
迭山北坡云冷杉林火烧迹地灌木群落特征和物种多样性研究*
2021-09-26 12:27:48赵安周晓雷田青史瑞锦黄海霞曹雪萍陆刚周旭姣
西部林业科学 2021年5期
赵安,周晓雷,田青,史瑞锦,黄海霞,曹雪萍,陆刚,周旭姣
(1.甘肃农业大学 林学院,甘肃 兰州 730070;2.广西大学 林学院,广西 南宁 530000;3.甘肃农业大学 草业学院,甘肃 兰州 730070)
林火是陆地生态系统的主要干扰类型之一,它不仅改变局部生境的植被结构与动态,而且能改变景观格局与生态过程[1-2]。林火干扰在20世纪初受到林学家和生态学家的关注,但一直被认为是破坏生态系统和导致群落逆行演替的干扰因子之一,直到20世纪60年代,人们才逐渐意识到林火干扰对生态系统具有两面性和普遍性[3-4],并对森林植被林火干扰开展了广泛的研究。研究表明,林火干扰后植被群落变化主要取决于火烧强度,轻度林火干扰能够加速植物群落的演替,有利于改善林地环境,促进物质循环与林木的更新、生长和发育,对于生态平衡与稳定起着重要作用;重度林火干扰能对森林造成毁灭性的破坏,使森林植被化为灰烬,森林结构和功能丧失,种子库遭到破坏,在灾后甚至能引发大面积的病虫害,造成更加严重的损失[5-8]。中度林火干扰对森林植被的影响尚未有一致定论,Swan等[9]和Roberts[10]认为与重度火烧相比,中、轻度能对植物物种多样性产生更积极的影响。杨一等[11]认为轻度火干扰可以增加次生灌丛生物多样性、促进群落更新,中度火干扰下木本植物物种多样性丧失较大,而草本植物多样性显著增加,不利于群落的正向演替。
林火干扰对森林群落的直接作用使短期内植物种类减少和物种多样性降低,但随着森林群落的演替与发展,物种多样性回归正常水平[5,8,12]。本研究以青藏高原东北边缘迭山北坡云冷杉林重度火后恢复15 a的灌木群落为研究对象,通过样方调查和数据分析,探究火烧迹地森林演替过程中不同坡位森林群落的恢复特征,阐明迭山北坡火烧迹地不同坡位处森林植物群落恢复与重建和群落演替机制,为当地林业部门对火烧迹地森林的恢复和演替进程中进行合理的干预,以为尽快恢复森林重要的生态效益提供合理的依据。
1 材料与方法
1.1 研究区概况
研究区域位于青藏高原东北边缘迭山北坡(34°10′40.66″~34°10′47.30″N,103°12′51.35″~103°12′57.43″E),地处秦岭西,白龙江中游高山峡谷之中,属于甘南高原山地植被区,是长江流域和黄河流域的重要分水岭,对长江流域和黄河流域的水土保持、水源涵养和生物多样性保护具有重要意义[13-14]。在气候分区上属于甘南高寒湿润区,处于大陆性气候与季风气候的过渡带上,年平均气温在8~11 ℃之间,无霜期147 d,降水量635 mm,平均海拔3 200 m,地形相对高差约1 200 m,平均坡度30°~50°。森林覆盖率为75%,乔木树种以针叶树为主,主要以云杉(PiceaasperataMast.)、冷杉[Abiesfabri(Mast.)Craib]、红桦(BetulaalbosinensisBurkill)等为优势树种;灌木层主要以唐古特忍冬(LoniceratanguticaMaxim.)、小叶蔷薇(RosawillmottiaeHemsl.)、大刺茶藨子(RibesalpestreWall.ex Decne.)、扁刺蔷薇(RosasweginzowiiKoehne)等为优势树种;草本层主要以密生薹草(CarexcrebraV.I.Krecz.)、总状橐吾[Ligulariabotryodes(C.Winkl.)Hand.-Mazz.]、酢浆草(OxaliscorniculataL.)、峨参[Anthriscussylvestris(L.)Hoffm.]、三角鳞毛蕨[Dryopterissubtriangularis(C.Hope)C.Chr.]等为优势种。
由于该区域是藏民族聚居地,其生活和取暖主要以放牧和薪柴为主,森林生态系统受到干扰较为严重。历年来,青藏高原东北边缘曾连续多年发生森林火灾,次数频繁,森林生态系统、群落结构和景观格局遭到破坏,生物多样性减少。本研究区森林火灾发生于2005年4月19日,位于迭部县益哇镇纳家村47林班,过火面积约为5.72 hm2,火势主要由下坡位向上坡位蔓延,森林火灾级别为重度,过火后其森林生态系统遭到毁灭性破坏,火烧迹地上生物多样性完全丧失,现以灌木种类为建群种。目前,对于该研究区域火烧迹地森林植物群落恢复与重建、群落演替机制鲜有相关报道。
1.2 研究方法
根据研究区域的实际地形地貌,可将研究区坡位分为上坡位、中坡位和下坡位,中坡位正中地段属于险坡(≥45°),故在设置样地时,下坡位至中坡位约50 m,中坡位至上坡位约80~100 m。采用典型抽样法在不同坡位处设置2个面积大小为20 m×20 m(闭合差<1/200)的样地,记录坡度、坡向、海拔、坡位、枯落物厚度、土层深度等(表1);在样地中设置5个5 m×5 m的灌木样方,调查灌木种类、株(丛)数、平均树高、地径、冠幅、盖度等;在每个灌木样方中设置1个1 m×1 m草本样方,调查草本种类、株(丛)数、平均高度、盖度等。此次共设置灌木样方和草本样方分别都为30个,共计60个样方。
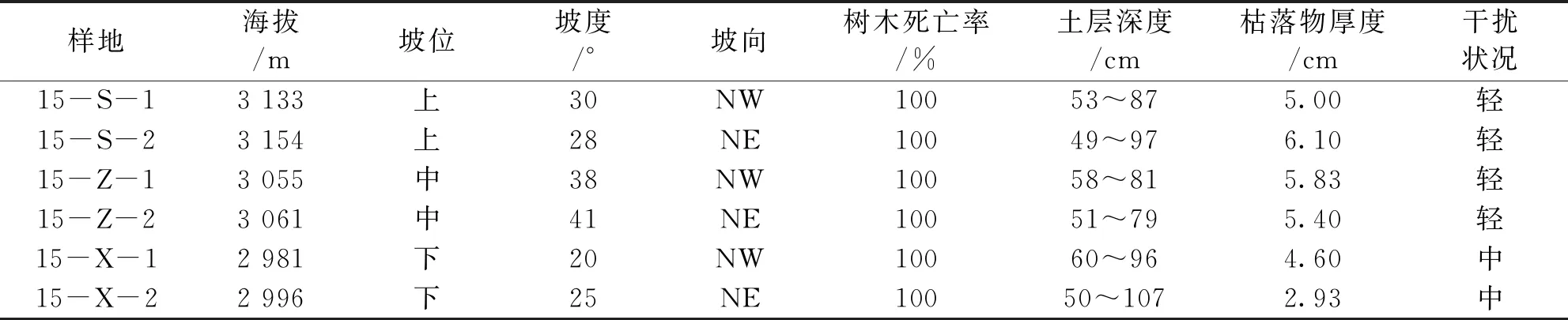
表1 火烧迹地样地的基本信息
1.3 数据分析
采用R语言[15]、SPSS和Excel 2010进行群落的重要值计算、方差分析、群落组成与结构和物种多样性分析。
1.3.1 重要值计算
重要值(IV)=相对密度+相对频度+相对盖度[16-17]。
1.3.2 群落结构特征
(1)垂直结构 以0.2 m为高阶来研究火烧迹地不同坡位群落垂直结构[18-19]。
(2)水平结构 以4 mm为径阶,通过分析灌木平均地径的方法来研究火烧迹地的水平结构[18-19]。
1.3.3 α多样性指数计算
选定α多样性中的物种丰富度R、Shannon-Wiener指数、Simpson指数、Pielou均匀度指数来分别衡量火烧迹地灌木层和草本层的植物丰富性、多样性、优势度和分布状况[19-20]。
(1)物种丰富度R
R=S
(2)Shannon-Wiener指数
(3)Simpson指数
(4)Pielou均匀度指数
J=H/Hmax,Hmax=lnS
式中:Pi为每个物种的相对频度;Hmax为最大物种多样性指数;i=1、2、3...;S为物种数。
2 结果分析
2.1 火烧迹地不同坡位典型群落与优势种划分
本次研究的火烧迹地是以灌木为建群种,采用灌木层+草本层优势种命名原则,根据重要值对火烧迹地不同坡位处恢复15 a的植被进行典型群落的划分,并以重要值大于或等于0.2来划分优势种,可将植被划分为以下3种典型群落:上坡位为唐古特忍冬+扁刺蔷薇-密生薹草群落,中坡位为箭竹(FargesiaspathaceaFranch.)-密生薹草群落,下坡位为黑水柳(SalixheishuiensisN.Chao)-野草莓(FragariavescaL.)群落(表2)。

表2 火烧迹地不同坡位处典型群落优势种重要值
2.1.1 唐古特忍冬+扁刺蔷薇-密生薹草群落
灌木层主要优势种有唐古特忍冬、扁刺蔷薇、大刺茶藨子、峨眉蔷薇(RosaomeiensisRolfe)、毛花忍冬(LoniceratrichosanthaBureau & Franch.)、箭竹、山梅花(PhiladelphusincanusKoehne)等。其中,唐古特忍冬重要值最大,为0.481;其次是扁刺蔷薇,重要值为0.431;接着重要值依次排序为大刺茶藨子0.394、峨眉蔷薇0.388、毛花忍冬0.339、箭竹0.327、山梅花0.325。草本层主要优势种有密生薹草、总状橐吾、婆婆纳(VeronicapolitaFries)、野草莓等。密生薹草重要值最大,为0.650;总状橐吾次之,重要值为0.375;其后按重要值排序依次为婆婆纳0.341、野草莓0.338。
2.1.2 箭竹-密生薹草群落
灌木层主要优势种有箭竹[21-22]、山梅花、黑水柳、扁刺蔷薇、大刺茶藨子、唐古特忍冬、峨眉蔷薇等。箭竹为主要建群种,重要值最大,为0.738;其后按重要值排序依次为山梅花0.392、黑水柳0.338、扁刺蔷薇0.336、大刺茶藨子0.333、唐古特忍冬0.295、峨眉蔷薇0.261。草本层主要优势种有密生薹草、总状橐吾、三角鳞毛蕨、野草莓等。密生薹草重要值最大,为0.733;其次是总状橐吾,重要值为0.375;接着三角鳞毛蕨与野草莓重要值都是0.372。
2.1.3 黑水柳-野草莓群落
灌木层主要优势种有黑水柳、峨眉蔷薇、小叶蔷薇、唐古特忍冬、大刺茶藨子、箭竹等。黑水柳为主要建群种,重要值为1.050,要远大于其他优势种;其余优势种重要值依次为峨眉蔷薇0.401、小叶蔷薇0.292、唐古特忍冬0.245、大刺茶藨子0.228、箭竹0.217。草本层主要优势种有野草莓、密生薹草、三角鳞毛蕨等植物。其中,野草莓为主要优势种,重要值最大,为0.568;接着优势种按重要值依次排序分别是密生薹草0.475、三角鳞毛蕨0.418。
2.2 火烧迹地不同坡位植物组成特征
从表2、表3可看出,火烧迹地植被恢复15 a后,在样方中共调查到27科49属60种植物。上坡位调查到植物共23科28属32种,科、属、种分别占火烧迹地总科数、总属数、总种数的77.78%、57.14%和53.33%。其中,灌木层有7科7属11种,以唐古特忍冬、扁刺蔷薇、大刺茶藨子、峨眉蔷薇、毛花忍冬、箭竹、山梅花等植物为主;草本层有16科21属21种植物,以密生薹草、总状橐吾、婆婆纳、野草莓等植物为主。

表3 火烧迹地不同坡位处群落组成特征
中坡位调查到植物共21科27属33种,科、属、种分别占火烧迹地总科数、总属数、总种数的66.67%、55.10%和55.00%。其中,灌木层有6科6属10种,以箭竹、山梅花、黑水柳、扁刺蔷薇、大刺茶藨子、唐古特忍冬、峨眉蔷薇等植物为主;草本层有15科21属23种植物,以密生薹草、总状橐吾、三角鳞毛蕨、野草莓等植物为主。
下坡位调查到植物共25科40属49种,科、属、种分别占火烧迹地总科数、总属数、总种数的88.89%、81.63%和81.67%。其中,灌木层有7科9属14种,以黑水柳、峨眉蔷薇、小叶蔷薇、唐古特忍冬、大刺茶藨子、箭竹等植物为主;草本层有18科31属35种植物,以野草莓、密生薹草、三角鳞毛蕨、车前等植物为主。
研究还发现,在同一坡位中,不同树种的个体数量差异较大(图1),如上坡位中,扁刺蔷薇个体数为31株(丛),大刺茶藨子个体数为104株(丛),箭竹个体数为52株(丛)等。在不同坡位中,同一树种的个体数量差异较大,如大刺茶藨子在上坡位中个体数104株(丛),在中坡位中为59株(丛),在下坡位中为11株(丛)等。下坡位各物种的数量相较于上坡位和下坡位都偏少。

图1 火烧迹地上坡位处各主要树种高阶个体数分布情况
2.3 火烧迹地不同坡位群落结构特征
2.3.1 火烧迹地不同坡位群落垂直结构
不同坡位处群落高度结构存在差异,上坡位和中坡位处群落高阶分布总体上都呈倒“J”型分布,但下坡位群落的高阶分布呈正态和双峰连续分散分布(图2)。其次群落中主要植物种的高度结构也存在着一定的差异,中坡位各主要灌木种的高阶分布类似于倒“J”型,个别树种呈双峰连续分散分布(图3)。上坡位与下坡位灌木层高阶分布基本类似于正态和多峰连续分散分布(图1、图4)。
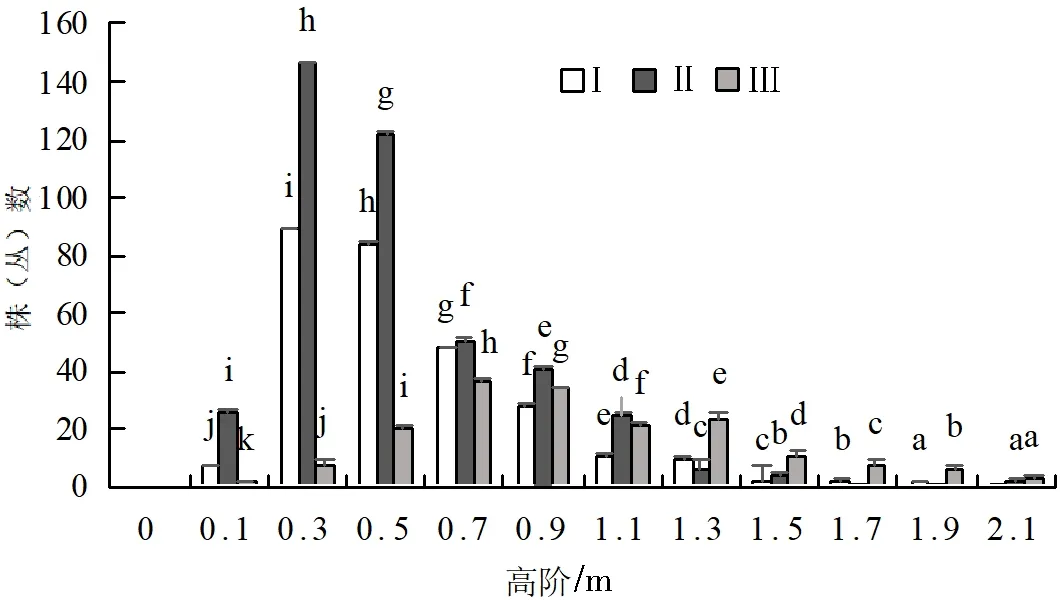
图2 火烧迹地群落高阶个体数分布情况

图3 火烧迹地中坡位处各主要树种高阶个体数分布情况
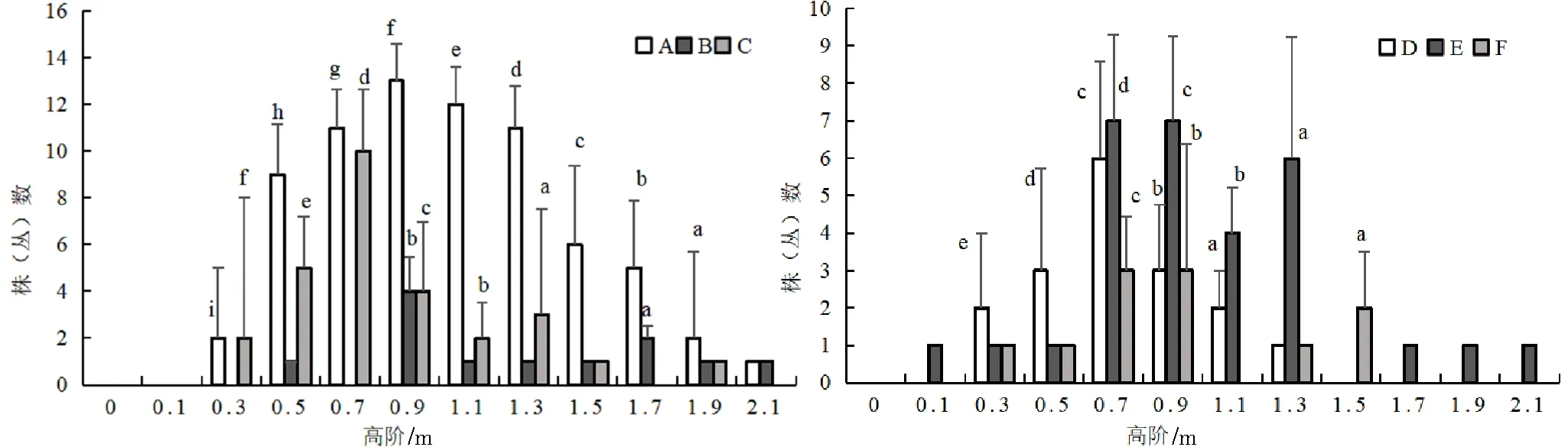
图4 火烧迹地下坡位处各主要树种高阶个体数分布情况
在同一坡位中,各主要树种在高阶范围出现最多个体数量差异较大,且出现最多个体数量时高阶中值也不同,如:中坡位中,高阶中值为0.3 m时,山梅花的最多个体数为41株(丛),扁刺蔷薇的最多个体数为6株(丛),唐古特忍冬的最多个体数为13株(丛)等;上坡位中大刺茶藨子个体数最大值为52株(丛),所对应的高阶中值是0.3 m,而唐古特忍冬个体数最大值为31株(丛),所对应的高阶中值是0.7 m。在不同坡位中,同一树种在高阶范围出现最多个体数量差异较大,如高阶中值为0.3 m时,上坡位中箭竹的最多个体数为15株(丛),中坡位中为35株(丛),下坡位中为10株(丛)。上坡位和中坡位中、小高度级树木数量占比较大,这说明群落具有良好的自我更新能力,下坡位树种高阶分布图呈正态和多峰连续分散分布,具有良好的稳定性[23-25]。
2.3.2 火烧迹地不同坡位群落水平结构
不同坡位处群落的径阶分布都呈现出倒“J”型(图5)。在不同坡位中各主要树种的径阶分布存在差异,在上坡位和中坡位中,各主要树种的径阶分布也基本上呈倒“J”型(图6、图7),下坡位中各主要树种的径阶主要呈正态和多峰连续分散分布(图8)。在同一坡位中,各树种在径级范围内出现最多个体数量差异较大,如在上坡位中,径阶中值为6 mm时,唐古特忍冬的最多个体数为51株(丛),大刺茶藨子最多个体数为75株(丛),毛花忍冬最多个体数为23株(丛)等。同一树种在不同坡位处径级范围内出现最多个体数量差异较大,如径阶中值为6 mm时,唐古特忍冬在上坡位的最多个体数为51株(丛),中坡位为21株(丛),下坡位为11株(丛)。在整个火烧迹地中,与上坡位和中坡位树高阶分布一样,中、小径级树木数量占比较大,这与高度结构相互印证,说明了群落的更新能力较强、稳定性较高,往正向演替[23-25]。

图5 火烧迹地群落径阶个体数分布情况
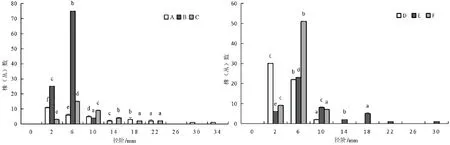
图6 火烧迹地上坡位处各主要树种径级阶个体数分布情况
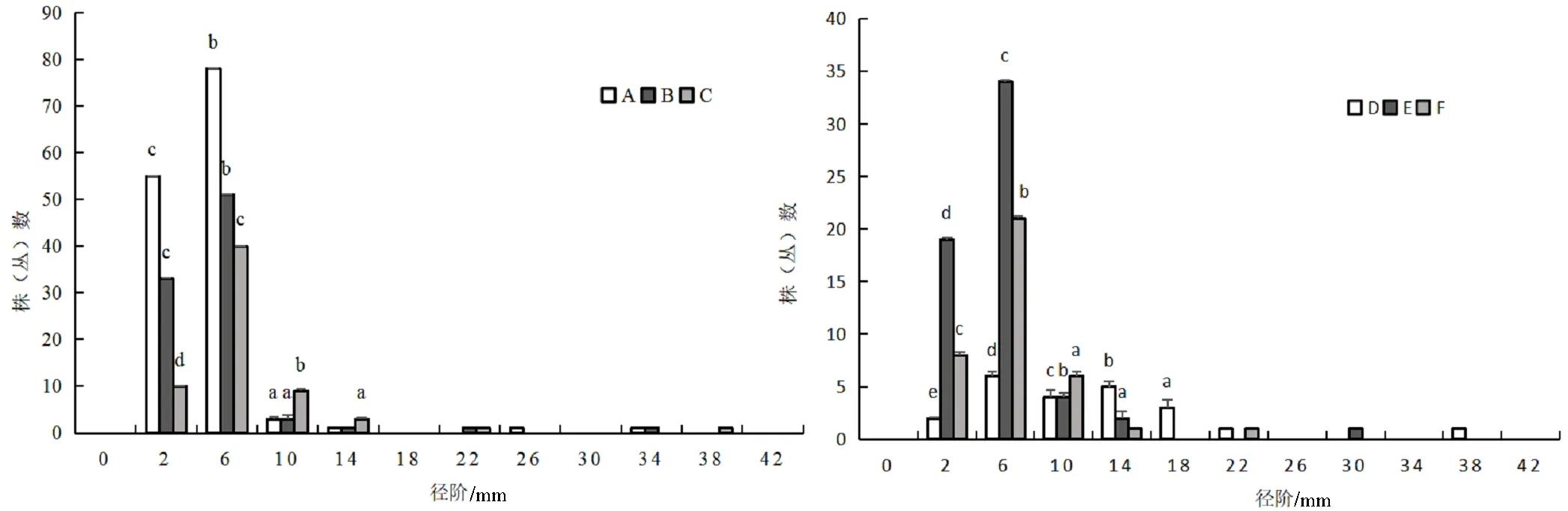
图7 火烧迹地中坡位处各主要树种径级阶个体数分布情况
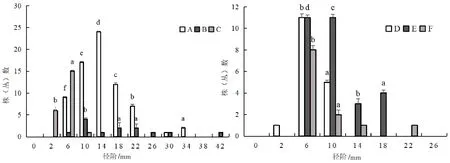
图8 火烧迹地下坡位处各主要树种径级阶个体数分布情况
2.4 火烧迹地不同坡位群落物种多样性特征
由表4可以看出,不同坡位的灌木层和草本层物种多样性指数值存在差异。在灌木层群落中,下坡位的物种丰富度R最大,上坡位次之,中坡位最小;Shannon-Wiener指数上坡位最大,下坡位次之,中坡位最小;Simpson指数与Pielou均匀度指数上坡位最大,中坡位次之,下坡位最小。在草木层群落中,物种丰富度R和Shannon-Wiener指数随着坡位的上升逐渐减少;Simpson指数中坡位最大,上坡位最小;Pielou均匀度指数中坡位最大,上坡位次之,下坡位最小。

表4 不同坡位处灌木层草本层群落物种多样性变化
对于整个火烧迹地植被群落,除上坡位Simpson指数值草本层比灌木层小外,草本层的物种丰富度R、Shannon-Wiener指数和Simpson指数值都大于灌木层,但灌木层Pielou均匀度指数值都大于草木层。这说明在火烧迹地中,草本层比灌木层物种丰富,优势度和多样性高,但草本层群落分布没有灌木层群落分布均匀。
3 讨论与结论
3.1 讨论
在研究区火烧迹地上,随着坡位的升高,形成了唐古特忍冬+扁刺蔷薇-密生薹草群落、箭竹-密生薹草群落和黑水柳-野草莓群落3种典型群落。根据实际走访调查和查阅相关资料得知,黑水柳和野草莓主要生活在退化森林或受干扰的地区[26],而下坡位以黑水柳和野草莓为主要建群种,群落的个体数量相较于上坡位和中坡位都偏少,原因是下坡位比较平坦,适于藏民生活放牧和梅花鹿(Cervusnippon)、野猪(Susscrofa)等人为活动和野生动物活动,对下坡位灌木下层植被、枯落物厚度、土层厚度以及土壤密度等干扰较大[27],从而降低群落个体数量,而动物对黑水柳和野草莓的嗜口性较差,最后形成下坡位以黑水柳和野草莓为主要建群种格局[28]。动物虽对中坡位和上坡位主要建群种的嗜口性较好,如箭竹、密生薹草等,但由于中坡位坡度较陡,动物很难爬上山顶,局部小生境遭到的干扰较小,在不同坡位形成了生境异质性,土壤的水肥气热良好,为中坡位和上坡位的植物演替提供了很好的演替条件[29-30],从而使上坡位和中坡位群落的个体数量大于下坡位个体数量。干扰、坡位、坡度等环境因子是相同树种在不同坡位个体数量存在较大差异的重要原因,对于不同树种在同一坡位的个体数量差异较大,这主要是取决于群落物种间的竞争作用和自身的生物学特性。上坡位和中坡位灌木层多个树种重要值相差不明显,优势种多样,这主要还是由于受到的干扰较少,火烧迹地还正处于恢复初期,各树种获得的养分充足,种间竞争较小,优势树种呈现出多样化[31-32]。草本层则相反,下坡位的优势种多样,上坡位和中坡位优势种单一,原因是下坡位灌木层和草本层的演替受到了野生动物、放牧和人为活动等的干扰,林地上阳光和资源空间充足,适合草本植物生长,而上坡位和中坡位遭到的干扰较轻,整个灌木层群落盖度较大,草本层植物对光的利用率减小,喜阴草种逐渐代替喜阳草种,草本层演替达到相对稳定状态,上坡位和中坡位优势种较下坡位单一[33-35]。
研究区范围内植物资源约有60科123属314种[36],在火烧迹地上,共调查到27科49属60种植物,与未过火的林分相比,植被群落恢复15 a后,结构组成依然单一,火烧迹地出现了红桦更新先锋种。而针叶树种更新甚少,森林群落仍以灌木为建群种,这说明火烧迹地森林生态系统仍需很长的时间才能恢复到原有水平[12,37]。在森林恢复与重建的过程中,由于受到环境因子、气候变化、竞争作用、各种干扰等的影响,存在着很多的不确定性[38]。不同坡位的灌木层物种多样性与植物群落科、属和种组成的特征比例不同,均呈现出在上坡位和下坡位物种多样性指数值与群落组成特征比例都比中坡位高,这与前人研究的随着坡位的升高物种多样性降低的结果相反[39],这可能是由于上坡位坡度较小,地势较平坦,水土流失不严重[37,40],上坡位和中坡位流失的土壤元素和有机质都富集于下坡位,从而使下坡位的土壤比较厚且肥沃[37,41]。上坡位除了物种丰富度小于下坡位外,其余3种物种多样性指数均最高,这与上坡位的土层深度、枯落物厚度、各种干扰程度和生境异质性有关,造成下坡位物种多样性比上坡位低;研究区中坡位虽干扰不严重,但由于火烧迹地火后初期,植被较少,坡度较陡,可能导致水土流失比较严重,导致土层变薄,土壤养分降低[37,42],从而使中坡位灌木层植被物种丰富度和多样性较低。对于草本层,除Pielou均匀度指数外,物种丰富度和多样性指数基本随着坡位的升高而降低,物种多样性指数普遍大于灌木层,草本层物种多样性指数普遍大于灌木层,这是由于群落恢复15 a后仍处于初始演替阶段,草本物种数量增加最快,且很快达到高峰[37,43]。与灌木层形成较大的差异,形成这种格局的可能是草本层植物耐阴性、适应能力较强、生态位较宽等[44]。相较于草本层,灌木层可能受坡度、土层厚度,枯落物厚度、干扰因素和光竞争等因素影响较大。草本层群落Pielou均匀度指数比灌木层低,说明草本层群落各种群分布不均匀,灌木层群落进行正向演替,对水肥气热的竞争较大,进而使草本层植物分布不均匀。
除下坡位高度结构呈双峰分布以外,火烧迹地群落高度结构和径级结构均呈现出倒“J”型分布(即随着树高和径级的增加,群落个体数量现出先升高而后降低的变化趋势,株数减少幅度趋于平稳),较低矮和小径级植物数量占比较多,下坡位树种数量少于上坡位和中坡位,这说明下坡位可能长期受到动物活动的干扰,对于幼树的影响较大,降低幼苗数量,主要灌木树高和地径值较大;而中坡位主要由于坡度较大,上坡位和中坡位植被群落受到干扰较小,幼树数量较多,较低高径和小径级数量占比大于下坡位。各主要树种的高度结构和径级结构均呈现出倒“J”型分布、正态和多峰连续分散分布,种群呈倒“J”型分布,说明更新能力较强[45],个别树种呈正态分布或多峰分布且连续分散分布,说明该种群稳定性较高[23-25];对于整个火烧迹地灌木群落来说,总体呈倒“J”型分布,个别树种种群又接近于正态或多峰连续分散分布,说明目前还正处于正向演替阶段,更新能力较强,稳定性较高[23-25]。
3.2 结论
通过对火烧迹地恢复15 a后植被群落进行初步分析,植物种组成受植被生物学特性、干扰因子、地形因子、生境异质性等多因素的综合作用,在不同坡位形成小生境,不同坡位群落的类型、物种组成、群落结构和生物多样性存在着一定的差异。按不同坡位从上到下可依次划分唐古特忍冬+扁刺蔷薇-密生薹草群落、箭竹-密生薹草群落和黑水柳-野草莓群落3种典型群落。在这3种典型群落中,共调查到27科49属60种植物。其中上坡位调查到23科28属32种植物,中坡位调查到21科27属33种植物,下坡位调查到25科40属49种植物。除下坡位高度结构呈双峰分布以外,火烧迹地灌木群落高度结构和径级结构均呈现出倒“J”型。除Pielou均匀度指数外,草本层物种多样性指数总体优于灌木层;灌木层下坡位植物种丰富度最高,但本研究的其他多样性指数上坡位最高,草本层Shannon-Wiener指数随坡位上升而逐渐降低,Simpson指数中坡位最高,上坡位最低,Pielou均匀度指数中坡位最高,下坡位最低。
本研究对火烧迹地上不同坡位处的典型群落进行划分,分析不同坡位处群落结构、物种组成和生物多样性等,可为后续深入探讨重度火烧迹地群落恢复与重建、植物种内种间关系、群落物种组成与环境因子之间的关系、生物多样性与环境因子之间的关系等重要生态问题的研究打下基础,同时,为当地林业生态部门火烧迹地恢复与重建过程中的管理提供理论依据。
猜你喜欢
兰台内外(2021年33期)2021-12-17 15:41:02
中南林业科技大学学报(2020年4期)2020-04-26 13:15:56
花卉(2020年4期)2020-03-16 08:20:26
江西农业(2018年15期)2018-11-23 08:02:12
军民两用技术与产品(2018年11期)2018-07-16 11:49:24
现代农村科技(2018年6期)2018-01-18 06:09:22
山西建筑(2017年15期)2017-07-01 22:03:22
防护林科技(2017年5期)2017-06-15 18:14:30
绿色科技(2017年1期)2017-03-01 10:14:33
湖南林业科技(2017年1期)2017-02-06 05:29:18
