
局域种库对云南干热河谷植物多样性的影响*
2021-09-26 12:27:30刘方炎高成杰冯德枫李昆
西部林业科学 2021年5期
刘方炎,高成杰,冯德枫,李昆
(1.中国林业科学研究院资源昆虫研究所,云南 昆明 650233;2.云南元谋荒漠生态系统国家定位观测研究站,云南 元谋 651300)
传统观点认为,物种共存和生物多样性的维持是基于物种生态位分化的群落构建理论[1-3],竞争排除原理是这一理论的基础。在竞争激烈的植物群落中,不同物种之间通过对同一生态位的竞争而发生生态位分化,进而形成了多物种的稳定共存,这种稳定状态最终决定了群落的物种多样性;但在竞争较弱的植物群落中,物种生态位分化不明显,多物种仍然能够稳定共存,即用生态位分化理论无法很好地解释这一现象。因此,许多学者认为,将生态位分化理论与强调物种生态等价性的中性理论结合能较好地解释群落构建的机理,并强调了扩散和随机过程对群落中物种共存和多样性特征的影响[3]。
群落中物种共存和生物多样性的维持是进化、历史和生态尺度上的过程决定的[4]。越来越多的生态学家认为,区域生境中潜在的物种库(或称种库)在很大程度上对其起着决定作用,并由此提出了种库假说[5-10]。根据空间尺度的大小和实际操作的便利,种库主要分为两个层次,即局域(或区域)种库和实际种库[9-11]。局域(或区域)种库通常指可以进入某一特定群落的潜在物种数量,即“原”群落(null community)中物种的数量,它由地史过程和区域过程所决定。实际种库即对象群落中实际调查的物种数量,它主要由竞争、捕食和干扰等局域过程和局域(或区域)种库共同决定[4,10]。
种库假说认为群落内物种之间存在生态位的分化,发生在个体水平上的随机性和扩散作用在群落物种共存和生物多样性的维持上起着重要作用。目前,争论的关键点在于,区域过程(如物种的形成、迁移扩散和消亡等)和局域过程(如竞争、捕食和干扰等)在对象群落实际种库物种组成和多样性的形成过程中各自所占比例有多大[12]。如果区域过程(具有随机性)在对象群落物种共存和多样性维持过程中占主导作用,群落中实际种库的物种组成和区系成分与局域(或区域)种库不会存在明显差异;如果局域过程(具有非随机性)在对象群落物种共存和多样性维持过程中占主导作用,群落中实际种库的物种组成和区系成分将会与局域(或区域)种库存在明显差异。
干热河谷植被是我国植被中比较独特的一种干旱植物群落类型,呈现出群落结构单一、树木矮化而散生、旱生禾草发达的稀树灌木草丛状,具有非地带性、稀有性和脆弱性等特点。植物多样性的维护是该区域生态治理的一个重要方面。目前为止,不同植物群落实际种库的物种组成和区系特征与局域种库之间的关系尚缺乏基本的研究。本研究通过总结历年来的研究资料,比较分析了不同类型植物群落的物种组成与局域种库之间的关系,目的在于揭示干热河谷区局域种库在植物群落构建、物种组成和多样性维持上的重要性。
1 研究区域与方法
1.1 研究区概况
干热河谷位于我国西南地区,大致跨越6个纬度和5个经度(23°00′~ 29°00′N、98°50′~ 103°50′E)。区域地貌主要特点是大江、大河及其大支流两侧的高、中山峡谷地貌。本研究位于金沙江干热河谷最为典型地段——元谋坝区。全年降雨量634 mm,年干燥度3.30,在我国干旱气候分类系统中,属于半干旱类型。区域内降雨量的分配极为不均:雨季短暂(4~5个月),集中了全年降雨量的92 %,旱季漫长(7~8个月),极为干旱。年平均气温21.8 ℃,最热月平均气温27 ℃,最冷月平均气温14 ℃,日均温≥10 ℃天数大于350 d。区域土壤以燥红土为主,水稻(Oryzasativa)种植双季稻,一年三熟。气候类型与世界典型萨王纳地区极为类似。区域植被在外观上呈稀树草原状,具有独特的植物区系组成,常被称为“半萨王纳植被(semi-savanna)”或“河谷型萨王纳植被(savanna of valley type)”[13]。
1.2 研究方法
2005—2018年间,通过野外实地踏查完成了研究区内已监测到的种子植物物种的收录,并针对研究区内主要植物群落灌木层、草本层植物物种组成和植物多样性特征进行了调查和监测。样地设置和调查的具体方法为:在每种植物群落中设置6个10 m×10 m的大样方,调查群落内灌木层植物,记录物种名称、株丛数、冠幅、生长状况,以及样方的海拔、坡度、坡向、坡位、优势种更新特征等指标;采用相邻格子法把每个10 m×10 m的样方细分为4个5 m×5 m的样方,在5 m×5 m的样方正中间设置一个1 m×1 m的小样方调查群落内的草本层植物,记录物种名称、株(丛)数、平均高度及盖度,以及幼苗的名称、高度、数量等林下更新状况。不同植物群落基本概况见表1。
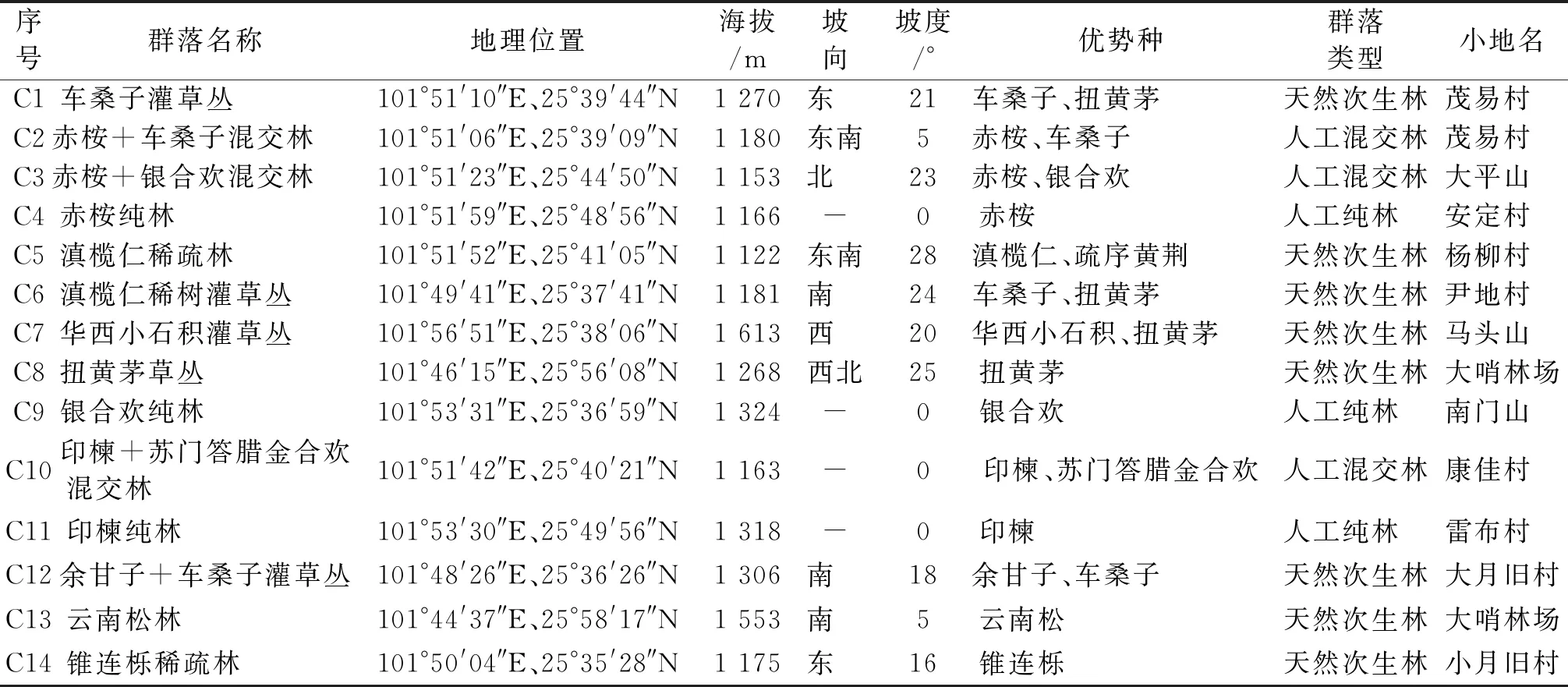
表1 不同植物群落基本概况
1.3 数据来源、处理与分析
本研究中使用的局域种库是在多年来实地调查收录的植物名录基础上,结合研究区历年来相关学者的研究结果[13-16]总结而成。种子植物分布区类型划分时,属的分布区类型采用吴征镒的标准[17]进行划分。植物群落与局域种库的属和种的分布特征以及区系特征采用皮尔逊相关性系数(Pearson Correlation Coefficient)进行比较。
2 结果与分析
2.1 局域种库中属和种的特征
局域种库中,种子植物属和种的数目最多的3个科均为禾本科(Gramineae)、蝶形花科(Fabaceae)和菊科(Compositae)。除此之外,唇形科(Labiatae)、大戟科(Euphorbiaceae)、爵床科(Acanthaceae)、玄参科(Scrophulariaceae)和莎草科(Cyperaceae)等种子植物属的数目相对较多;莎草科、锦葵科(Malvaceae)、唇形科、大戟科、苏木科(Caesalpiniaceae)和蓼科(Polygonaceae)等种子植物种的数目相对较多。由于研究区内单种属植物较多,有17个科种子植物属的数目和种的数目均排在前20的科中(图1、图2)。从属的分布区类型看,局域种库中热带分布的属占绝对优势,合计达70%以上(图3)。
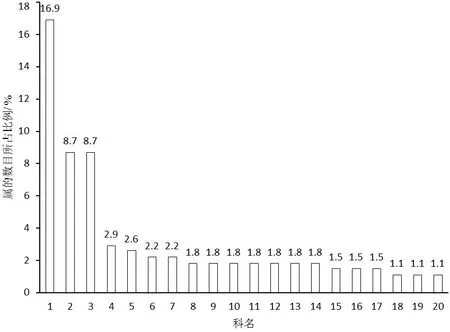
图1 局域种库中属的数目最多的前20科

图2 局域种库中种的数目最多的前20科
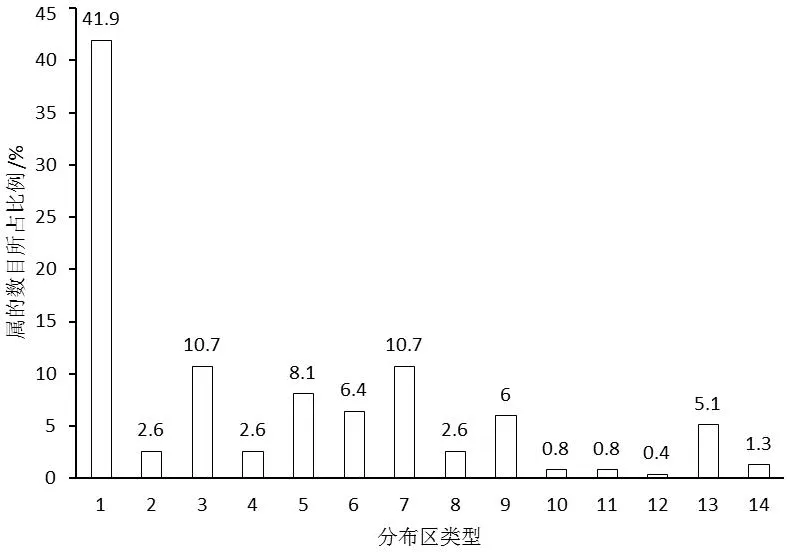
图3 局域种库中种子植物属的分布区类型(数据引自欧晓昆,1988)[16]
2.2 实际种库中属的组成特征
不同植物群落实际种库中属的组成特征存在较大差异(图4)。在人工恢复植物群落中,除了印楝(Azadirachtaindica)纯林(C11)外,其它群落中,包括赤桉(Eucalyptuscamaldulensis)+车桑子(Dodonaeaviscosa)混交林(C2)、赤桉+银合欢(Leucaenaleucocephala)混交林(C3)、赤桉纯林(C4)、银合欢纯林(C9)、印楝+苏门答腊金合欢(Acaciaglauca)混交林(C10)均为禾本科或蝶形花科植物在属的组成上占有绝对优势;印楝纯林中,除了蝶形花科植物外,菊科植物也在属的组成上占有较大比例。不同天然次生植物群落中,禾本科和蝶形花科植物所占比例仍然较高,但属的数目排在前3位的科呈现多样化的特点,除了禾本科、蝶形花科和大戟科外,还有唇形科、菊科、莎草科,表明天然次生植物群落在属的组成上单个科植物具有绝对优势的现象有所减弱,呈现出更加多元化的特点。
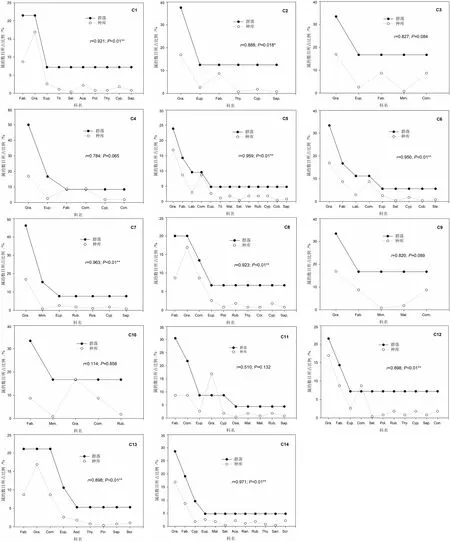
图4 局域种库和实际种库(植物群落)中属的组成特征比较
从不同植物群落实际种库与局域种库在属的组成上的比较可知,人工恢复植物群落中,除了赤桉+车桑子混交林(C2)的实际种库与局域种库之间呈显著相关(P<0.05)外,其它植物群落的实际种库与局域种库之间均没有显著相关性(P>0.05);天然次生植物群落中,所有群落的实际种库均与局域种库呈极显著相关(P<0.01)。表明局域种库对大多数植物群落实际种库属的组成特征具有显著或极显著影响。
2.3 实际种库中种的组成特征
不同植物群落实际种库中种的组成特征与属的组成特征无明显差异(图4、图5),实际种库中种的数目所占比例与属的数目所占比例基本保持一致。表明实际种库中单种属植物所占比例较高。
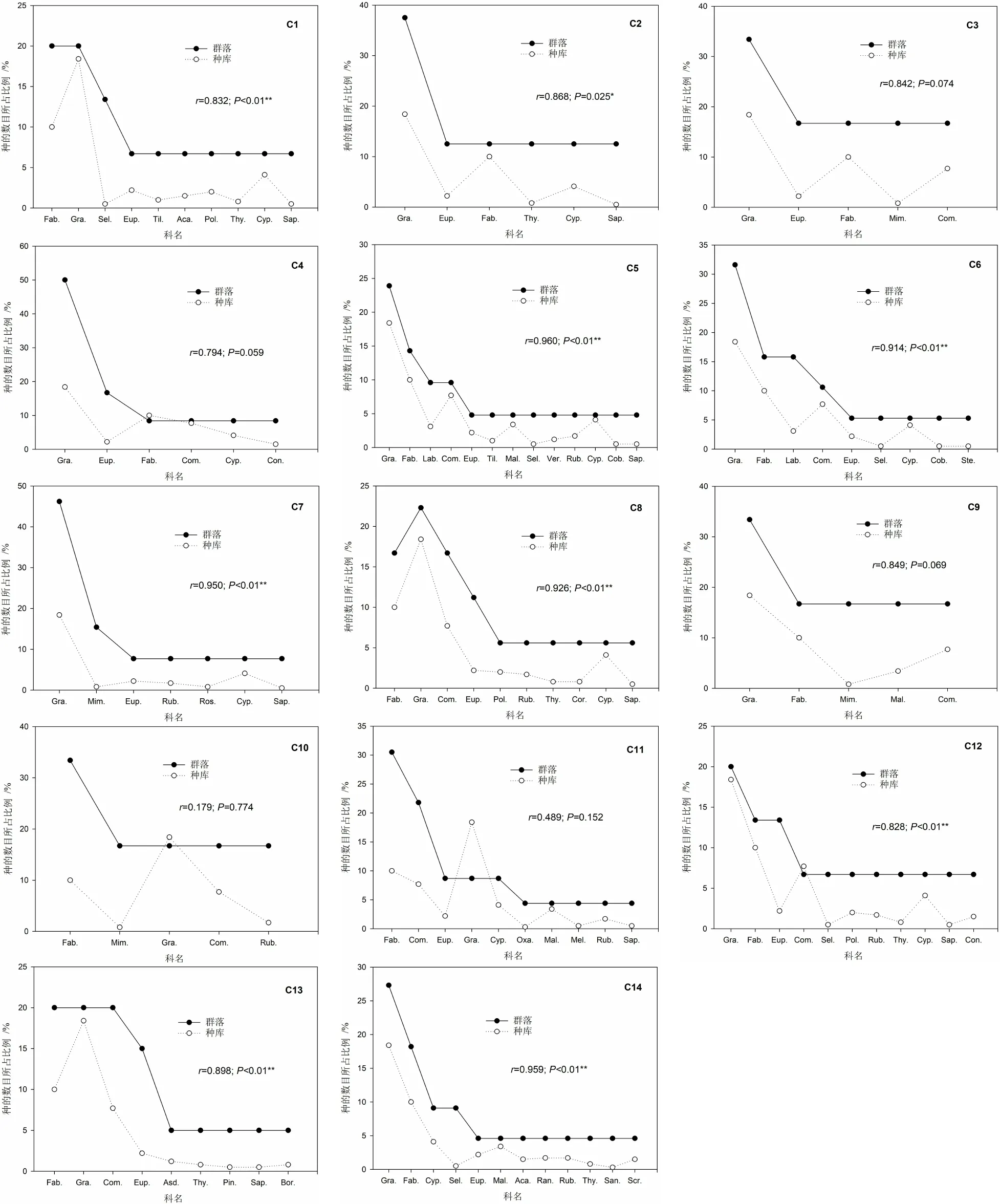
图5 局域种库和实际种库(植物群落)中种的组成特征比较
从实际种库与局域种库在种的组成上的比较可知,除了赤桉+银合欢混交林(C3)、赤桉纯林(C4)、银合欢纯林(C9)、印楝+苏门答腊金合欢混交林(C10)和印楝纯林(C11)等人工恢复植物群落外,其它植物群落实际种库种的组成特征均与局域种库呈显著(P<0.05)或极显著(P<0.01)相关。表明局域种库对天然次生植物群落和少量人工恢复植物群落实际种库种的组成特征具有显著或极显著的影响。
2.4 实际种库中属的分布区类型
不同植物群落实际种库属的分布区类型中,除了世界分布类型以外,热带分布成分均占有绝对优势,但不同植物群落属的分布区类型存在较大差异(表2)。人工恢复植物群落中,除了印楝纯林(C11)外,其它植物群落,包括赤桉+车桑子混交林(C2)、赤桉+银合欢混交林(C3)、赤桉纯林(C4)、银合欢纯林(C9)、印楝+苏门答腊金合欢混交林(C10)实际种库属的分布区类型均少而集中,天然次生植物群落中植物属的分布区类型相对较多,但与局域种库相比,仍然不完整。研究结果表明:不同植物群落实际种库中的物种丰富度仍有很大的上升潜力。
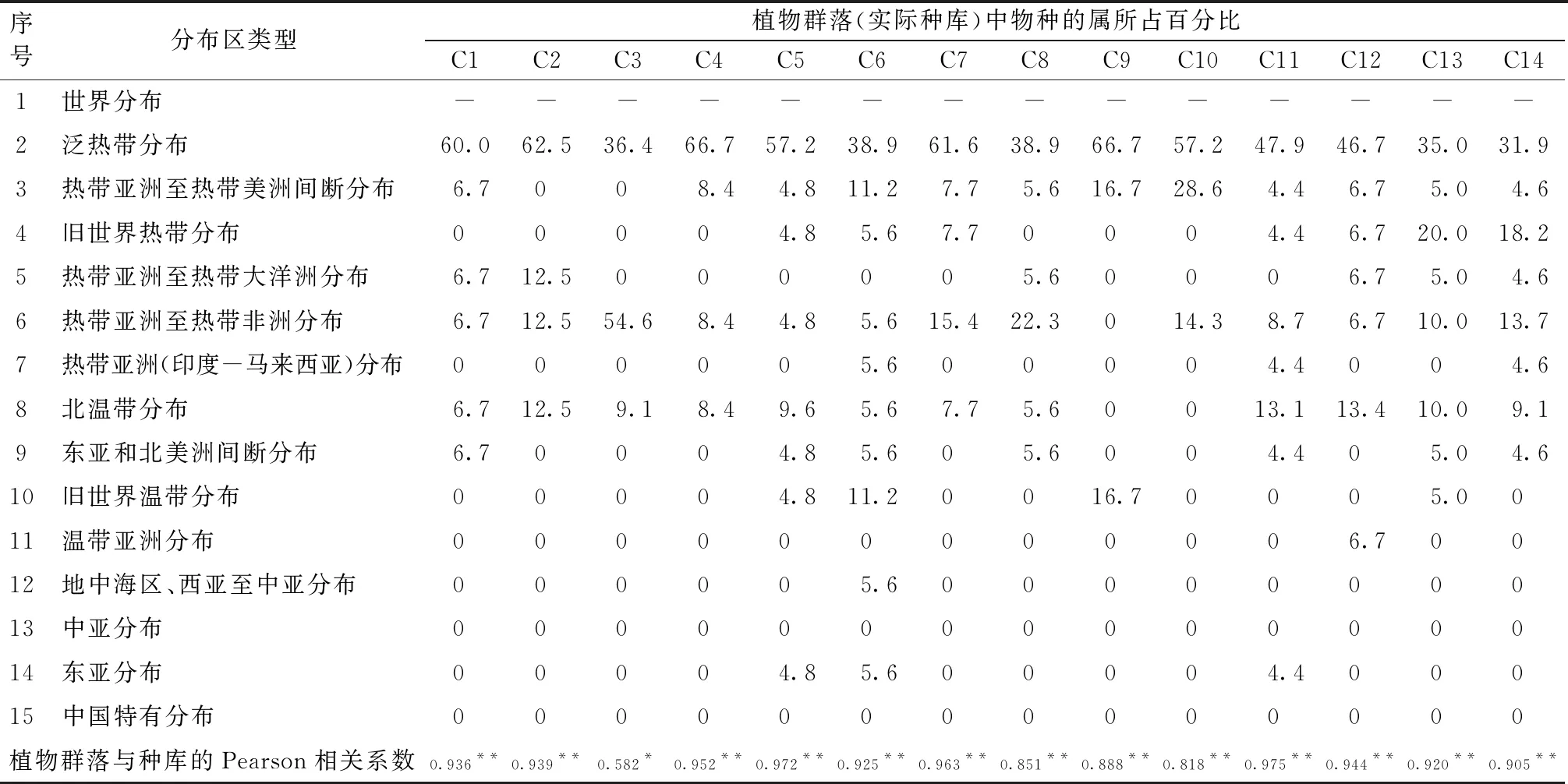
表2 局域种库和实际种库中属的分布区类型比较
Pearson相关性分析结果显示,不同类型植物群落的实际种库在属的分布区类型上与局域种库均呈显著(P<0.05)或极显著(P<0.01)相关。结果表明局域种库对植物群落实际种库属的分布区类型具有显著或极显著影响。
3 讨论
3.1 局域种库特征及其代表性
在研究局域种库与对象群落(实际种库)植物种类组成和多样性特征的关系方面,种库的确定是最为困难的。根据空间尺度的大小,种库可以分为区域种库、局域种库和实际种库[11]。局域种库是指分布在某一局域尺度上(如某一特定流域等)、能够在对象群落中潜在共存的一群物种[4,10]。区域种库涉及的范围和面积更大,但在实际操作过程中,很难明确区域种库和局域种库的界限。这也是大多数学者没有将这一潜在共存种库进行细分的重要原因[10,18-21]。因此,在研究过程中,首先应该确定的是种库涉及的区域范围。涉及面积过大,地形、地貌以及气候差异也会较大,种库不具有典型的代表性。故无论将这一种库称作局域种库还是区域种库,只要它涉及的范围或表示的类型可以代表某一群落中能够潜在共存的一群物种即可。本研究中涉及的植物群落均位于元谋坝区海拔1 600 m以下的干热河谷区,在地貌特点和气候指标上明显不同于四周的高原环境,使用金沙江干热河谷典型地段——元谋坝区作为局域种库的研究范围,具有较强的代表性。
通常,采用不同的方法明确区域内完整的植物区系资料和植物分布的环境信息,是确定一个区域植物种库的必需环节。研究表明,利用Braun-Blanquet 法瑞植物社会学方法确定植物种库的效果好于生态学的方法(基于物种的Ellenberg指数)[10]。干热河谷植被是我国极为特殊的植被类型,一直受到学术界的广泛关注。Braun-Blanquet 法瑞植物社会学学派作为植物生态学研究的四大学派之一,其理论和方法也被广泛用于干热河谷植被的研究。目前,作为金沙江流域最为典型的干热河谷区,元谋坝区的植被类型、植物区系特征、生态环境状况、特征种、标志种、广布种和特有种均已非常明确[13-16]。本研究中使用的局域种库,是在多年研究过程中收集整理的物种库基础上,结合研究区内已有的研究成果总结而成,相对准确和完整,具有一定可信度。
3.2 局域种库与实际种库之间的关系
局域种库大小主要由大尺度的地史过程(如地球气候变化)和区域过程(如物种的形成、灭绝、迁入和迁出)所决定。生境类型的面积越大、地质年代越古老,局域种库就越大,实际群落中物种丰富度就越高[10]。局域种库对植物群落物种共存和物种丰富度(即实际种库大小)具有重要影响。但由于植物群落所处的环境不同,实际种库除了受到局域种库限制外,还受局域过程,如竞争、捕食和干扰等因素的严重影响,因此,不同立地条件下植物群落的实际种库与局域种库之间的关系是不同的。而局域过程对实际种库的相对重要性在不同环境条件下也是存在差异的[11]。研究表明,当植物群落内物种竞争较弱时(空余生态位充足),实际种库随局域种库的增加而增加;当群落内物种竞争激烈时(空余生态位缺乏),实际种库大小上升到一定程度后将达到饱和而不再上升[22]。本研究中,在属和种的组成特征方面,天然次生植物群落和极少量人工恢复植物群落的实际种库与局域种库存在显著或极显著相关。表明在这些植物群落的实际种库大小受到局域种库的显著或极显著影响。此时,若群落内空余生态位充足,其物种丰富度和植物多样性仍有较大上升空间;若空余生态位缺乏,其物种丰富度和植物多样性可能已经达到饱和状态而不再上升了。其具体情况有待进一步研究。另一方面,大多数人工恢复植物群落的实际种库与局域种库并没有显著相关性。表明在这些植物群落中实际种库大小并没有受到局域种库的显著影响,受到局域过程,如竞争、捕食和环境变动(干扰)等因素的影响可能更加严重。在属的分布区类型上,所有植物群落的实际种库均与局域种库呈显著或极显著相关,表明研究区内所有植物群落的实际种库在植物区系特征上与局域种库均是一致的,这与整个研究区生态环境密切相关。
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:18
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
科学大众(中学)(2019年3期)2019-05-17 10:04:30
电子测试(2018年18期)2018-11-14 02:30:34
汽车观察(2018年10期)2018-11-06 07:05:26
中国医学装备(2015年10期)2015-12-29 12:00:24
少儿科学周刊·少年版(2015年1期)2015-07-07 17:15:12
电测与仪表(2015年7期)2015-04-09 11:39:50
水生生物学报(2015年1期)2015-02-28 16:01:05
