
深紫色大白菜-芥菜附加系的创制及特性分析
2021-10-11 03:57:00张德双辛晓云张凤兰余阳俊赵岫云于拴仓汪维红苏同兵李佩荣卢桂香
中国蔬菜 2021年9期
张德双 辛晓云 赵 鸿 张凤兰 余阳俊 赵岫云 于拴仓 汪维红 苏同兵 李佩荣 卢桂香
(北京市农林科学院蔬菜研究中心,北京 100097)
大白菜起源于我国,种质资源丰富,生态类型繁多。虽然我国拥有紫色的芸薹属蔬菜资源,但是缺乏具有深紫色叶片的大白菜种质,尤其匮乏叶片正面和背面均为深紫色的大白菜材料。紫色大白菜富含花青苷,具有抗氧化衰老、癌症、动脉硬化,减少夜盲、近视,防止神经紊乱、老年痴呆症,增强机体免疫力等保健功能。随着人们对美好、健康、高质量生活方式的不断追求,紫色大白菜因其色泽艳丽,富含花青苷等营养成分,日益受到消费者喜爱。
在高等植物中,以种间杂交和回交方法,将单个外源染色体附加到受体细胞所含的一整套染色体中,即可创制异源单体附加系(monosomic alien addition lines,MAALs)。异源单体附加系是阐明基因组结构和基因转导的有效工具,也是创制种质材料的有效方法。
目前,紫色大白菜紫色性状的来源主要有4种:紫色芥菜(孙日飞 等,2006;张德双 等,2017)、紫色大白菜(张德双 等,2007)、紫菜薹(张明科 等,2008)和紫红色芜菁(Hayashi et al.,2010)。韩国KWONNONG SEED 公司最早育成了紫色大白菜品种紫宝和紫裔,但由于紫宝和紫裔是不育系配制的品种,没有花粉,无法自交,不能被利用。所以,国内急需类似韩国紫色大白菜的资源材料。自2015 年开始,笔者利用大白菜15 绿1-1-26与深紫色芥菜进行种间杂交,创制出深紫色大白菜种质15NG28(张德双 等,2017)。本试验继续利用深紫色大白菜种质15NG28 与普通绿色大白菜进行杂交,选留深紫色植株再自交、回交,经染色体压片观察,筛选获得纯合的深紫色亲本材料,为选育深紫色大白菜新品种奠定了基础。
1 材料与方法
1.1 试验材料
供试材料深紫色大白菜种质15NG28 及其与绿色大白菜杂交F1中深紫色植株自交获得的深紫色自交系17M-236、17M-237、17M-240、17M-241、17M-243、17M-244、17M-245、17M-247,子叶和真叶的正面、背面、叶脉均为深紫色,但叶柄的正面为深紫色,背面为绿色;普通绿色大白菜纯合自交系5 个,分别为春大白菜2 个(15 绿1-1-25 和15 绿2-1-28),秋大白菜3 个(15-1208、16-1081 和16-1096),杂交种1 个(14-CK2-10),叶片均为绿色,带有毛刺。所有材料均来源于北京市农林科学院蔬菜研究中心白菜课题组。
1.2 试验方法
2016 年春季,种植深紫色大白菜15NG28 和普通绿色大白菜14-CK2-10、15 绿1-1-25、15 绿2-1-28,开花后采用人工剥蕾授粉的方法进行杂交,获得F1。2017—2019 年,选留深紫色个体继续进行自交,与绿色大白菜15-1208、16-1081、16-1096 杂交或回交。同时,从苗期到成株期连续调查杂交、自交、回交各世代中深紫色和绿色植株的数量。2020 年春季,利用稳定遗传的深紫色大白菜与绿色大白菜株系进行杂交,观察后代深紫色的分离比例。2021 年春季,利用纯合的深紫色株系相互杂交,配制8 个F1,秋季播种鉴定8 个F1的性状表现。
2018 年,以细胞质雄性不育系CMS96(绿色大白菜)为母本,与深紫色大白菜自交系17M-236、17M-237、17M-240、17M-241、17M-243、17M-244、17M-245、17M-247 进行杂交,选留深紫色植株的不育系,继续与上述纯合的深紫色株系进行回交,获得深紫色大白菜细胞质雄性不育系,并分别调查不育系×深紫色植株的杂交、回交后代中深紫色和绿色植株的数量。
对稳定遗传的深紫色大白菜株系(深紫色性状不再分离)和后代性状分离的深紫色大白菜株系染色体进行压片观察。在盛花期,取长度1~2 mm的花蕾,立刻固定于卡诺溶液(100%乙醇∶冰醋酸为3V∶1V)中,置于4 ℃条件下4 h 以上,将花蕾转移到70%酒精中备用。取出花蕾中的花药,浸润到蒸馏水中,在室温下放置30 min 以上。再将花药放入含有2.5%纤维素和2%果胶酶的溶液中,置于37 ℃条件下2.5 h,再在蒸馏水中浸润20 min。取出花药置于载玻片上,用刀片捣碎,滴1滴DAPI 溶液,在显微镜下计数。
2 结果与分析
2.1 深紫色15NG28 与绿色大白菜杂交F1 的性状分析
由表1 可见,在以深紫色15NG28 为母本的杂交F1中,16M-170 和16M-171 均出现一定比例的深紫色植株,而在以普通绿色大白菜为母本的杂交F1(16M-172)中没有发现深紫色植株。所有深紫色植株的叶片颜色均表现为深紫色,包括子叶和真叶的正面、背面、叶脉以及叶柄的正面全部为深紫色,但是叶柄背面为绿色,不存在中间的过渡紫色。说明以深紫色植株为雌配子体、绿色植株为雄配子体的杂交后代更容易出现深紫色基因型植株,但是深紫色和绿色植株的数量比没有规律性,深紫色和绿色不存在显隐性关系。可见深紫色15NG28 的紫色性状不是简单的质量性状或数量性状。
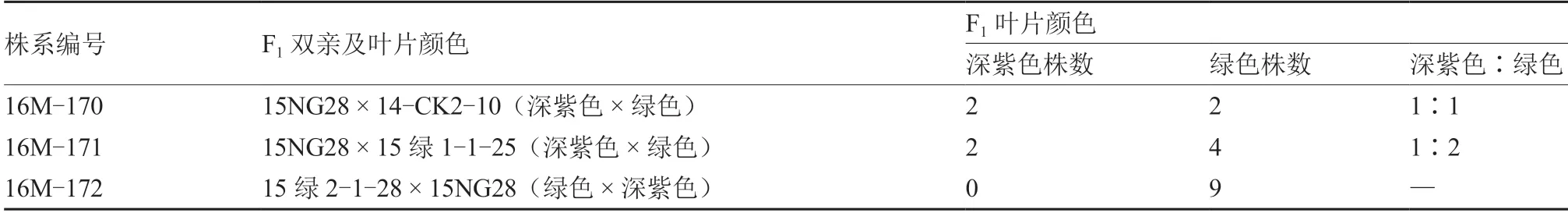
表1 深紫色15NG28 与绿色大白菜杂交F1 的性状表现
2.2 深紫色15NG28 杂交后代中深紫色植株自交、回交以及杂交后代的性状分析
由表2~4 可见,在深紫色性状出现分离的株系中,全部株系的深紫色与绿色的分离比例均不符合孟德尔遗传规律,同时分离比例差异很大,且与自交、回交以及繁殖代数均没有明显的相关性。由表5 可见,深紫色BC1F3中,19M-617 深紫色株出现的比例最高,19M-639 深紫色株出现的比例最低。
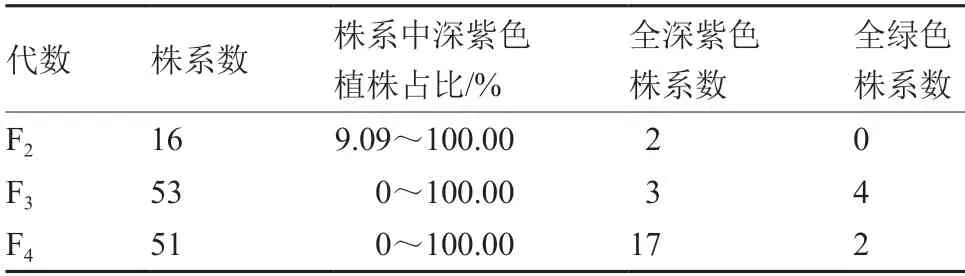
表2 深紫色植株自交后代的性状表现
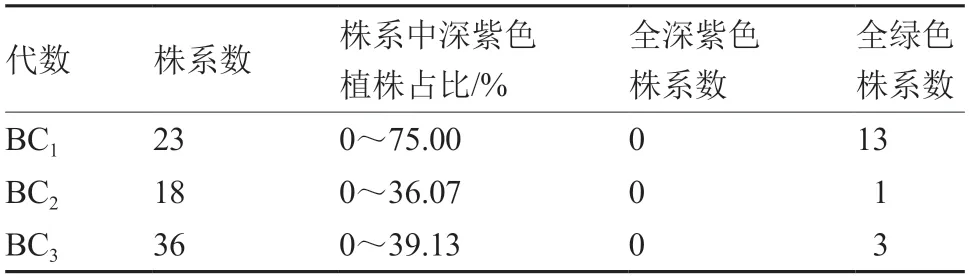
表3 深紫色植株与绿色大白菜回交后代的性状表现
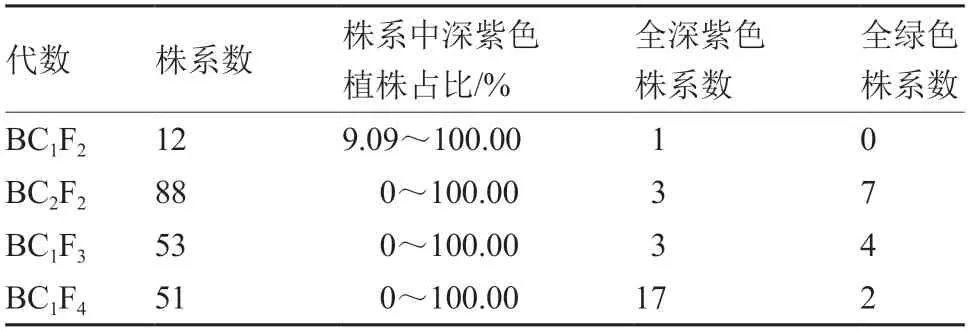
表4 深紫色植株与绿色大白菜回交后再自交后代的性状表现
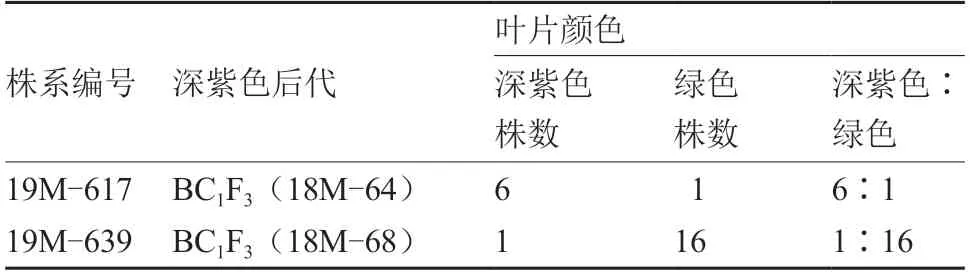
表5 深紫色大白菜自交BC1F3 的性状表现
2.3 纯合深紫色植株的自交后代及与绿色大白菜杂交F1 的性状分析
为了验证大白菜深紫色性状的稳定性,将自交F4的17 个全紫株系继续自交2 代,结果发现所有自交后代全部表现为深紫色,深紫色性状不再分离。利用这17 个纯合的深紫色株系与普通绿色大白菜进行杂交,共获得14 个杂交F1株系。这14个F1株系的深紫色性状依然分离,其中12 个株系出现深紫色和绿色的分离,深紫色植株占比最高的达到31.25%,2 个株系全部为绿色。2021 年8 月,鉴定纯合深紫色亲本间的杂交F1,结果8 个F1没有叶色分离,全部表现为深紫色,这一结果充分说明本试验获得的深紫色大白菜亲本可以应用于新品种选育,配制深紫色的大白菜杂交种。
观察深紫色大白菜苗期至抱球期的性状,发现在深紫色大白菜自交、回交以及杂交后代中,所有的深紫色植株叶片颜色都呈现深紫色,包括子叶和真叶的正面、背面、叶脉以及叶柄的正面全部为深紫色,叶柄背面为绿色,无中间的过渡颜色(图1)。

图1 纯合深紫色大白菜的性状表现
2.4 纯合深紫色大白菜转育细胞质雄性不育系CMS96 后代的性状分析
由表6 可见,17 个纯合深紫色大白菜与细胞质雄性不育系CMS96 回交获得的13 个BC1株系和46 个BC2株系中,均未出现全紫株系,但随着回交代数的增加,深紫色植株所占比例逐步增加,如从BC1的5.26%~20.51%到BC2的3.70%~50.79%,再到BC3的12.12%~64.29%。由此推测,如果以纯合深紫色大白菜株系为父本系,与深紫色CMS96 不育系进行多代回交,随着回交代数的增加,可以获得纯合的深紫色不育系。
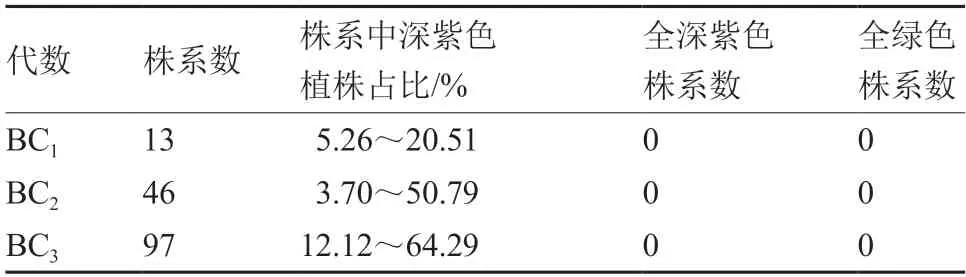
表6 纯合深紫色大白菜转育细胞质雄性不育系CMS96后代的性状表现
2.5 杂合深紫色植株和纯合深紫色植株的染色体数
由上述研究发现,大白菜叶片深紫色性状不符合孟德尔遗传规律,也没有明显的规律性,推测深紫色植株和绿色植株比值的无规律性结果可能是由异源单体附加系导致的,即深紫色的出现可能是由于含有异源染色体附加系。
为了验证该推断,在显微镜下观察稳定遗传和性状分离的深紫色大白菜的染色体数差异。结果表明(图2),性状分离的深紫色大白菜16M-170-21所含的染色体数为21 条,而稳定遗传的深紫色大白菜20M-7 紫鉴所含的染色体数为22 条。

图2 遗传稳定和性状分离的深紫色大白菜染色体数
3 讨论与结论
3.1 深紫色大白菜异源单体附加系的创制
本试验结果表明,在深紫色大白菜细胞质雄性不育系CMS96 杂交后代和纯合深紫色大白菜×绿色大白菜杂交后代中,只发现了深紫色和绿色2 种颜色的基因型,并没有中等紫色或淡紫色叶片的后代。充分说明本试验创制的深紫色性状不符合经典的孟德尔规律,控制叶片深紫色的基因不具有显隐性等特点。这一结论与张淑江(2014)得出的紫红色大白菜自交系B90335-5 的紫红色性状不符合孟德尔遗传规律一致。另外,染色体压片观察结果也表明,稳定遗传的深紫色大白菜的染色体数目增多。深紫色大白菜15NG28 是由绿色大白菜和深紫色芥菜种间杂交获得的种质材料(张德双 等,2017),绿色大白菜的基因组为AA,而深紫色芥菜的基因组为AABB,两者的A 基因组同源性较高,由此推测深紫色大白菜的附加系应来源于B 染色体。为了进一步验证异源单体附加系的试验结论,笔者采用荧光原位杂交技术(fluorescence in situ hybridization,FISH)进行验证,15NG28 及其自交、杂交、回交后代中深紫色植株均为大白菜A 基因组附加黑芥B 基因组的异源单体附加系,纯合基因型为20A+2B,杂合基因型为20A+1B(结果未列出)。
3.2 在深紫色植株的自交4 代中获得了纯合的深紫色大白菜
本试验中,分别在深紫色大白菜F2、F3、F4、BC1F2、BC2F2、BC1F3和BC1F4中发现了2、3、17、1、3、3、17 株深紫色的大白菜材料;在自交4 代中,部分深紫色大白菜株系自交后代全部为深紫色,即全部株系的所有深紫色性状不再分离。这一结果为转育不同类型深紫色大白菜材料,快速获得纯合的深紫色大白菜亲本提供了依据。诚然,考虑到深紫色大白菜为异源单体附加系材料,B 基因组的存在可能增加A 与A 基因组以及A 与B 基因组的交叉和置换,因此获得多个性状都纯合的深紫色大白菜材料可能还需要再自交2 代或更多的代数。截至2020 年底,已获得纯合的深紫色大白菜材料17 份,菜薹(菜心)材料5 份。
3.3 控制深紫色大白菜的深紫色代谢调控途径基因有待进一步研究
本试验还发现,有些深紫色材料的自交后代纯合速度快,由附加1 条B 基因组(染色体数为21条)变为附加2 条B 基因组(染色体数为22 条)只需4 代,虽然很多材料也经过同样的自交4 代,但是染色体数依然还是21 条。同样,虽然同是回交或杂交的后代材料,但是深紫色和绿色植株出现的比例差异很大,如19M-617 深紫色植株所占比率高达85.71%,远高于相同代数的19M-639(深紫色植株所占比率仅为5.88%)。
理论上,假设绿色基因型为AA,产生10 个A 配子;纯合深紫色基因型为AA+2B,产生10个A 和1 个B 配子;二者杂交F1的基因型应该是AA+1B,F1的性状应该表现为深紫色。但是,本试验中,绿色×纯合深紫色或纯合深紫色×绿色的杂交F1性状都出现深紫色和绿色的分离,且深紫色所占的比例很低。同理,如果非纯合深紫色的基因型为AA+1B,配子为10个A或10个A和1个B;绿色基因型为AA,产生配子为10 个A;二者杂交F1表现为绿色AA 和深紫色AA+1B,出现深紫色和绿色植株的随机性很强。考虑到异源附加系1 条B 基因组被A 基因组所排斥或滞后等原因,可能导致B 基因组在后代中更容易被抛弃,深紫色所占的比例应该很少。但是实际上,深紫色所占的比例并没有一定的规律,且随着回交代数的增加,深紫色所占的比例有所提高。相反,纯合的深紫色大白菜无论是自交还是杂交,其后代的紫色性状都不再分离,这是由于B 基因组可以正常联会、配对,均等分配到子代的缘故。由此说明,可能有某种机制控制异源B 基因组在后代中的传递、加倍和深紫色性状遗传等。深紫色大白菜的深紫色性状可能是芸薹种A 与黑芥B 基因组的相互影响,或者是B 基因组对A 基因组影响等所导致的结果。
3.4 异源单体附加系B 基因组对大白菜性状的影响
本试验发现,15NG28 与普通绿色大白菜杂交、回交后代分离出的深紫色植株子叶和真叶的正面、背面、叶脉以及叶柄的正面均表现为深紫色。与绿色大白菜相比,纯合的深紫色大白菜除了深紫色性状外,农艺性状主要表现在叶片数增多,抱球所需的时间稍长,叶球略松,球内叶紫红色,心叶浅黄色,花青苷成分增加。当然,B 基因组对大白菜其他性状还应该有影响。对纯合深紫色大白菜与韩国紫色大白菜品种紫宝和紫裔的叶球剖面进行比较,发现3 份材料的叶球均为深紫色,心叶浅黄色,叶柄剖面为白色。考虑到深紫色大白菜心叶浅黄色的特点,可以借鉴绿色大白菜与紫菜薹杂交、自交后筛选紫心大白菜的方法(张明科 等,2008;He et al.,2020),利用深紫色大白菜与紫心大白菜或者紫菜薹杂交、回交和自交,进一步获得叶球剖面全部为深紫色的深紫色大白菜。
猜你喜欢
中国-东盟博览(政经版)(2023年1期)2023-03-15 01:05:18
海洋通报(2022年4期)2022-10-10 07:40:26
农业工程学报(2022年5期)2022-06-22 12:15:58
亚热带植物科学(2022年1期)2022-05-17 12:39:32
中华戏曲(2020年2期)2020-02-12 05:17:58
四川农业科技(2019年5期)2019-07-01 09:46:46
蔬菜(2016年8期)2016-10-10 06:49:14
浙江柑橘(2016年1期)2016-03-11 20:12:31
阅读与作文(小学高年级版)(2016年3期)2016-03-08 23:38:58
华东理工大学学报(自然科学版)(2015年3期)2015-11-07 09:17:36
