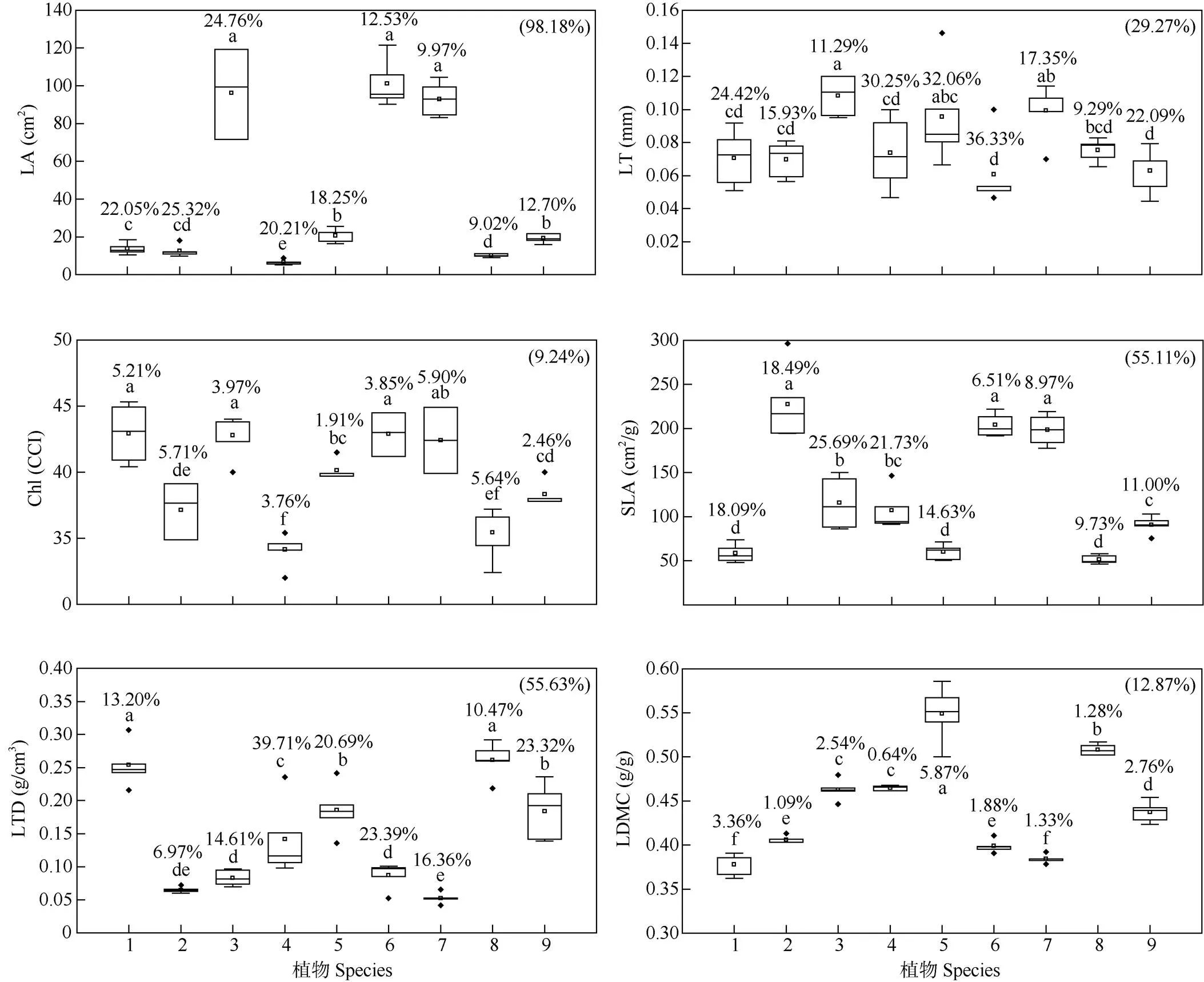
图1 9种优势藤本植物的叶片功能性状。□: 平均值; ◆: 异常值; 不同字母表示差异显著(P<0.05); 字母上方的数据为种内变异系数,括号内数据为种间变异系数; LA: 叶面积; LT: 叶厚度; Chl: 叶绿素含量; SLA: 比叶面积; LTD: 叶组织密度; LDMC: 叶干物质含量。1~9见表1。以下图表同。
在物种水平上,同一性状在不同植物间均存在差异,且不同性状具有不同的权衡。小果蔷薇的比叶面积最小,而叶干物质含量较大,野葡萄的叶组织密度最小,而叶厚度偏大。插田泡的叶组织密度较小,而比叶面积最大,白木通的叶干物质含量最小,而叶绿素含量最大。这表明植物通过不同性状的组合来适应复杂多变的喀斯特生境。
2.2 叶性状间的相关性
通过对喀斯特地区9种藤本植物叶片性状进行Pearson相关分析(表2),结果表明,叶面积与叶绿素含量、比叶面积呈极显著正相关(<0.01),与叶干物质含量呈显著负相关(<0.05),叶厚度与叶绿素含量呈显著正相关(<0.05),与叶组织密度呈显著负相关(<0.05),叶组织密度与叶面积、比叶面积呈极显著负相关(<0.01),与叶干物质含量呈极显著正相关(<0.01),叶干物质含量与叶绿素含量、比叶面积呈极显著负相关(<0.01)。
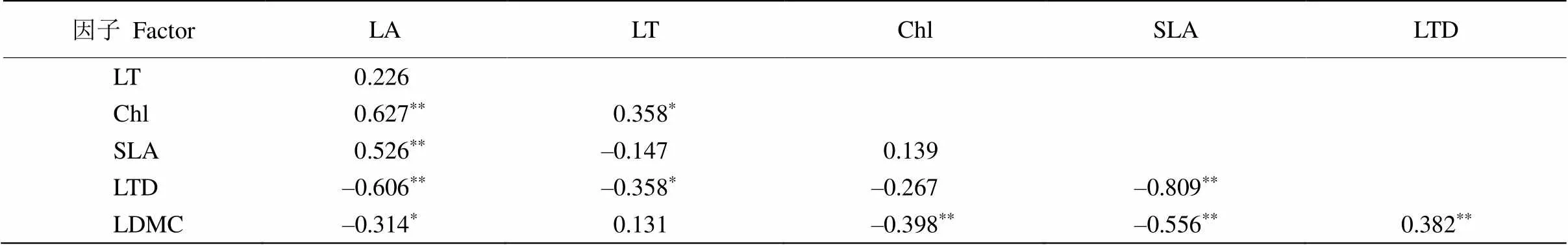
表2 叶片功能性状的Pearson相关性
**:<0.01; *:<0.05.
2.3 主成分分析
主成分分析(PCA)结果表明(图2),第一排序轴解释的物种变异为84.7%,第二排序轴为15.0%,两者累积解释率达99.7%,包含了原始变量的绝大部分信息。第一轴与叶面积、叶厚度以及叶绿素含量呈正相关关系,与叶组织密度呈负相关关系。第二轴与比叶面积呈正相关关系,与叶干物质含量呈负相关关系。
在物种-性状排序图中可发现,6个叶功能性状指标发生明显的趋异分化现象,9种植物被明显地划分为2类功能类群。刺葡萄、野葛和野葡萄植物位于第一主成分轴的正向区域,具有较高的叶面积、叶厚度、叶绿素含量和比叶面积,白木通、插田泡、勾儿茶、红毛悬钩子、小果蔷薇和崖豆藤植物位于第一主成分轴的负向区域,具有较高的叶组织密度和叶干物质含量。其中,插田泡和勾儿茶分布在排序轴的外围,性状趋异分化较明显。插田泡的比叶面积最大,勾儿茶的叶面积和叶绿素含量最小。
进一步分析表明(表3),刺葡萄、野葛、野葡萄植物总体倾向于“快速偿还”型,其叶面积、叶绿素含量和比叶面积显著高于白木通、红毛悬钩子、小果蔷薇和崖豆藤倾向于“慢速偿还”型的植物,但叶组织密度显著较低。而插田泡和勾儿茶的叶面积和叶绿素含量小、比叶面积大,叶组织密度和叶干物质含量介于“快速偿还”型和“慢速偿还”型之间,因此该类植物更倾向于“中间”型。可见,植物对资源的分配与投资策略具有差异性。
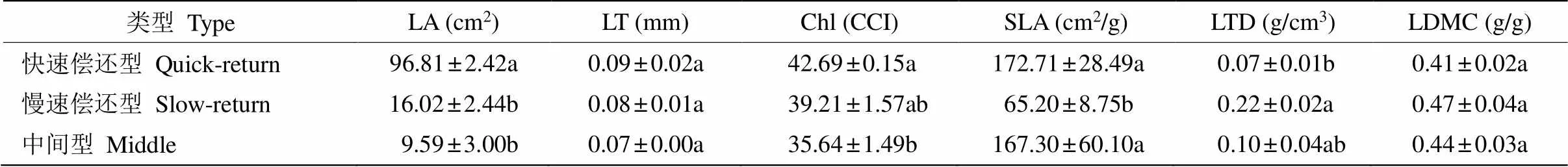
表3 不同生活策略植物性状特征
同列数据后不同字母表示差异显著(<0.05)。
Data followed different letters within column indicate significant difference at 0.05 level.
3 结论和讨论
3.1 喀斯特藤本植物叶性状的变异特征
本研究结果表明,黔中9种藤本植物叶功能性状在种间和种内均存在不同程度的变异,说明不同物种及性状对环境胁迫的适应策略具有明显差异。藤本植物的种内变异平均为13.27%,低于黔中恢复较好的次生林木本植物(21.4%)[22]和粤东常见植物(76.4%)[23]。在相同环境下,藤本植物擅长攀援,比树木的截光能力更强,能够充分获得阳光进行光合作用[1],光资源获取量高,竞争小,且本研究区群落演替属于中前期,藤本生物量较高,性状可塑性低,变异幅度较小[24]。与气候、土壤等自然条件优越的粤东地区相比,黔中喀斯特地区气候寒凉,岩石裸露率达43.27%,石灰岩抗蚀性和抗冲性强,石漠化发育强烈。藤本植物虽拥有发达的根系,但石生环境制约了根系的垂直深入,可能使得部分根系暴露于地面,无法扎入岩土深部[25–26],生长受到抑制,降低了性状可塑性。植物性状主要受遗传因素和环境的影响,具有不同遗传背景的物种在性状方面存在较大种间差异[4]。本研究中,藤本植物的种间变异平均为43.38%,高于种内变异水平。其中,叶面积的种间变异系数(98.18%)最大。独特的喀斯特地貌孕育着丰富的生物多样性,其物种遗传的多样性对碳和捕获光的潜在能力强。如部分藤本植物长期未找到支持物,可能会以横向扩散方式即产生侧根并增加叶面积来满足光能需求[27]。叶绿素含量(9.24%)和叶干物质含量变异系数(12.87%)较小,说明叶绿素含量、叶干物质含量是资源获取轴上较稳定的变量,藤本植物随贫瘠干旱环境的影响而发生趋同效应。研究区落叶藤本植物各性状的种内平均变异水平低于常绿,与前人的研究结果一致[13,22]。本区域海拔较高,冬季气温偏低且喀斯特环境所带来的季节性干旱使得植物为避免低温和水分胁迫,通过落叶方式缩短生长期,季节变化小,轻微的环境扰动对其影响较小,诱导叶性状发生变异的程度低[28]。
3.2 喀斯特藤本植物叶片特征
喀斯特地区土层浅薄,土壤养分总量少,再加上临时性干旱,植物多以较小的叶面积、比叶面积和较大的叶厚度、叶组织密度以及叶干物质含量的性状组合。位于同一生态站的天龙山与陈旗流域的距离较近,土壤等环境条件相对一致。与该研究区的乔灌树木[22]相比,藤本植物的叶厚度(0.08 mm)和叶组织密度(0.15g/cm3)较低,而叶面积(41.52 cm2)和叶干物质含量(0.44g/g)较高。这可能是为了争夺阳光,藤本植物通过减少叶厚度碳构建而增加叶片面积碳投资[29]。密度大的叶片可能会增加CO2在叶肉组织中传导的阻力[30–31],植物减少叶组织密度来扩大光在叶肉组织中的传播范围,并将其合成的干物质作用于对环境抗逆性的提高。因此,相较于喀斯特树木而言,藤本植物表现出较低的叶厚度、叶组织密度和较高的叶面积、叶干物质含量等性状特征。
3.3 喀斯特藤本植物叶性状间的权衡关系明显
本研究结果表明,比叶面积与叶组织密度、叶干物质含量呈极显著负相关,叶组织密度与叶干物质含量呈极显著正相关,与琅岐岛4种优势植物和桂西北主要木本经济植物的研究结果相同[32–33]。与其他地区的藤本植物[31]相比,本研究区的比叶面积(123.73 cm2/g)相对较小,意味着植物对叶片的构建成本较高,对光合器官的投资较低。侯皓等[34]研究表明,落叶物种的叶片构建成本小于常绿物种,采取提高比叶面积等的低成本构建策略。本文研究对象多为落叶物种,需在较短时间内积累能量为越冬和次年的生长做准备,但由于喀斯特山区水土匮乏,该类植物通过减小比叶面积来降低光合作用和蒸腾作用的成本[35],同时从根系吸收的养分多用于构建叶片防御结构,使得单位面积叶片分配到更多的生物量,以此提高对不良生境的耐受性[36–37],这可能是对喀斯特环境的一种适应策略。本研究的叶片叶绿素含量与叶干物质含量呈极显著负相关,说明藤本植物采用降低光合作用能力来提高对养分的积累,增强叶片的坚韧性,使其对非生物胁迫的抵抗力更强[22]。本研究中叶厚度与叶绿素含量呈显著正相关,与滇中喀斯特41种不同生长型植物的研究结论一致[38]。叶厚度与叶组织密度呈显著负相关,藤本植物为适应层间荫蔽环境减少叶片厚度而增加叶肉密度,加强在微弱光线下的光合作用能力。叶片是植物固碳的主要部位,面积直接影响光截取和碳获取的能力[39],因此叶面积与其他性状(除叶厚度外)间存在显著或极显著的相关性。本文中的大部分叶性状间关系紧密,表明藤本植物在应对环境变化时,表现出更加灵活多变的适应方式。
3.4 喀斯特藤本植物总体存在叶经济谱现象
植物在漫长进化中,通过形成特定的功能性状属性减少来自外界环境的不利影响,使得不同的物种在叶经济谱的位置不同[39]。本文研究表明,刺葡萄、野葛和野葡萄植物具有较高的叶面积、叶厚度、叶绿素含量和比叶面积以及较低的叶组织密度和叶干物质含量,在叶经济谱中的位置总体靠近“快速偿还”型一端。该类藤本植物趋向于增加叶片的面积与叶绿素含量,扩大对光的吸收范围,维持更好的光合能力。且在荫蔽环境下,部分植物以较低的光补偿点来充分利用弱光。此外,刺葡萄和野葡萄的自身抗旱基因强,可溶性蛋白较高,体内水分散失少[40]。而野葛对土壤几乎无要求且侧根发达, 为叶片光合作用运输大量的水分和养分[41]。刺葡萄、野葡萄和野葛植物对环境适应性较强,偏重于获取更多的营养物质以达到快速生长的目的。由于局部立地土壤条件或样品数量有限等导致叶经济谱中的叶厚度出现相反现象[42],其具体原因仍需进一步研究。
白木通、红毛悬钩子、小果蔷薇和崖豆藤植物总体靠近“慢速偿还”型一端,具有较低的叶面积、叶绿素含量和比叶面积以及较高的叶组织密度和叶干物质含量。研究区土壤有机质、有效磷等养分含量偏少,植物为获取营养物质,可能将更多的能量投入到根系的构建,叶片则采取保守的策略。该类植物的平均叶面积小于20.25 cm2,属于小型叶。研究表明,较低的比叶面积有利于热耗散和气孔对水分的调节[43],减少能量消耗。随干旱的加剧,比叶面积越小,水分利用效率和渗透调节能力越强,生产更多的生物量来抵抗干旱生境[44]。小果蔷薇的比叶面积最小,叶组织密度最大,叶干物质含量较大,对生长的投资缓慢。红毛悬钩子有浓密的表皮毛,能阻碍叶表面空气流动,降低水分蒸发[45]。其叶干物质含量最大,表明抵御外界干扰的能力强[21]。崖豆藤为常绿植物,比叶面积小,比叶质量大,对枝条的机械需求高[22],以较强的木质部抗栓塞能力来维持叶片的功能运行[46]。而白木通叶片为革质叶,水分保持较好,利于干物质的生产和积累。另外,木质藤本导管受水分胁迫时易于空穴化,且随植物生长可能会丧失传导功能[27],再加上喀斯特地区季节性干旱,叶片水分摄入量少,植物可能保持较低的生长速率。
此外,插田泡和勾儿茶的功能性状分化明显且比叶面积较大(167.30 cm2/g),因此单独作为一类进行分析。通过表3可知,该类植物的叶面积和叶绿素小而比叶面积大,叶组织密度和叶干物质含量介于“快速偿还”型和“慢速偿还”型,植物更趋向于两端之间。一方面表明物种在严酷生境中竞争激烈,生态位分化强烈,减少不同物种间的竞争强度[33],另一方面也说明了物种遗传对植物性状的影响,如插田泡的不定根发育扩大了根系对营养物质的获取量,为叶片的光合作用供给充足的养分,通过比叶面积来提高生长速率[4]。勾儿茶没有表皮毛,需降低叶面积和叶绿素含量、增大叶肉密度来避免强光对叶肉细胞的灼伤[21]。
目前,叶经济谱成为研究植物对全球环境变化响应机制的主要方法之一[33]。本研究表明,叶经济谱现象在黔中喀斯特地区同样存在,如刺葡萄、野葛和野葡萄植物靠近“快速偿还”型,白木通、红毛悬钩子、小果蔷薇和崖豆藤植物靠近“慢速偿还”型,插田泡和勾儿茶的性状趋异分化明显,在叶经济谱中介于两者之间。受环境胁迫的影响,植物叶功能性状的相互关系明显,且侧重于对叶片的构建,说明藤本植物在喀斯特地区长期生存和繁衍而采取不同的生存策略。需要说明的是,藤本植物对环境的适应策略在形态结构性状和生理生化性状都有相应的表现。但由于实验条件以及采样的数量有限,未能对生理生化性状进行分析,因此本试验的结论主要基于植物形态结构性状的响应,仅能在一定程度上说明9种藤本植物如何适应喀斯特环境。另外,尽管国内有学者对叶性状和环境因子的关系进行了分析[23,32–33],但叶经济谱与全球变化环境因子的联系研究仍不足,且植物经济谱地上和地下部分是怎样耦合的,地下部分根经济谱是否随环境变化而出现差异,这都是今后需要进一步研究的内容。
[1] SCHNITZER S A, BONGERS F. The ecology of lianas and their role in forests [J]. Trend Ecol Evol, 2002, 17(5): 223–230. doi: 10.1016/ S0169-5347(02)02491-6.
[2] NABE-NIELSEN J. Diversity and distribution of lianas in a neotro- pical rain forest, Yasuni National Park, Ecuador [J]. J Trop Ecol, 2001, 17(1): 1–19. doi: 10.1017/S0266467401001018.
[3] JIANG H, ZHOU G Y, HUANG Y H, et al. Photosynthetic characteri- stics of canopy-dwelling vines in lower subtropical evergreen broad- leaved forest and response to environmental factors [J]. Chin J Plant Ecol, 2011, 35(5): 567–576. doi: 10.3724/SP.J.1258.2011.00567.
江浩, 周国逸, 黄钰辉, 等. 南亚热带常绿阔叶林林冠不同部位藤本植物的光合生理特征及其对环境因子的适应 [J]. 植物生态学报, 2011, 35(5): 567–576. doi: 10.3724/SP.J.1258.2011.00567.
[4] MENG T T, NI J, WANG G H. Plant functional traits, environments and ecosystem functioning [J]. Chin J Plant Ecol, 2007, 31(1): 150–165. doi: 10.17521/cjpe.2007.0019.
孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能 [J]. 植物生态学报, 2007, 31(1): 150–165. doi: 10.17521/cjpe.2007.0019.
[5] Gonzalez-Paleo L, Ravetta D A. Relationship between photo- synthetic rate, water use and leaf structure in desert annual and peren- nial forbs differing in their growth [J]. Photosynthetica, 2018, 56(4): 1177–1187. doi: 10.1007/s11099-018-0810-z.
[6] JIN Y, WANG C K. Trade-offs between plant leaf hydraulic and economic traits [J]. Chin J Plant Ecol, 2015, 39(10): 1021–1032. doi: 10.17521/cjpe.2015.0099.
金鹰, 王传宽. 植物叶片水力与经济性状权衡关系的研究进展 [J]. 植物生态学报, 2015, 39(10): 1021–1032. doi: 10.17521/cjpe.2015. 0099.
[7] ZHAO H Y, LI Y L, WANG X Y, et al. Variations in leaf traits of 52 plants in Horqin Sand Land [J]. J Desert Res, 2010, 30(6): 1292–1298.
赵红洋, 李玉霖, 王新源, 等. 科尔沁沙地52种植物叶片性状变异特征研究 [J]. 中国沙漠, 2010, 30(6): 1292–1298.
[8] REICH P B, WALTERS M B, ELLSWORTH D S, et al. Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: A test across biomes and functional groups [J]. Oecologia, 1998, 114(4): 471–482. doi: 10.1007/s004420050471.
[9] READ Q D, MOORHEAD L C, SWENSON N G, et al. Convergent effects of elevation on functional leaf traits within and among species [J]. Funct Ecol, 2014, 28(1): 37–45. doi: 10.1111/1365-2435.12162.
[10] Swenson N G. The functional ecology and diversity of tropical tree assemblages through space and time: from local to regional and from traits to transcriptomes [J]. Int Sch Res Not, 2012, 2012: 743617. doi: 10.5402/2012/743617.
[11] OSNAS J L D, LICHSTEIN J W, Reich P B, et al. Global leaf trait relationships: Mass, area, and the leaf economics spectrum [J]. Science, 2013, 340(6133): 741–744. doi: 10.1126/science.1231574.
[12] Diaz S, Hodgson J G, Thompson K, et al. The plant traits that drive ecosystems: Evidence from three continents [J]. J Veg Sci, 2004, 15(3): 295–304. doi: 10.1111/j.1654-1103.2004.tb02266.x.
[13] TANG Q Q, HUANG Y T, DING Y, et al. Interspecific and intra- specific variation in functional traits of subtropical evergreen and deci- duous broad-leaved mixed forests [J]. Biodiv Sci, 2016, 24(3): 262– 270. doi: 10.17520/biods.2015200.
唐青青, 黄永涛, 丁易, 等. 亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异 [J]. 生物多样性, 2016, 24(3): 262–270. doi: 10.17520/biods.2015200.
[14] GUO Z W, ZHAO W X, LUO J F, et al. The variation characteristics of plant functional traits among 16 woody plants in subtropical broad- leaved forest at Dagang Mountain [J]. J Fujian Norm Univ (Nat Sci), 2019, 35(1): 82–87. doi: 10.12046/j.issn.1000-5277.2019.01.012.
郭志文, 赵文霞, 罗久富, 等. 大岗山亚热带常绿阔叶林16种木本植物功能性状的变异特征[J]. 福建师范大学学报(自然科学版), 2019, 35(1): 82–87. doi: 10.12046/j.issn.1000-5277.2019.01.012.
[15] Albert C H, Thuiller W, Yoccoz N G, et al. Intraspecific functional variability: Extent, structure and sources of variation [J]. J Ecol, 2010, 98(3): 604–613. doi: 10.1111/j.1365-2745.2010.01651.x.
[16] LI X K, SU Z M, LÜ S H, et al. The spatial pattern of natural vegetation in the karst regions of Guangxi and the ecological signality for ecosystem rehabilitation and reconstruction [J]. J Mount Sci, 2003, 21(2): 129–139. doi: 10.3969/j.issn.1008-2786.2003.02.001.
李先琨, 苏宗明, 吕仕洪, 等. 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义 [J]. 山地学报, 2003, 21(2): 129–139. doi: 10.3969/j.issn.1008-2786.2003.02.001.
[17] WANG D, GOU G Q, SUN Q L, et al. Diversity and exploitation of the wild woody vines resources in Qiannan, Guizhou [J]. Pratacult Sci, 2017, 34(7): 1506–1515. doi: 10.11829/j.issn.1001-0629.2016-0507.
王灯, 苟光前, 孙巧玲, 等. 贵州黔南野生木质藤本植物资源多样性及开发利用 [J]. 草业科学, 2017, 34(7): 1506–1515. doi: 10.11829/j. issn.1001-0629.2016-0507.
[18] WANG Y S, CHEN L J, LI Y H, et al. Analysis of species composition and attribute characteristics of related traits of lianas growing in the karst areas of south China [J]. Pratacult Sci, 2020, 37(1): 126–138. doi: 10.11829/j.issn.1001-0629.2019-0269.
王业社, 陈立军, 黎颖惠, 等. 中国南方喀斯特地区藤本植物物种组成及其相关性状属性特征分析 [J]. 草业科学, 2020, 37(1): 126– 138. doi: 10.11829/j.issn.1001-0629.2019-0269.
[19] YUAN X X, LIU R H, PAN Y F, et al. Comparative study on func- tional traits of trees and shrubs in different slope aspects on karst hills of Guilin [J]. J Guangxi Acad Sci, 2019, 35(4): 253–261. doi: 10. 13657/j.cnki.gxkxyxb.20191129.005.
苑晓霞, 刘润红, 盘远方, 等. 桂林喀斯特石山不同坡向乔灌木功能性状对比研究 [J]. 广西科学院学报, 2019, 35(4): 253–261. doi: 10.13657/j.cnki.gxkxyxb.20191129.005.
[20] ZHOU J L, DENG P Y, WU Y X. Study on the drought resistance of three liana species in their seedling stages [J]. Guizhou For Sci Technol, 2015, 43(2): 23–28.
周杰良, 邓沛怡, 吴易雄. 3种重度石漠化区生态修复藤本植物苗期抗旱性分析 [J]. 贵州林业科技, 2015, 43(2): 23–28.
[21] Cornelissen J H C, Lavorel S, Garnier E, et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide [J]. Aust J Bot, 2003, 51(4): 335–380. doi: 10.1071/ BT02124.
[22] ZHONG Q L, LIU L B, XU X, et al. Variations of plant functional traits and adaptive strategy of woody species in a karst forest of central Guizhou Province, southwestern China [J]. Chin J Plant Ecol, 2018, 42 (5): 562–572. doi: 10.17521/cjpe.2017.0270.
钟巧连, 刘立斌, 许鑫, 等. 黔中喀斯特木本植物功能性状变异及其适应策略 [J]. 植物生态学报, 2018, 42(5): 562–572. doi: 10.17521/ cjpe.2017.0270.
[23] CHEN W, WANG J H, MA R J, et al. Variance in leaf functional traits of 89 species from the eastern Guangdong of China [J]. Chin J Ecol, 2016, 35(8): 2101–2109. doi: 10.13292/j.1000-4890.201608.033.
陈文, 王桔红, 马瑞君, 等. 粤东89种常见植物叶功能性状变异特征[J]. 生态学杂志, 2016, 35(8): 2101–2109. doi: 10.13292/j.1000- 4890.201608.033.
[24] ZHAI S H, WANG P, SHENG L X. Phenotypic plasticity of plant functional traits in competition environments [J]. J Beihua Univ (Nat Sci), 2017, 18(4): 538–546. doi: 10.11713/j.issn.1009-4822.2017.04.026.
翟偲涵, 王平, 盛连喜. 竞争条件下植物功能性状的表型可塑性研究进展 [J]. 北华大学学报(自然科学版), 2017, 18(4): 538–546. doi: 10.11713/j.issn.1009-4822.2017.04.026.
[25] LUO D H, XIA J, YUAN J W, et al. Root biomass of karst vegetation in a mountainous area of southwestern China [J]. Chin J Plant Ecol, 2010, 34(5): 611–618. doi: 10.3773/j.issn.1005-264x.2010.05.015.
罗东辉, 夏婧, 袁婧薇, 等. 我国西南山地喀斯特植被的根系生物量初探 [J]. 植物生态学报, 2010, 34(5): 611–618. doi: 10.3773/j.issn. 1005-264x.2010.05.015.
[26] ZHAO Y Y, ZHOU Y C, DUAN X. Anti-erodibility and anti-scour- ability of different lithological soils in karst area of central Guizhou Province [J]. J Anhui Agric Sci, 2007, 35(29): 9311–9313,9317. doi: 10.3969/j.issn.0517-6611.2007.29.090.
赵洋毅, 周运超, 段旭. 黔中喀斯特地区不同岩性土壤的抗蚀抗冲性研究 [J]. 安徽农业科学, 2007, 35(29): 9311–9313,9317. doi: 10. 3969/j.issn.0517-6611.2007.29.090.
[27] CHEN Y J, CHEN J W, CAI Z Q. Lianas and their functions in tropical forests [J]. Chin Bull Bot, 2007, 24(2): 240–249. doi: 10.3969/j.issn. 1674-3466.2007.02.017.
陈亚军, 陈军文, 蔡志全. 木质藤本及其在热带森林中的生态学功能 [J]. 植物学通报, 2007, 24(2): 240–249. doi: 10.3969/j.issn.1674- 3466.2007.02.017.
[28] KITAJIMA K. Relative importance of photosynthetic traits and allo- cation patterns as correlates of seedling shade tolerance of 13 tropical trees [J]. Oecologia, 1994, 98(3/4): 419–428. doi: 10.1007/BF0032 4232.
[29] MCDOWELL N, POCKMAN W T, ALLEN C D, et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? [J]. New Phytol, 2008, 178 (4): 719–739. doi: 10.1111/j.1469-8137.2008.02436.x.
[30] ZHANG J L, POORTER L, CAO K F. Productive leaf functional traits of Chinese savanna species [J]. Plant Ecol, 2012, 213(9): 1449–1460. doi: 10.1007/s11258-012-0103-8.
[31] DING L Z, CHEN Y J, ZHANG J L. Leaf traits and their associations among liana species in tropical rainforest [J]. Plant Sci J, 2014, 32(4): 362–370. doi: 10.3724/SP.J.1142.2014.40362.
丁凌子, 陈亚军, 张教林. 热带雨林木质藤本植物叶片性状及其关联 [J]. 植物科学学报, 2014, 32(4): 362–370. doi: 10.3724/SP.J.1142. 2014.40362.
[32] ZHANG X F, MU Z B, LIN M J, et al. Functional traits of leaves of four dominant plants on Langqi Island, Fuzhou, and factors influencing these traits [J]. Chin J Appl Environ Biol, 2020, 26(3): 667–673. doi: 10.19675/j.cnki.1006-687x.2019.07058.
张秀芳, 穆振北, 林美娇, 等. 琅岐岛4种优势植物叶功能性状及其影响因子 [J]. 应用与环境生物学报, 2020, 26(3): 667–673. doi: 10.19675/j.cnki.1006-687x.2019.07058.
[33] PANG S L, OU Z Y, SHEN W H, et al. Leaf function traits variations and adaptive strategies of major woody economic plants in karst area of southwest Guangxi [J/OL]. (2020-07-01). doi: 10.11931/guihaia. gxzw202002005.
庞世龙, 欧芷阳, 申文辉, 等. 桂西南喀斯特地区主要木本经济植物叶功能性状变异及其适应策略 [J/OL]. (2020-07-01). doi: 10. 11931/guihaia.gxzw202002005.
[34] HOU H, LIU H, HE P C, et al. Different leaf construction strategies in evergreen and deciduous species of Magnoliaceae [J]. J Trop Subtrop Bot, 2019, 27(3): 272–278. doi: 10.11926/jtsb.3978.
侯皓, 刘慧, 贺鹏程, 等. 木兰科常绿与落叶物种叶片构建策略的差异 [J]. 热带亚热带植物学报, 2019, 27(3): 272–278. doi: 10. 11926/jtsb.3978.
[35] ZHU J Y, YU Q, LIU Y P, et al. Response of plant functional traits and leaf economics spectrum to urban thermal environment [J]. J Beijing For Univ, 2018, 40(9): 72–81. doi: 10.13332/j.1000-1522.20180132.
朱济友, 于强, 刘亚培, 等. 植物功能性状及其叶经济谱对城市热环境的响应 [J]. 北京林业大学学报, 2018, 40(9): 72–81. doi: 10. 13332/j.1000-1522.20180132.
[36] SONG J, LI R H, ZHU S D, et al. Leaf functional traits of ferns from different habitats in monsoon evergreen broad-leaved forest in Dinghu- shan Mountain [J]. J Trop Subtrop Bot, 2013, 21(6): 489–495. doi: 10. 3969/j.issn.1005-3395.2013.06.001.
宋娟, 李荣华, 朱师丹, 等. 鼎湖山季风常绿阔叶林不同生境蕨类植物的叶片功能性状研究 [J]. 热带亚热带植物学报, 2013, 21(6): 489–495. doi: 10.3969/j.issn.1005-3395.2013.06.001.
[37] BACELAR E A, MOUTINHO-PEREIRA J M, GONÇALVES B C, et al. Changes in growth, gas exchange, xylem hydraulic properties and water use efficiency of three olive cultivars under contrasting water availability regimes [J]. Environ Exp Bot, 2007, 60(2): 183–192. doi: 10.1016/j.envexpbot.2006.10.003.
[38] PANG Z Q, LU W L, JIANG L S, et al. Leaf traits of different growing plants in karst area of Shilin, China [J]. Guihaia, 2019, 39(8): 1126– 1138. doi: 10.11931/guihaia.gxzw201810009.
庞志强, 卢炜丽, 姜丽莎, 等. 滇中喀斯特41种不同生长型植物叶性状研究 [J]. 广西植物, 2019, 39(8): 1126–1138. doi: 10.11931/gui haia.gxzw201810009.
[39] YU H Y, CHEN Y T, XU Z Z, et al. Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei Mongol [J]. Chin J Plant Ecol, 2014, 38(10): 1029–1040. doi: 10.3724/SP.J.1258.2014.00097.
于鸿莹, 陈莹婷, 许振柱, 等. 内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析 [J]. 植物生态学报, 2014, 38(10): 1029–1040. doi: 10.3724/SP.J.1258.2014.00097.
[40] WANG Y J, YANG Y Z, ZHANG J X, et al. Preliminary identification of drought resistance of Chinese wildspecies and its interspecific hybrids [J]. Acta Hort Sin, 2004, 31(6): 711–714. doi: 10.3321/j.issn: 0513-353X.2004.06.001.
王跃进, 杨亚州, 张剑侠, 等.中国葡萄属野生种及其种间F1代抗旱性鉴定初探 [J]. 园艺学报, 2004, 31(6): 711–714. doi: 10.3321/ j.issn:0513-353X.2004.06.001.
[41] CHEN L T. Study on the ecological characteristics of[J]. Sci Technol Inno, 2011, 15(20): 228. doi:10.3969/j.issn.1673-1328. 2011.20.228.
陈丽婷. 葛藤的生态学特性研究 [J]. 科学技术创新, 2011, 15(20): 228. doi:10.3969/j.issn.1673-1328.2011.20.228.
[42] SAURA-MAS S, SHIPLEY B, LLORET F. Relationship between post-fire regeneration and leaf economics spectrum in Mediterranean woody species [J]. Funct Ecol, 2009, 23(1): 103–110. doi: 10.1111/j. 1365-2435.2008.01474.x.
[43] CHAVES M M, MAROCO J P, PEREIRA J S. Understanding plant responses to drought-from genes to the whole plant [J]. Funct Plant Biol, 2003, 30(3): 239–264.
[44] LIU C C, LIU Y G, GUO K. Ecophysiological adaptations to drought stress of seedlings of four plant species with different growth forms in karst habitats [J]. Chin J Plant Ecol, 2011, 35(10): 1070–1082. doi: 10. 3724/SP.J.1258.2011.01070.
刘长成, 刘玉国, 郭柯. 四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性 [J]. 植物生态学报, 2011, 35(10): 1070–1082. doi: 10.3724/SP.J.1258.2011.01070.
[45] YIN X L, WEN J, LIU X, et al. Study on leaf stoma density of 12 representative genus of Rosaceae family [J]. N Fruit, 2008, 31(1): 4–6. doi: 10.3969/j.issn.1001-5698.2008.01.002.
尹秀玲, 温静, 刘欣, 等. 蔷薇科12属代表植物叶片气孔密度研究 [J]. 北方果树, 2008, 31(1): 4–6. doi: 10.3969/j.issn.1001-5698.2008. 01.002.
[46] LIU J Y, FU P L, WANG Y J, et al. Different drought-adaptation strategies as characterized by hydraulic and water-relations traits of evergreen and deciduous figs in a tropical karst forest [J]. Plant Sci J, 2012, 30(5): 484–493. doi: 10.3724/SP.J.1142.2012.50484.
刘金玉, 付培立, 王玉杰, 等. 热带喀斯特森林常绿和落叶榕树的水力特征和水分关系与抗旱策略 [J]. 植物科学学报, 2012, 30(5): 484–493. doi: 10.3724/SP.J.1142.2012.50484.
Studies on Leaf Functional Traits of Nine Woody Lianas in the Karst Area of Central Guizhou Province
WANG Mengjie1,2, RONG Li1,2*, LI Tingting1,2, WANG Qi1,2, YE Tianmu1,2
(1. School of Geography and Environmental Sciences (School of Karst Science), Guizhou Normal University, Guiyang 550025, China; 2. Puding Karst Ecosystem Research Station, Chinese Academy of Sciences, Puding 562100, Guizhou, China)
In order to reveal the ecological strategies of lianas in karst habitats, the leaf function traits and their relationships of nine woody lianas in Chenqi watershed of the Puding Karst Ecosystem Observation and Research Station of Chinese Academy of Sciences were studied. The results showed that six leaf functional traits, such as leaf area, leaf thickness, chlorophyll content, specific leaf area, leaf tissue density and leaf dry matter content, had varied in different degrees. The interspecific variation of traits ranged from 9.24% to 98.18%, while intraspecific variation from 0.64% to 39.71%. The intraspecific coefficient of variation of traits in deciduous plant was lower than that in evergreen plants. Compared with leaves of karst trees and shrubs, lianas had lower thickness, tissue density and higher area and dry matter content. There was close relationship among leaf functional traits in lianas. Specific leaf area had significant negative correlation with leaf tissue density and leaf dry matter content, and leaf tissue density had significant positive correlation with leaf dry matter content. Chlorophyll content had significant negative correlation with leaf dry matter content. Leaf thickness had significant positive correlation with chlorophyll content, and significant negative correlation with leaf tissue density. Except for leaf thickness, leaf area had significant correlation with other traits. Combined the leaf economic spectrum theory analysis,var.,, andbelonged to “quick-return type”, andand,andbelonged to “slow-return type”. The functional differentiation ofandwas obvious, fall in between. Therefore, it was suggested that lianas adopt different adaptation strategies by weighing the relationship among leaf functional traits in order to adapt to the arid and barren karst environment.
Karst; Woody lianas; Leaf; Functional trait; Variation; Leaf economic spectrum
10.11926/jtsb.4328
2020-11-02
2021-03-12
国家十三五重点研发计划课题(2016YFC0502607);贵州省科技计划重大专项([2017]5411);贵州省世界一流学科建设计划项目([2019]125)资助
This work was supported by the Thirteenth Five Program for National Key Research and Development of China (Grant No. 2016YFC0502607), the Key Program for Science and Technology of Guizhou (Grant No. 2017-5411), and the Program for World-class Discipline Construction of Guizhou (Grant No. 2019-125).
王梦洁(1996~ ),女,硕士研究生,主要从事植物地理学研究。E-mail: 1074664997@qq.com
. E-mail:ronglit@126.com
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
阅读(科学探秘)(2020年8期)2020-11-06
新疆农业科学(2020年1期)2020-02-14
乡村地理(2019年2期)2019-11-16
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
中国麻业科学(2015年5期)2015-12-28
文化月刊·下旬刊(2014年6期)2014-08-28
食品工业科技(2014年6期)2014-05-10
