
嵌有非天然氨基酸蛋白质的生物合成及其纯化鉴定
2021-09-24 02:42马韩轲李闰婷张丽萌裴若兰陈肖皖陈龙欣
河南科学 2021年8期
马韩轲, 李闰婷, 张丽萌, 张 函, 裴若兰, 陈肖皖, 陈龙欣
(1.郑州师范学院分子生物学实验室,郑州 450044; 2.郑州师范学院生命科学学院,郑州 450044)
蛋白质是生命活动中不可或缺的基本组成部分,它具有催化、运输、调节、免疫以及构成生物体等多种功能. 绝大多数蛋白质是由二十种天然氨基酸构成,为了进一步拓展蛋白质的功能,研究人员开始逐渐将视角转到可控和易操作的天然氨基酸侧链衍生物——非天然氨基酸(Unnatural amino acids,UAAs)上[1]. 非天然氨基酸特异性强、对蛋白结构扰动小、灵敏度高、使用灵活,通过非天然氨基酸对蛋白质进行修饰,可实现蛋白质的新功能,对其结构和功能的理论研究与应用带来了新的契机[2]. 为了在肽链或蛋白质中引入携带独特官能团的非天然氨基酸,人们不断探索多样的化学合成方法,但由于蛋白质分子量过大,化学合成往往并不容易,而基于密码子扩展技术[3],可以通过简单的生物合成,在蛋白质中定点引入非天然氨基酸,这无疑为蛋白质的生物化学研究和蛋白质操纵提供了全新的技术手段[4]. 该技术最初由Scripps研究所的Peter G.Schultz教授团队建立并推广[3,5-6]. 它的出现打破了自然界中生物体内二十种常见氨基酸的编码限制,使得在蛋白中扩展了更多功能基团[7-8].
目前,基于密码子扩展技术,利用非天然氨基酸作为重要工具在活细胞中对蛋白进行了标记和研究,比如通过“点击”化学修饰可以高效地进行蛋白标记[9],通过显色基团可以进行光谱分析[10],通过荧光基团可以优化超高显微观测[11]等. 近期,利用基因密码子扩展技术制备复制缺陷型流感病毒,有望在新型疫苗领域有所突破[12],甚至在不需要外源供给复杂的非天然氨基酸的情况下,通过大肠杆菌中半胱氨酸的生物合成途径“劫持”半胱氨酸合成酶,便可以实现几十种新型非天然氨基酸嵌合到重组蛋白中[13]. pBpa(p-苯甲酰-L-苯丙氨酸)是一种在苯丙氨酸侧链连接有光敏感基团的非天然氨基酸[14-16],最近,通过光敏型非天然氨基酸pBpa插入蛋白质的功能表位,更是拓展了抗体药物筛选领域[17].
在本研究中,我们基于基因密码子扩展技术,将非天然氨基酸pBpa通过生物合成的方式,插入到白细胞介素-1β(IL-1β)蛋白质的预设位点中,能够较为容易地获得大量的具有特定功能的非天然氨基酸嵌合体蛋白质.
1 材料与方法
1.1 材料
pET28a原核表达载体质粒、pEVOL-pBpaRS 质粒由郑州师范学院分子生物学实验室保存;非天然氨基酸pBpa购自J&K Scientific,货号204322(CAS104504-45-2);大肠杆菌BL21(DE3)和DH5α化学感受态细胞以及质粒小提试剂盒购自天根生化科技(北京)有限公司;DNA Maker购自北京全式金生物技术有限公司;蛋白质Marker购自Thermo Fisher Scientific;IPTG、阿拉伯糖购自AMERSCO公司;胶回收试剂盒购自ZOMANBIO公司.
1.2 方法
1.2.1 表达工程菌的构建 在编码的IL-1β第5个氨基酸精氨酸的质粒编码位点CGG突变为琥珀密码子碱基TAG(在本研究中TAG 编码pBpa),将IL-1β突变基因进行人工合成(苏州金唯智生物科技有限公司),合成后的基因连接到pET28a载体中,转化到DH5α感受态细胞,涂布于含有卡那霉素(50 μg/mL)的2×YT固体培养基(16 g/L胰蛋白胨,10 g/L酵母浸粉,5 g/L氯化钠,15 g/L琼脂)的平皿中,37 ℃过夜倒置培养,挑取多个单克隆测序验证,测序正确的克隆质粒命名为pET28a-IL-1β-R5TAG,小量提取质粒备用.
将pEVOL-pBpaRS 载体[18](含有编码正交的RNA 合成酶和tRNA 正交对的元件,能将非天然氨基酸pBpa 氨酰化运载于tRNA 上[19-20]),以及产生R5 位点突变的IL-1β质粒pET28a-IL-1β-R5TAG 共同转化到BL21(DE3)感受态细胞中,涂布于含有卡那霉素(50 μg/mL)和氯霉素(15 μg/mL)双抗性的2×YT 固体培养基平皿上,37 ℃过夜倒置培养,单克隆便为能够生物合成嵌合有非天然氨基酸pBpa突变体蛋白质的工程表达菌株.
1.2.2 非天然氨基酸嵌合蛋白的表达 用含有卡那霉素和氯霉素双抗性的2×YT液体培养基(不含琼脂)复壮工程表达菌株,将其培养至菌的吸光度OD600约为0.6后,加入0.22 μm除菌的质量体积比为0.2%的阿拉伯糖、1 mM pBpa以及0.5 mM IPTG,37 ℃250 r/min振荡培养5 h形成嵌合蛋白的可溶性表达,5000×g离心收集菌体沉淀,而后沉淀悬溶于PBS缓冲液中,冰上超声破碎(超声5 s停7 s)20 min,10 000×g离心收集超声破碎后的上清溶液,上清便含有R5pBpa的嵌合蛋白. 同时,表达时设置不加入非天然氨基酸pBpa的培养物作为阴性对照. 最后,SDS-PAGE实验鉴定嵌合蛋白的表达.
1.2.3 非天然氨基酸嵌合蛋白的纯化 将超声破碎的上清分多次加入Ni-NTA 纯化介质柱中(金斯瑞生物
科技有限公司L00250-C),20倍柱体积的洗涤缓冲液(含有20 mM咪唑的PBS溶液)洗涤纯化介质柱,最后用250 mM的洗脱缓冲液(含有250 mM咪唑的PBS溶液)洗脱纯化介质柱中的嵌合蛋白R5pBpa. 为了进一步纯化嵌合蛋白,将其通过AKTA蛋白纯化仪的Superdex75 Increase 10/300 GL分子筛按照1 min/mL的流速进行精细纯化,收集8~15 mL上的主峰流出液,SDS-PAGE实验鉴定纯化质量.
1.2.4 非天然氨基酸嵌合蛋白的质谱鉴定 在安捷伦6530 电喷雾四极飞行时间质谱仪前端串联安捷伦1290 高效液相色谱仪,将10 μL 1 μg/μL 的精细纯化的嵌合蛋白通过高效液相色谱仪装载到反相柱(300SB-C8,2.1 mm×50 mm,3.5 μm颗粒)上,然后在流速为0.2 mL/min 的梯度相上洗脱分离后,进入电喷雾电离在线引入ESI-Q-TOF质谱[17]. 最后收集的质谱数据通过安捷伦公司的MassHunter BioConfirm软件按照其工作流程进行分析.
2 结果与分析
2.1 嵌合蛋白的表达
工程菌具有两种抗性,说明双质粒工程菌成功构建. 表达R5pBpa嵌合蛋白的工程菌经IPTG及阿拉伯糖的共同诱导,在有和无非天然氨基酸pBpa两种情况下,表达5 h 后,通过SDS-PAGE 实验对全菌体进行检测,如图1 所示,在加入pBpa 的样品中,15~25 kDa 处有明显条带,而未加入pBpa 样品中没有发现明显条带,符合嵌合蛋白约19 kDa的理论分子量. 因此,本实验构建的双质粒工程菌在非天然氨基酸存在的条件下,能够表达R5pBpa嵌合蛋白.
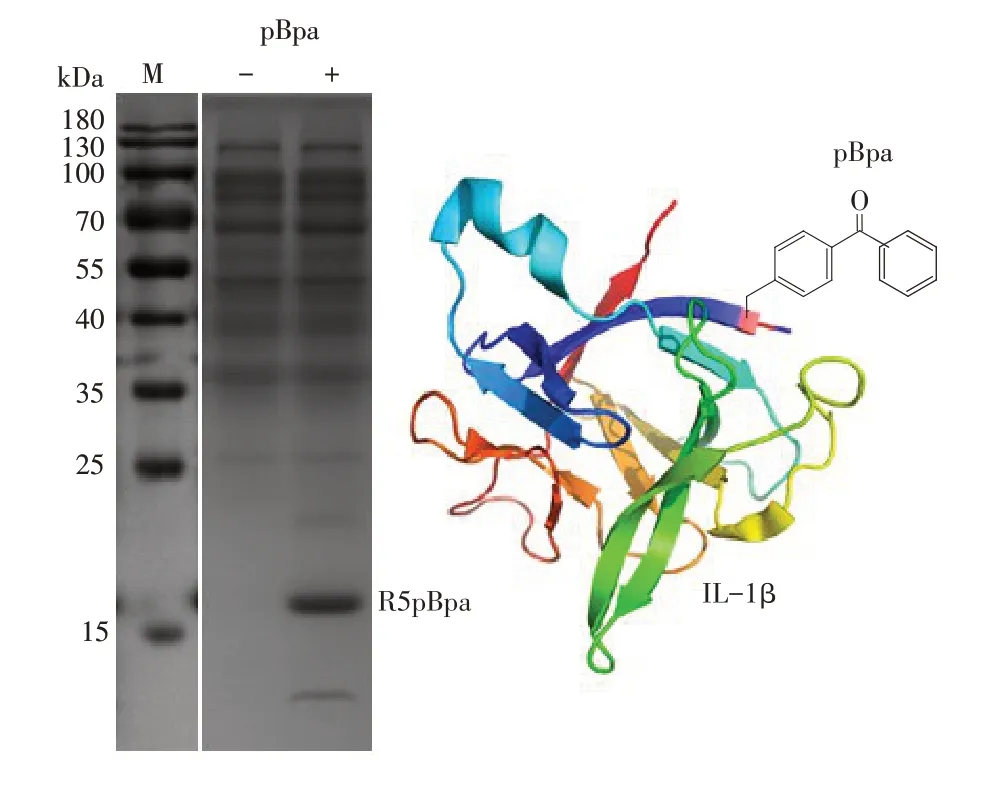
图1 全菌体的SDS-PAGE结果及其R5pBpa嵌合蛋白的模拟图Fig.1 SDS-PAGE results of whole cell and the pattern of chimeric protein R5pBpa
2.2 嵌合蛋白的纯化
含有His标签的IL-1β的R5pBpa嵌合蛋白通过Ni-NTA 纯化介质初步纯化后,获得一定纯度的R5pBpa,如图2 中的1 泳道,工程菌表达R5pBpa 的产量约为30 mg/L. 为了进一步提高纯度,用于后期的精密实验,根据蛋白的大小利用分子筛进行了精细纯化,收集峰中的蛋白溶液,如图2 所示,蛋白的主峰出现在出峰体积为12.5~15 mL 处,符合分子量约为19 kDa 的出峰时间,将主峰收集的5 管蛋白溶液依次SDS-PAGE上样,如图2的2~6泳道. 主峰中获得了相当高纯度的非天然氨基酸pBpa嵌合IL-1β蛋白R5pBpa.
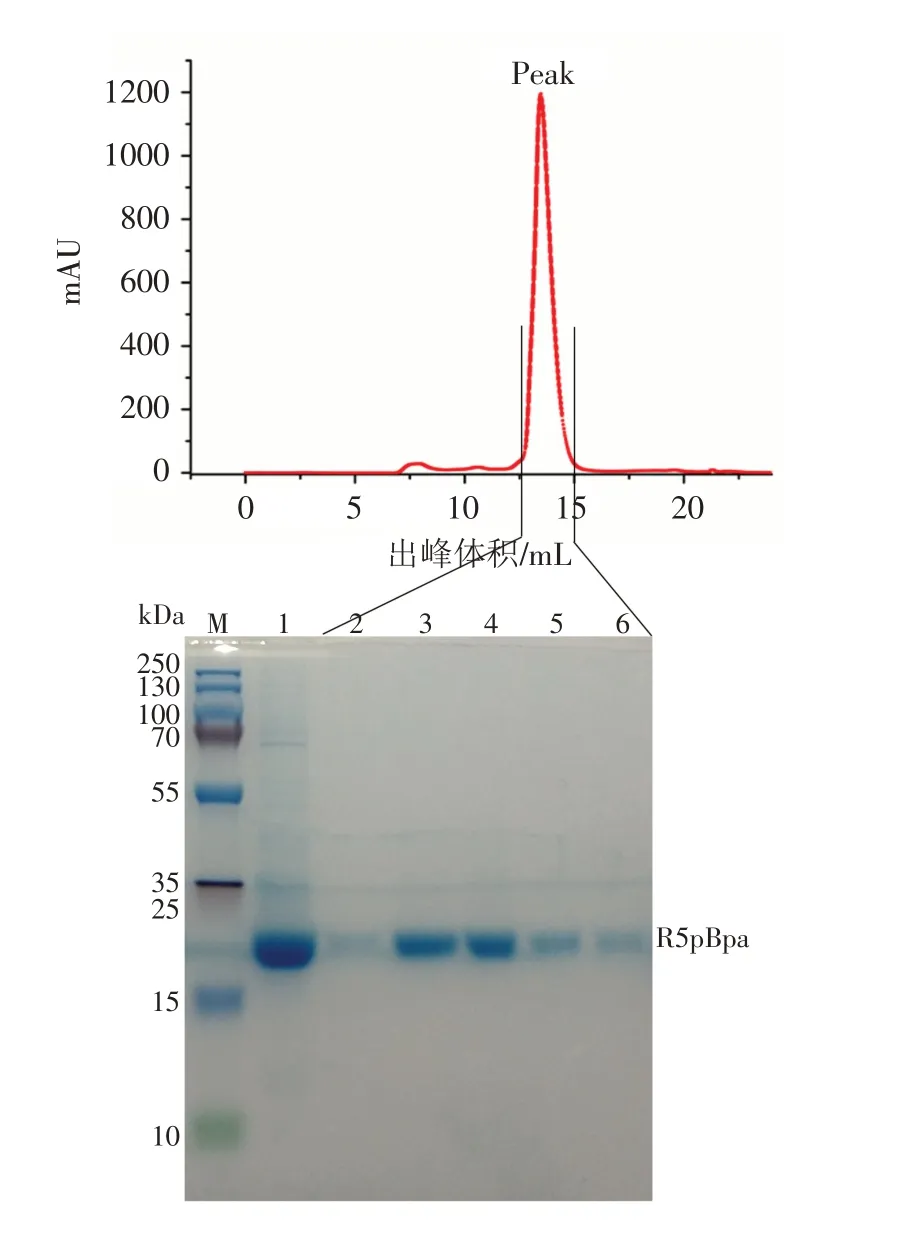
图2 分子筛精细纯化峰图及其对应的SDS-PAGE结果Fig.2 Purification peak diagram of molecular sieve and its corresponding SDS-PAGE results
2.3 嵌合蛋白精准分子量的质谱鉴定
为了进一步确认非天然氨基酸pBpa确实插入到了IL-1β蛋白中,我们对纯化的嵌合蛋白进行了精准分子量ESI-Q-TOF质谱鉴定. 相比野生型IL-1β而言,突变体R5pBpa 中的pBpa 非天然氨基酸残基取代了野生型蛋白的第5 位精氨酸残基,使得嵌合蛋白分子量比野生型蛋白分子量更大一些. pBpa 取代精氨酸R后两者相对分子质量之差为95.1 Da,因此在野生型IL-1β分子量(18 809.45 Da)的基础上加上两者之差即为R5pBpa的理论分子量,Expected mass=18 904.55 Da,如图3 所示,实际测得的主峰中突变体R5pBpa 分子量为Observed mass=18 905.76 Da,符合理论分子量的数值,表明通过本实验可以获得较为纯净的pBpa嵌合蛋白.

图3 ESI-Q-TOF质谱精准分子量鉴定结果Fig.3 Accurate molecular weight identification by ESI-Q-TOF mass spectrometry
3 结语
目前,越来越多领域都出现了非天然氨基酸的身影,非天然氨基酸由于其特殊性可根据人们的需求被赋予特定的功能,从而为诊断、疾病的治疗等多个领域提供崭新思路[21]. 在本研究中,我们只需要将两个质粒(一个质粒为提供识别非天然氨基酸pBpa的氨酰-tRNA合成酶/tRNA正交对的pEVOL-pBpaRS质粒,另外一个质粒为在含有编码目标蛋白的基因的特定任意位点上设置琥珀密码子供非天然氨基酸的插入)转化到大肠杆菌感受态中,便可以简单而高效地表达插入有特定基团的非天然氨基酸到蛋白质中,在中性条件下,通过常规的蛋白质纯化便可以获得大量纯度极高的具有生物活性的蛋白质. 另外,在研究其他感兴趣的蛋白质时,我们也可很容易地改变质粒编码的基因和位点,来实现pBpa对其他功能蛋白的嵌入,更进一步的,作为研究或应用的工具,我们甚至可以较为容易地在pEVOL骨架质粒上突变氨酰-tRNA合成酶/tRNA正交对,实现其他非天然氨基酸的嵌入.
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
石油沥青(2022年2期)2022-05-23
福建农业学报(2021年6期)2021-08-18
纺织科学研究(2021年7期)2021-08-14
江西农业学报(2021年4期)2021-04-20
教学考试(高考生物)(2020年4期)2020-11-18
生物学通报(2020年11期)2020-10-22
三农资讯半月报(2020年11期)2020-06-21
海峡科技与产业(2019年4期)2019-10-26
