
浓香型白酒新、老窖池分层池底窖泥菌群总氢代谢与乳酸含量之间的关系
2021-09-28 03:27孟雅静王艳丽李安军刘国英何宏魁张会敏周庆伍梁金辉胡心行张光耀
食品科学 2021年18期
孟雅静,王艳丽,丁 峰,李安军,刘国英,何宏魁,张会敏*,王 录,周庆伍,梁金辉,胡心行,张光耀
(安徽古井贡酒股份有限公司,安徽 亳州 236820)
窖泥对浓香型白酒的发酵具有重要作用,一般认为,窖泥越老,酒质越好[1]。新、老窖泥的质量差异,主要表现在菌群组成和理化性质的差异。窖泥菌群组成方面,与新窖泥相比,老窖泥中更丰富的菌属组成形成了完整的窖泥菌群代谢网络。老窖泥中含量更大的瘤胃菌(Ruminococcaceae)[2]和梭菌(Clostridium)[3]可以降解乳酸生成己酸、丁酸、乙酸和氢气等;沉积菌属(Sedimentibacter)[4]和胺杆菌属(Aminobacterium)[5]可以降解氨基酸生成乙酸和丁酸等;互营单胞菌属(Syntrophomonas)[6]可以降解C4~C8的脂肪酸形成乙酸、丙酸和氢气等。以上氢气产生菌可以与氢营养型甲烷菌共生转化氢气生成甲烷[7],形成完整的窖泥菌群代谢链。甲烷菌是窖泥老熟的标志。窖泥菌群代谢链可以将乳酸等有机酸降解生成氢气、甲烷等含氢气体释放。在理化性质方面,与新窖泥相比,老窖泥的乳酸含量远小于新窖泥[8]。窖泥老熟过程中,其菌群组成与理化性质相互适应相互影响,老窖泥中乳酸含量的减少与菌群总氢代谢功能之间的关系是一个值得探讨的问题。
窖泥菌群的功能主要在于产生浓香型白酒的各种风味物质。已知窖泥pH值对窖泥质量影响很大,Tao Yong等[1]研究发现窖泥pH值越接近中性,窖龄越老。优质窖泥pH值偏中性,而新窖泥和退化窖泥pH值偏酸性。窖泥pH值受其中各种有机酸含量的影响,已知乳酸是窖泥中含量最大的有机酸[8],且乳酸的pKa值最小(3.86),对窖泥的pH值影响最大,张会敏等[9]证实窖泥中乳酸与pH值的相关性很强。因此,乳酸对窖泥质量具有重要影响。分析新、老窖泥中总氢相对含量差异,及其与pH值和乳酸之间的关系,有利于分析窖泥菌群代谢与窖泥质量之间的关系。
本研究分别选取安徽省北部某著名浓香型白酒公司的新、老窖池,进行分层分位点池底窖泥取样。分析新、老窖池分层池底窖泥总氢相对含量的变化规律、总氢与pH值及乳酸之间的线性关系,以及乳酸与菌群含量的线性关系,深入讨论新、老窖泥总氢代谢与乳酸含量之间的关系,旨在为窖泥改良提供理论依据。
1 材料与方法
1.1 材料与试剂
窖泥样本取自安徽北部某知名浓香型白酒企业。
乳酸标准品 美国Fluka公司;磷酸二氢钾 上海麦克林生化科技有限公司。
1.2 仪器与设备
FE20 pH计 梅特勒-托利多仪器(上海)有限公司;Acquity超高效液相色谱(配二极管阵列检测器和HSS T3色谱柱(100 mm×2.1 mm,1.8 μm))美国Waters公司;vario EL cube有机元素分析仪 德国Elementar公司。
1.3 方法
1.3.1 分层分位点池底窖泥样本采集
分别选取窖龄不同而发酵工艺相同的新窖池(窖龄6 a)和老窖池(窖龄≥50 a)各6 个,采用三位点四层取样方法对池底窖泥取样[10]。老窖泥样本添加O前缀表示;新窖泥样本添加Y前缀表示。如老窖池中心点1~3 cm的窖泥样本表示为O_Cen_13。每个窖池12 个窖泥样本,共144 个窖泥样本。
1.3.2 pH值和乳酸含量检测
将新鲜窖泥与去离子水按照1∶3(g/mL)混匀静置后用pH计检测窖泥pH值[11]。将新鲜窖泥与去离子水以1∶9(g/mL)混匀,0.22 μm滤膜过滤后,通过液相色谱仪检测乳酸含量。进样量1 µL,流动相为0.02 mol/L KH2PO4溶液,柱流速0.1 mL/min,检测波长208 nm,柱温箱30 ℃。共检测了新、老窖池各6 个,总共12 个窖池的144 个窖泥样本。
1.3.3 窖泥中氢元素的半定量分析
新鲜窖泥经风干粉碎过40 目筛后,用有机元素分析仪在CHNS模式下进行总氢的半定量分析,燃烧管温度1 150 ℃,还原管温度850 ℃。检测新、老窖池各3 个,共6 个窖池的72 个窖泥样本。
1.4 数据处理与统计学分析
Pearson相关性和线性回归分析通过Origin 9实现。差异显著性分析通过方差分析(ANOVA,SPSS 25.0)实现。优势菌属拷贝数是通过每个样本中各优势属序列数占每个样本总序列数的百分比,乘以每个样本的菌群16S rDNA总拷贝数得到。其中伯克氏菌属(Burkholderia)即为Silva数据库注释得到的Burkholderia-Caballeronia-Paraburkholderia菌属。
2 结果与分析
2.1 窖泥中的总氢相对含量及其与pH值之间的关系
新、老窖泥中总氢相对含量均随窖泥深度的增加而降低(图1)。老窖泥中的递降幅度较大,尤其是从0~1 cm到1~3 cm,以及从1~3 cm到3~5 cm显著降低(P<0.05),从3~5 cm到5~7 cm不具有显著差异(P>0.05)。新窖泥的递降幅度较小,除0~1 cm与5~7 cm,以及1~3 cm到3~5 cm有显著差异(P<0.05),其余各层彼此之间无显著差异。新、老窖泥总氢的变化代表其中含氢成分的变化,已知窖泥中有Caproiciproducens、Syntrophomonas和Methanoculleus等产生含氢气体的菌[8],这些菌群在新、老窖泥中的丰度不同[1],因此推测新、老窖泥中总氢相对含量变化的差异可能与新、老窖泥中菌群代谢释放氢气和甲烷等含氢气体的强度不同有关。

图1 新、老窖池分层池底窖泥中总氢相对含量Fig.1 Relative contents of total hydrogen in young and old BPMs
新、老窖泥总氢相对含量与pH值的Pearson相关性分析(图2)显示,两者的Pearson相关系数均呈较强负相关(-0.598 5和-0.865 7),表明新、老窖泥的总氢相对含量随pH值的升高而降低。而两者的回归分析显示:老窖泥总氢相对含量与pH值的线性回归的拟合度更高(R2=0.745 8),指数回归略有提高(R2=0.750 0);新窖泥总氢相对含量与pH值的拟合度较低(R2=0.349 0),三项式回归略有提高(R2=0.365 9)。表明窖泥总氢相对含量的降低(含氢气体的释放)与pH值的升高有很强的联系,且直线回归与非直线回归相比(图2),新、老窖泥的非直线回归相对拟合度更高(R2值略有增大),且老窖泥中的拟合度高于新窖泥。其次,新、老窖泥的直线回归显示其总氢相对含量随pH值增加均匀递降(图2A),而老窖泥的非直线回归显示其总氢相对含量随pH值增加而递降的幅度增加,尤其是在pH 8.0~9.5范围内总氢相对含量下降最快;新窖泥的非直线回归显示其总氢相对含量随pH值增加而递降的幅度递减,其递降幅度主要集中在pH 4.0~7.0之间,pH 7.0之后递降幅度基本不变(图2B)。

图2 新、老窖池分层池底窖泥总氢与pH值之间的直线回归(A)与非直线回归(B)分析(基于72 个新/老窖泥样本)Fig.2 Linear and non-linear regression curves between total hydrogen and pH of young and old BPMs (based on 72 young/old BPM samples)
窖泥pH值与窖泥质量直接相关[1,11-15],窖泥pH值的升高伴随着窖泥总氢的减少(图2),推测窖泥pH值升高所伴随的酸降解与产氢气菌和产甲烷菌的代谢产生含氢气体有关。已知窖泥中乳酸是含量最大的有机酸,窖泥pH值的升高与其中乳酸的降解有关[8]。所以推测窖泥中乳酸降解和氢气、甲烷等含氢气体的释放,使得窖泥中总氢相对含量降低。新、老窖泥样本的pH值(4.33±0.24和5.08±0.32)[8]差异与其中相关菌群降解乳酸并产生含氢气体的强度有直接关系。Van Ginkel等[16]指出活性污泥菌代谢产生氢气的最适宜pH值为5.5,表层老窖泥的pH值(5.87±0.73)[10]正适合氢气产生,0~1 cm到1~3 cm总氢减少幅度最大,表明0~3 cm老窖泥氢气释放量最大。老窖泥丰富的氢气产生菌,如产己酸菌(Caproiciproducens)[17-18]、梭菌(Clostridium)[3]和胺杆菌属(Aminobacterium)[5],产生氢气达到一定浓度时,与之共生的氢营养型甲烷菌的氢转移作用[19]转化氢气形成甲烷[7]。甲烷菌倾向于在偏碱性环境中生存,当环境pH值从中性上升到9.0的弱碱性环境时,氢气产生速率逐渐增大[20],甲烷菌的氢转移作用解除氢抑制,进一步促进氢气产生,因此老窖泥在pH值8.5~9.5范围内更适宜氢气产生(图2B)。不同于老窖泥,新窖泥pH值集中在4.0~7.0,在7.0~9.0内样本的总氢随pH值增加而降低的程度(斜率)反而减小,接近于0。分析其原因,可能与新窖泥菌群丰度比老窖泥低几个数量级[21],以及没有形成完整的菌群代谢链[1]有关。一方面,可能丰度较低的新窖泥菌群代谢生成的氢气浓度较低,无法提供甲烷菌生存的条件;另一方面,甲烷菌的繁殖速度很慢[22],需要较长的时间才可以达到解除氢抑制所需要的代谢强度。因此新窖泥可能在后期pH 7.0~9.0范围内处于氢抑制作用下无法继续产氢。
2.2 窖泥中的总氢相对含量与乳酸之间的关系
如图3所示,新、老窖泥总氢与乳酸的Pearson相关系数(0.600 7和0.719 2)表明,新、老窖泥的总氢相对含量随乳酸含量的增加而增加,换言之新、老窖泥的总氢相对含量随乳酸含量的减少而减少。为了验证窖泥中总氢相对含量与乳酸含量之间的统计学关系,进一步建立两者之间的线性回归分析,图3显示:窖泥总氢相对含量与乳酸含量呈较强的线性关系。新、老窖泥相比,老窖泥中两者直线回归的拟合度较强(R2=0.510 3),新窖泥中的拟合度较弱(R2=0.351 7)。新、老窖泥中的总氢相对含量随乳酸含量的减少而减少,推测新、老窖泥中总氢相对含量的减少与乳酸的降解有关,与已有结论一致[8]。与新窖泥斜率(5.101×10-5)相比,老窖泥中总氢相对含量与乳酸含量的直线回归斜率更大(0.001),且新窖泥中乳酸含量远高于老窖泥,表明老窖泥总氢相对含量随乳酸含量减少的幅度远大于新窖泥中的幅度。窖泥中乳酸绝对量受3 方面影响:黄水中乳酸的浸入量、窖泥菌群产生的乳酸量、窖泥菌群降解的乳酸量,其中前两者为增量,后者为减量。与新、老窖泥中的乳酸含量(28.8 g/kg和10.72 g/kg)[9]相比,新、老窖池黄水中乳酸质量浓度高达80 g/L,且新、老窖池的黄水乳酸含量之间没有显著差异[23],推测黄水浸入的乳酸含量对窖泥中乳酸增量起决定作用。新、老窖泥中乳酸的减少主要都是乳酸被降解的原因所致。新、老窖泥相比,老窖泥样本较集中,新窖泥样本较分散,推测老窖泥中菌群组成与理化性质比较均匀,而新窖泥的理化性质与菌群组成的均一度较差。已知窖泥中含有降解乳酸生成氢气的菌[24]和产生甲烷的菌,能够协同互助利用氢气产生甲烷而解除氢抑制,如Ruminococcaceae[2]、Clostridium[3]、Methanoculleus[11]这些菌在窖泥中均为优势菌属,且在老窖泥中的含量高于新窖泥[8]。因此推测老窖泥中降解乳酸并产生含氢气体的菌的丰度比新窖泥大。
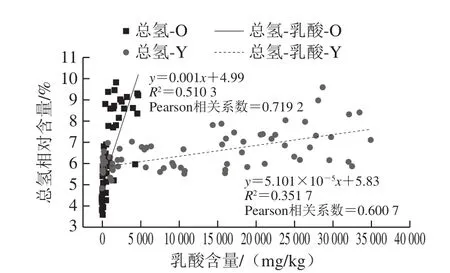
图3 新、老窖池分层池底窖泥乳酸含量与总氢之间的线性回归分析Fig.3 Linear regression curve between lactic acid and total hydrogen of young and old BPMs
2.3 新、老窖池分层池底窖泥中优势菌属的拷贝数
为进一步验证新、老窖泥中乳酸降解菌的丰度与乳酸之间的相关性,有必要首先分析其中所有优势菌属的丰度。本研究借鉴新、老窖池分层池底窖泥中16S rDNA V4区高通量测序数据(SRR8985614~SRR8985757),将可操作分类单元列表和其实时聚合酶链式反应绝对定量的16S rDNA V4区总拷贝数相结合,计算出各组窖泥样本中优势菌属(相对丰度>2%)[10]的拷贝数如图4所示。Aminobacterium(3.56×103copies/g和3.67×107copies/g)和Fastidiosipila(3.94×103copies/g和2.75×107copies/g)是新、老窖泥中含量差异最显著的菌属,且主要存在于老窖池的1~7 cm中。本研究结果显示老窖池分层池底窖泥中各优势菌属拷贝数普遍高于新窖池分层池底窖泥。老窖池各分层池底窖泥中的优势菌属含量比较接近,推测与老窖泥的均一性较好有关,可能老窖泥经过多年发酵驯化已达到老熟状态,其菌群变化已趋于稳定,其理化环境更适合这些优势菌属的生长。新窖池分层池底窖泥中的优势菌属含量略有差异,推测新窖泥的均一性较差,新窖池窖泥的理化环境相对较差,如pH值偏低、乳酸含量偏高[1]等,不利于各优势菌属的生长,因此新窖池各分层池底窖泥中的优势菌属的含量不均衡。

图4 新、老窖池分层池底窖泥中优势菌属拷贝数Fig.4 Absolute copy numbers of dominant genera in different layers of young and old BPMs
2.4 窖泥中的乳酸含量与窖泥优势菌属之间的关系
鉴于新、老窖泥的乳酸含量差异与菌群丰度差异有关,因此有必要建立两者之间的关系,确认两者之间的相关性。老窖池分层池底窖泥的乳酸含量与优势菌属总拷贝数的Pearson相关系数为0.025 6,表明在老窖泥中两者基本没有相关性,可能是老窖泥的菌群已经成熟,代谢功能比较完善,且各分层池底窖泥的菌属含量变化范围较小,乳酸含量已经不会对它造成威胁。而在新窖泥中两者呈很强的负相关(-0.665 2),表明新窖泥中乳酸含量随窖泥优势菌属总拷贝数的增加而减少。为了验证这个关系,建立新窖池分层池底窖泥乳酸含量与优势菌属总拷贝数之间的回归分析(图5)。发现新窖泥的乳酸含量与优势菌属总拷贝数呈线性关系,拟合度适中(R2=0.430 4),乳酸含量随优势菌属总拷贝数的增加而减少,表明降解乳酸是新窖泥菌群很重要的一项代谢机能,推测新窖泥中大量乳酸的存在影响其菌群生存,因此新窖泥菌群集中于降解乳酸的代谢机能。

图5 新、老窖池分层池底窖泥乳酸含量与优势菌属总拷贝数之间的关系Fig.5 Relationship between lactic acid content and total copy number of dominant genera in young and old BPMs
鉴于新、老窖泥中乳酸含量与优势菌属丰度差异有关,以及两者在新、老窖泥中的相关性不同,分别给出新、老窖池分层池底窖泥中21 个优势属的拷贝数与乳酸含量之间的Pearson相关系数(图6),发现新窖泥中与乳酸含量相关性较强(>0.5)的优势菌属有15 个,均与乳酸含量呈负相关;而老窖泥优势菌属拷贝数与乳酸的相关系数值正负参半,且相关性较强(>0.5)的菌属较少,只有Caproiciproducens和Hydrogenispora。新、老窖泥菌群优势菌属拷贝数与乳酸含量的相关系数进一步表明新窖泥中的大部分菌属都处在降解乳酸的状态,而老窖泥中已经没有降解乳酸的压力。

图6 新(A)、老(B)窖池分层池底窖泥中21 个优势属拷贝数与乳酸含量的Pearson相关系数Fig.6 Pearson correlation coefficients between total copy numbers of 21 dominant genera and lactic acid of young and old BPMs
将新窖泥(图6A)中与乳酸含量相关性强度大于0.5的15 个优势菌属总拷贝数和乳酸含量计算Pearson相关系数(-0.743 6),发现新窖泥中这15 个优势属总拷贝数与乳酸的相关性高于新窖泥菌群(-0.665 2)(图5)。表明这15 个菌属在参与降解新窖泥的乳酸过程中发挥重要作用。为验证此结论建立线性回归分析(图7),发现新窖泥中这15 个优势属总拷贝数与乳酸含量线性回归的拟合度(R2=0.546)高于新窖泥菌群(R2=0.430 4),证实是这15 个菌属共同参与新窖泥中乳酸的降解。已知,Caldicoprobacter和Sedimentibacter具有降解乳酸的功能[24],尽管其余菌没有直接参与降解乳酸,但它们彼此之间的协同作用[25]形成代谢网络,为降解乳酸服务,而非每种菌属均有降解乳酸的功能。比如Caproiciproducens可以代谢产生H2、乙醇、乙酸、丁酸和己酸[26-27],Fermentimonas和Proteiniphilum能够产生乙酸、丙酸和H2[28],Hydrogenispora[29]可产生醋酸盐、乙醇和H2,Garciella[30]和Caldicoprobacter[31]发酵产H2、CO2、乳酸盐、乙酸盐,Caloribacterium发酵产生醋酸盐、H2、CO2[32],Syntrophomona与甲烷菌共培养,可以降解C4~C8的中长链脂肪酸(如己酸和丁酸等)产生乙酸、丙酸和H2[33-34]。上述产气菌产生的H2、CO2和乙酸可以作为Methanoculleus[22,35](氢营养型甲烷菌)的底物产生甲烷。含氢气体(H2、CH4)的释放,不仅降低窖泥总氢相对含量,使窖泥pH值升高,又优化窖泥的厌氧环境,有利于乳酸降解菌的生长代谢。

图7 新窖池分层池底窖泥中15 个优势属(Pearson相关系数绝对值>0.5)总拷贝数与乳酸之间的线性回归分析Fig.7 Linear regression analysis of total copy number of 15 dominant genera (absolute value of Pearson coefficient > 0.5) and lactic acid in different layers of young BPMs
以上菌属所形成的代谢网络,其降解乳酸的功能与产氢气、甲烷等直接相关,而窖泥中的乳酸降解产氢菌[2-3]甲烷菌协作降解乳酸,产生弱酸,并释放氢气与甲烷等含氢气体,减少窖泥总氢的同时提高窖泥pH值和解除产氢菌受氢抑制作用。新、老窖泥相比,老窖泥菌群的代谢网络相对更完整,新窖泥中甲烷菌(如Methanoculleus)的丰度更少,而甲烷菌是解除“氢抑制”的终端,但是甲烷菌的繁殖速度很慢,如Methanoculleus的生长速率为2.5~35 h/代[22]。因此新窖泥需要很多年才能老熟,使其菌属代谢网络完整。
综上,老窖池由于已经达到老熟,各分层池底窖泥中菌群代谢功能的重点已经不在降解乳酸上;而新窖泥还在不断进化中,各分层池底窖泥的菌群功能不够成熟,面对的生存压力主要是乳酸对其影响。乳酸不仅对窖泥pH值影响很大,还具有一定的抑菌功能[36-37],因此新窖泥中菌群为了生长繁殖,整体功能都在于降解乳酸,改善窖泥的理化环境。
3 结 论
本研究通过分析新、老窖池分层池底窖泥总氢代谢与乳酸含量之间的关系,得出结论:1)新、老窖池分层池底窖泥中总氢相对含量均随窖泥深度的增加而降低,且老窖泥的递降幅度较大,呈显著降低趋势,新窖泥之间总氢变化不显著;新、老窖泥总氢相对含量随窖泥的pH值升高而降低,且老窖泥的递降幅度大于新窖泥;新窖泥总氢相对含量在pH 7.0之后递降幅度趋于平缓,而老窖泥的递降幅度增大。2)新窖泥中乳酸含量远大于老窖泥,新、老窖泥总氢相对含量随乳酸含量的减少而减少,与新窖泥(5.101×10-5)相比老窖泥的直线回归斜率更大(0.001),表明老窖泥总氢相对含量随乳酸含量减少的幅度远大于新窖泥。3)老窖池分层池底窖泥中的优势菌属总拷贝数普遍高于新窖池;老窖池各分层池底窖泥的优势菌属含量分布更均匀,新窖池各分层池底窖泥的优势菌属含量略有差异。4)老窖泥的乳酸含量与优势菌属总拷贝数之间基本没有相关性(0.025 6),新窖泥乳酸含量与优势菌属总拷贝数有较强的负相关(-0.665 2)。新窖泥中与乳酸含量相关性较强(>0.5)的优势菌属有15 个,均与乳酸含量呈负相关;而老窖泥优势菌属与乳酸的相关系数值正负参半,与乳酸含量相关性较强(>0.5)的优势菌属只有Caproiciproducens和Hydrogenispora。可能新窖泥中大部分菌属都处在降解乳酸的状态,而老窖泥中已经没有降解乳酸的压力。5)新窖泥中相关性较强的15 个优势属总拷贝数与乳酸含量的回归拟合度(R2=0.546 0)和相关性(-0.743 6)均高于新窖泥菌群的回归拟合度(R2=0.430 4)和相关性(-0.665 2),可能是这15 个菌属在共同参与新窖泥的乳酸降解过程中发挥重要作用。
综上,推测老窖泥的菌群代谢链比较完整,各分层池底窖泥的菌属含量相对稳定,而新窖泥的菌群还在进化中,代谢链不够完善。因此老窖泥中氢代谢及酸降解速度均比新窖泥快,推测降解乳酸已经不是老窖泥菌群代谢的主要压力。新窖泥还没达到老熟状态,优势菌属相对较少,所以乳酸大量存在可能对新窖泥菌群的生长影响很大,推测新窖泥的大部分菌属都参与降解乳酸及氢代谢,才能缓解和改善窖泥的理化环境。
猜你喜欢
廉政瞭望(2022年19期)2022-11-16
右江民族医学院学报(2022年2期)2022-05-19
河北医学(2021年10期)2021-10-27
装备维修技术(2021年45期)2021-03-03
渔业致富指南(2020年6期)2020-12-19
岷峨诗稿(2020年4期)2020-11-18
中国临床医学影像杂志(2019年6期)2019-08-27
酿酒科技(2019年10期)2019-01-05
中国化妆品(2018年6期)2018-07-09
含笑花(2018年1期)2018-01-24
