
鸡CEBPA基因CDS区克隆、表达及生物信息学分析
2021-09-14 04:37杜振伟朱帅鹏马向飞李东华孙桂荣
生物技术通报 2021年8期
杜振伟 朱帅鹏 马向飞 李东华 孙桂荣
(河南农业大学动物科技学院,郑州 540046)
CEBPA是碱性亮氨酸拉链蛋白家族的一个亚家族成员中最早发现的[1-2].,该增强子结合蛋白家族在脂肪细胞生长、分化、骨髓组织细胞生成及机体免疫过程中发挥重要作用[3],同时在脂肪的生长过程中起正向调和反向调节作用。CEBPA被认为是调节脂肪形成的关键分子之一[4-5],在脂肪细胞的终末分化中有着很重要的作用[6]。牙髓衍生的间充质干细胞(DPSc)沿成骨和成软骨途径分化时,因DPSc不含脂泡,检测到CEPPA的表达水平降低[7]。在特发性肺纤维化(IPF)细胞中过表达CEBPA可增加脂肪生成潜能和脂肪成纤维细胞标记物的表达,抑制CEBPA的表达会降低了成脂潜能并促进了成纤维细胞的活化[8]。全反式维甲酸(atRA)可通过RARG-FRA1-PPARG2或CEBPA轴或两者抑制骨髓间充质干细胞(BMSC)的脂肪形成[9]。白藜芦醇通过使PPARG、CEBPA、FABP4和LMO3的表达下调来抑制脂肪分化[10]。CEBPA增强了Slc2a4基因和GLUT4蛋白的表达,是脂肪细胞分化的标志[11]。Gao等[12]研究表明CEBPA 的启动子区 CpG 位点甲基化使瘦肉系鸡显着高于脂肪系鸡,且启动子区域-1 494 bp - -1 478 bp 的 CpG CEBPA mRNA 表达呈显著负相关,这就表明该基因在脂肪组织中发生甲基化并调控脂肪发育。CEBPA与脂肪分解、脂肪生成和脂肪酸饱和相关基因聚在一起[13],同时秦川牛的脂肪沉积也与CEBPA有关[14]。本实验通过对CEBPA基因编码序列进行生物信息学分析,并测得其在不同组织,不同发育时期及不同品种的表达量且加以分析,为以后CEBPA基因在胸肌脂肪沉积方面的研究提供了理论基础。
1 材料与方法
1.1 材料
本试验的试验对象为固始鸡和罗斯鸡,均由河南农业大学家禽种质资源场提供。分别取6、12、22、30、55周龄的健康固始鸡各3只和6周龄罗斯鸡3只,置于相同的环境下饲养一段时间后,进行颈部放血法处死试验动物后,迅速采集胸肌、肝脏、腿肌、皮下脂肪,腹脂组织并及时用液氮进行冷冻,并置于-80℃保存备用。本研究通过河南农业大学科学伦理委员会批准(批准号20-0086)。
1.2 方法
1.2.1 总RNA的提取与反转录 使用TRIzol法提取固始鸡胸肌、肝脏、腿肌、皮下脂肪,腹脂以及罗斯肉鸡胸肌的总RNA,并用反转录试剂盒制备cDNA,放入-20℃冰箱保存以备后用。
1.2.2 鸡CEBPA CDS区的克隆 根据NCBI上鸡(NP_001026630.1)的CEBPA基因的CDS区,在Oligo 7上设计克隆引物见表1,以肝脏的cDNA模板。PCR反应体系:cDNA 4 μL,上、下游引物各 6 μL,KOD OneTMPCRMaster Mix 100 μL,加水至200 μL。PCR反应条件:98℃ 3 min;98℃ 10 s,70℃ 5 s,68℃ 15 s,15个循环;98℃ 10 s,55℃ 5 s,68℃ 15 s,25个循环;68℃ 10 min。反应结束后,为DNA平末端加A尾,再向PCR产物中添加约体系1/3的 2×Rapid Taq Master Mix,震荡混匀,再放入PCR仪中,72℃ 10 min。反应结束后,将PCR扩增产物进行1.5%琼脂凝胶电泳检测,并将目的片段进行切胶回收,用回收产物与pMDTM18-T Vector Cloning Kit连接,16℃ 16 h后转化DH5α感受态细胞,涂板,37℃培养,挑选阳性菌落,经菌液PCR鉴定,送上海生工生物公司测序。
1.2.3 生物信息学分析 利用TMHMM和SignalP 5.0在线软件分析其跨膜结构域与信号肽;运用PSIPRED和 SWISS-MODE在线软件对其蛋白的二级,三级结构进行预测;利用PSORT II Prediction在线软件对蛋白进行亚细胞定位;用DNAStar软件对鸡(NP_001026630.1)、小鼠(NP_001274443.1)、人(NP_001272758.1)、猪(XP_003127063.1)、牛(NP_789741.2)、家兔(XP_008255494.1)、日本鹌鹑(XP_015729433.1),斑马鱼(NP_571960.1)的氨基酸序列进行同源性分析;用 MEGA 7.0 软件构建CEBPA蛋白物种进化树。并用STRTNG进行蛋白互作分析,相关参数设置为:最小相关得分为0.40,一级最大相关因子数为10。在NCBI网站中搜索鸡CEBPA基因的核苷酸序列,以其转录起始位点为界限,截取基因序列上游-2 000 bp - -1 bp共2 000 bp序列作为CEBPA基因5'调控区进行序列分析,并利用在线软件BDGP:Neural Network Promoter Prediction,Promoter 2.0和FPROM对 鸡CEBPA基因5'调控区的启动子进行预测和分析,同时用在线软件EMBOSS和MethPrimer对其启动子区进行甲基化CpG岛预测,相关参数设置为:CpG岛长度>200 bp,CpG检测含量/期望含量(Obs/Exp)>0.60,GC%>50%。
1.2.4 荧光定量PCR 该反应以得到的cDNA为模板,采用SYBR GreenⅠ染料法和LightCycler96实时荧光定量PCR仪,按照试剂盒中的操作说明,以β-action作为内参基因,检测目的基因的表达量。qRT-PCR反应体系为10 μL,其中TBGreen Mix 5 μL,无RNA酶水 3 μL,上、下游引物(表1)各0.5 μL,cDNA 1 μL。qRT-PCR扩增反应程序:95℃ 5 min;95℃ 30 s,60℃ 30 s,72℃ 30 s,40个循环;95℃ 10 s,65℃ 60 s,97℃ 1 s;37℃ 30 s。每组实验包含2个技术重复和6个生物重复。
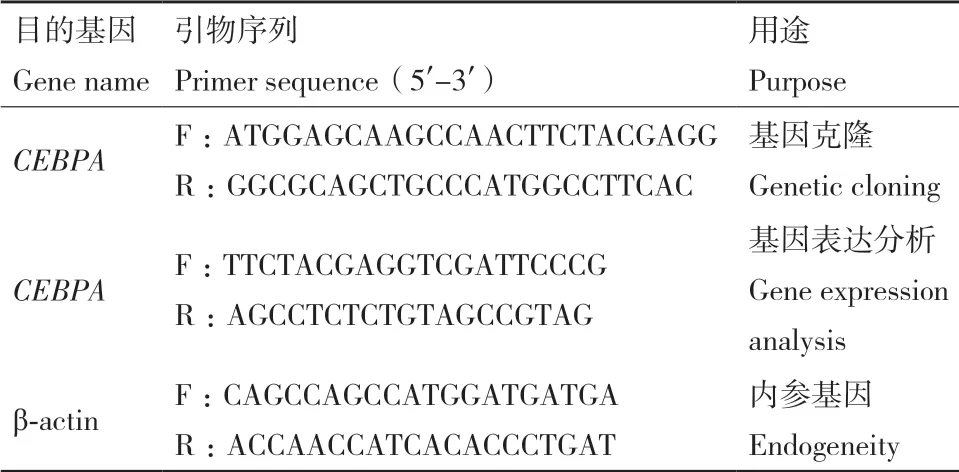
表1 基因引物序列Table 1 Gene primer sequences
1.2.5 实时定量数据分析 采用SPSS 23.0软件单因素分析方法,利用Duncan法进行差异显著性检验。P<0.01表示差异具有极显著性,P<0.05表示差异具有显著性。数值表示为平均数±标准差,利用GraphPad Prism5.0软件作图。
2 结果
2.1 鸡CEBPA CDS区的克隆
以鸡肝脏的cDNA为模板,通过PCR扩增用琼脂凝胶电泳检测目的片段以及阳性菌落测序(图1)。该片段为CEBPA基因的CDS区的全长(GenBank登录号:NC_006098.5),975 bp,共编码氨基酸325个,位于Chromosome 11:10295396-10296370,一个外显子。
2.2 鸡CEBPA的生物信息学分析
2.2.1 CEBPA蛋白结构和功能的预测 预测CEBPA蛋白结构表明,该蛋白不存在跨膜域,且无信号肽(图2-A,2-B),有211个氨基酸组成的无规则卷曲序列,占整个序列的65.12%,是CEBPA蛋白的重要组成部分,有113个氨基酸构成了α-螺旋占全部结构的34.88%,与在线预测的蛋白三级结构的结果基本相似(图2-C,2-D)。同时预测得知鸡CEBPA蛋白主要在细胞核中占60.9%,一部分还分布在细胞质中占21.7%。运用STRTNG显示该蛋白与Runx转录因子家族、TRBs家族、PPARs核受体超级家族等有相互作用的关系,结果见图2-E。
2.2.2 CEBPA氨基酸序列的同源性分析 为了研究该基因的生物进化进程,对不同物种CEBPA的氨基酸序列进行同源性分析。结果表明,鸡与鹌鹑、人、猪、牛、兔、小鼠、斑马鱼的同源性分别为99.4%、93.9%、83.5%、85.3%、82.7%、83.3%、74.8%(图3-A)。并用这8个物种的CEBPA氨基酸序列构建系统进化树(图3-B),发现鸡与鹌鹑的亲缘关系最近,鸡与人和小鼠的亲缘关系最远。
2.2.3 CEBPA启动子和CpG岛的预测 在NCBI数据库获得鸡CEBPA 5'调控区-2 000 bp序列,在染色体上的位点为:Chromosome 11:10293396-10295395。并将这-2 000 bp作为候选启动子区用3种启动子预测软件进行预测分析,BDGP:Neural Network Promoter Prediction预测鸡CEBPA有7个启动子,Promoter 2.0预测鸡CEBPA有2个启动子,FPROM预测鸡CEBPA有4个启动子(图4-A)。使用EMBOSS和MethPrimer对鸡CEBPA 5'调控区-2 000 bp序列进行CpG岛预测,结果(图4-BC)两个预测软件结果完全一样,CpG岛位于CEBPA 5'调控区-2 000 bp序列的-265 bp - -1 895 bp之间,大小为1 640 bp。

图 1 鸡 CEBPA CDS 区克隆结果Fig.1 Cloning results of chicken CEBPA CDS region
2.3 鸡CEBPA在固始鸡时空表达谱的构建

本试验利用qRT-PCR方法构建12周龄固始鸡CEBPA的组织表达谱(图5-A)。结果表明,CEBPA胸肌和腿肌的表达量显著低于腹脂、皮下脂肪、肝脏的表达量(P<0.05)。利用qRT-PCR方法测量的6周龄、12周龄、22周龄、30周龄、55周龄固始鸡胸肌中CEBPA的表达量,构建时序表达图谱(图5-B)。结果表明,CEBPA在6周龄到55周龄的相对表达量呈U型,6周龄到22周龄CEBPA的相对表达量逐渐降低,22周龄到55周龄CEBPA的相对表达量逐渐增高。在22周龄时,相对表达量最低,且显著低于6周龄和55周龄(P<0.05)。
2.4 CEBPA在固始鸡和罗斯肉鸡胸肌中表达谱的构建
通过对6周龄的罗斯鸡(LS)和固始鸡(GS)胸肌中CEBPA的表达量的测定,发现罗斯肉鸡中的相对表达量显著高于固始鸡(P<0.05),大约为2.5倍,结果见图6。

图 2 CEBPA 蛋白结构和功能的预测Fig.2 Prediction of CEBPA protein structure and function
3 讨论
传统的生物学认为,蛋白质的序列决定了它的功能[15]。因此我们预测鸡CEBPA蛋白的结构和功能,表明该蛋白不存在跨膜域、无信号肽,不是分泌蛋白。结构域是蛋白质中具有特异空间结构和独立功能的区域,能够决定蛋白质发挥关键的生物学效用[16]。秦川牛脂肪沉积基因CEBPA的两种蛋白亚型都定位在细胞核中[14],与鸡CEBPA蛋白预测的结果不同,鸡CEBPA蛋白有21.7%存在于细胞质中,这为后续研究CEBPA在脂肪沉积中功能提供了理论基础。
PPI网络图中显示CEBPA蛋白多个家族存在互作作用,PPARG是一种依赖于配体激活的转录因子。PPARG、PPRα和PPARβ一起组成核受体超级家族,PPARs主要调节糖代谢和脂肪分布[17];HNF4A是肝细胞核因子(heptocytenuclear、HNF)家族的一员。在肝脏中表达量较高的一类转录因子,调节肝脏基因的特异性表达。HNF4A主要调控葡萄糖、氨基酸和脂质的代谢[18-19];TRBs家族有TRIB1、TRIB2和TRIB3,拥有类似激酶结构,却缺乏激酶的催化活性。其主要通过与靶蛋白结合,调节靶蛋白的功能的促进或抑制或蛋白稳定性[20]。TRIB2在人类急性髓系白血病表达增高,可抑制CEBPA的功能进而促进白血病的进展[21]。

图 3 CEBPA 氨基酸序列保守性分析Fig.3 Conserved amino acid sequence analysis of CEBPA

图 4 CEBPA 启动子和CpG 岛预测结构示意图Fig.4 Schematic diagram of the predicted structure of CEBPA promoter and CpG island
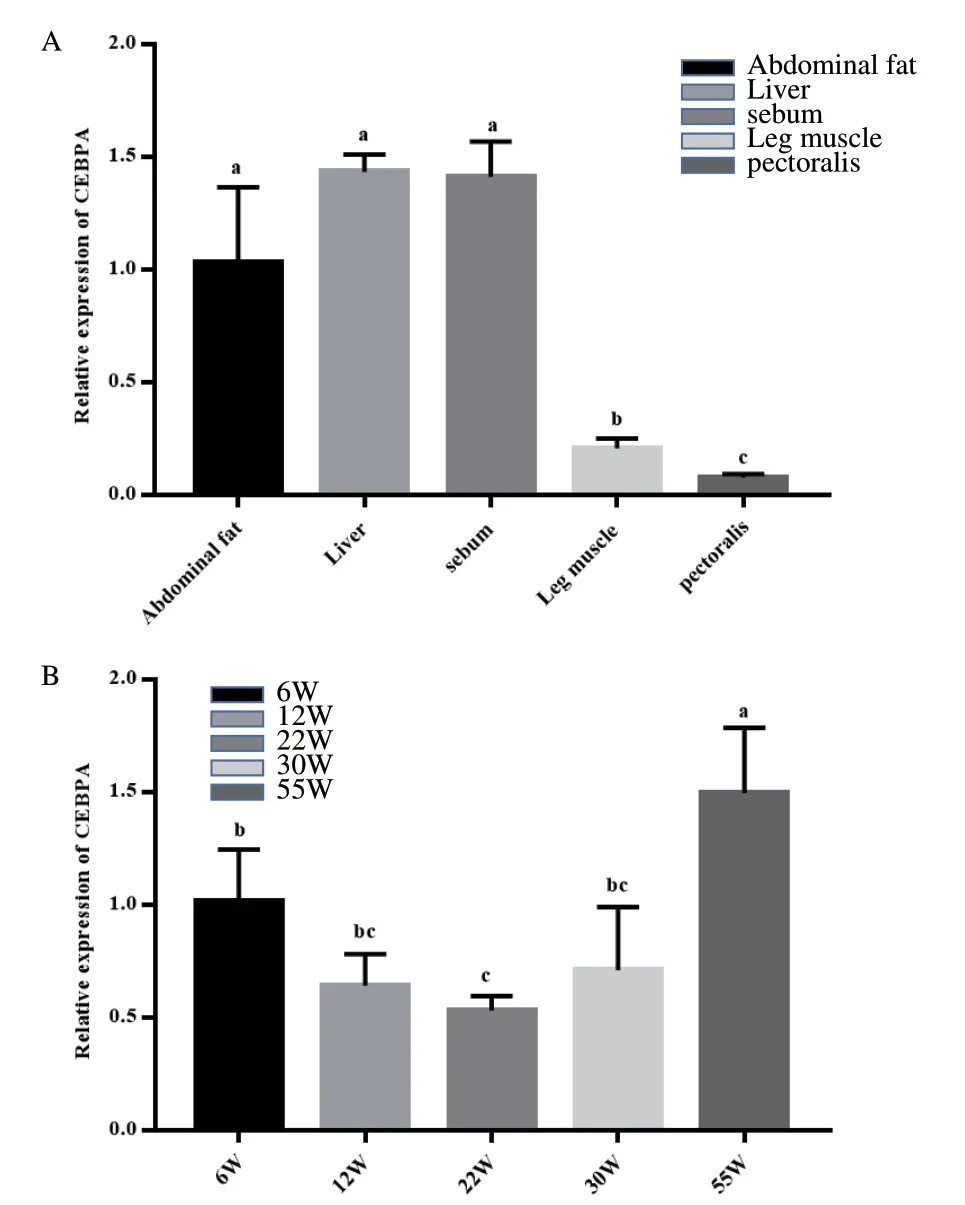
图 5 CEBPA 在固始鸡中的时空表达谱Fig.5 Spatial and temporal expression profiles of CEBPA in Gushi chickens

图 6 CEBPA 在 6 周龄固始鸡和罗斯鸡胸肌中的相对表达量比较Fig. 6 Comparison of the relative expression of CEBPA in the pectoral muscle of 6-week-old Gushi and Rose chickens
C/EBP通过结合抑制干细胞更新的基因以及激活髓样分化基因的启动子来诱导骨髓祖细胞的粒细胞分化和维持成人造血干细胞(HSC)静止[22-24]。人类已知的大多数CEBPA启动子在肝细胞或脂肪细胞中表达调控[25],因此我们利用在线软件预测鸡CEBPA基因5'调控区-2 000 bp序列的启动子,NNPP预测得到7个启动子,其中有两个启动子的分值在0.9以上。FPROM预测得到的4个启动子中只有一个有TATA盒。而Promoter2.0预测得到的2个启动子可信度都是临界性预测,且分值都在0.9以下,假阳性的可能性较大,为后来研究CEBPA的激活对脂肪沉积提供基础的信息;CpG岛主要位于基因的启动子和第一外显子区域,约有60%以上基因的启动子含有CpG岛。而人类的基因组中,约80%的CpG二核苷酸是甲基化的,常位于基因启动子区域的CpG二核苷酸是未甲基化的[26]。DNA的甲基化具有稳定性,可阻止转录因子的结合,从而调控基因的低表达或不表达[27]。在乳腺癌发病过程中,CEBPA的甲基化通过阻断CEBPA与精氨酸甲基化转移酶1(PRMT1)抑制因子HDAC3结合,促进细胞周期蛋白D1的表达,导致肿瘤细胞快速生长[28]。抑制组蛋白赖氨酸去甲基化酶7A(KDM7A)使CEBPA和分泌型卷曲相关蛋白1(Sfrp1)的表达下降,从而抑制了细胞的成脂分化,促进了成骨分化[29]。因此预测CEBPA的CpG岛,为后续研究提供了基础的资料。
基因在同一种生物不同器官组织中存在表达广泛性和特异性[30],基因在组织中的表达受多种因素调控,不同组织的功能不同,导致同一基因在不同组织中的表达也有所不同[31]。前人实验证实CEBPA是维持造血系统粒系分化过程中的重要的转录因子,在调节细胞增殖与分化的平衡中也起着关键作 用[32-34]。CEBPA是第一个在脂肪细胞分化中发挥重要作用的转录因子。它在大多数特异性脂肪细胞基因转录之前就已经得以表达,此外许多特异性脂肪细胞基因的最近端启动子上都含有CEBPA结合点。Wang等[13]在小鼠体内干扰了CEBPA,小鼠表现出白色脂肪组织发育停滞现象。本试验通过构建固始鸡不同组织表达谱可以发现,皮下脂肪、腹脂、肝脏中CEBPA的相对表达量较高,且差异不显著。因肝脏是产生脂肪的主要组织,由此可见肝脏中CEBPA相对表达量较高可能与肝脏的功能有关。同时腹脂与皮脂都是产生脂肪的组织,因此推测CEBPA对家禽体内脂肪的形成有重要作用。
本研究测定6周龄的罗斯鸡与固始鸡胸肌中CEBPA的表达量,发现罗斯鸡的表达量远远高于固始鸡中的表达量。经查资料可知,罗斯鸡为肉鸡,体内脂肪含量多,且生长快,饲料报酬高[35]。而固始鸡为蛋鸡,胸肌内的脂肪含量少。CEBPA与脂肪的合成有关,因此在罗斯鸡的表达量高与脂肪含量含量多有关。同时本研究还测得了固始鸡胸肌不同发育时期CEBPA的表达量,结果呈U型。在22周龄时表达最低,55周龄表达量最高。另课题组测得的6周龄、12周龄、22周龄、30周龄固始鸡胸肌的脂肪含量与CEBPA表达的趋势一致[36],还测得55周龄与20周龄相比胸肌的脂肪含量明显增加,也与CEBPA表达趋势相同。22周龄时固始鸡进入产蛋期,55周龄时已经经历了产蛋高峰期,进入衰老期,将要被淘汰[37]。产蛋期的鸡体内激素分泌旺盛,代谢能力增强[38],因此在产蛋期鸡体内的脂肪含量会有所变化。但随着鸡的年龄日渐增大,产蛋高峰期也逐渐过去[39],鸡体内的脂肪沉积会越来越多,所以推测此时期CEBPA基因对胸肌中脂肪沉积发挥着较大功能。
4 结论
本研究克隆了鸡CEBPA CDS区的全长序列(Genebank登录号:NC_006098.5),全长为975 bp,一个外显子。生物信息学分析表明,该蛋白在物种间的保守性高,不是分泌蛋白,定位于细胞核与细胞质中;该蛋白还与Runx转录因子家族、TRBs家族、PPARs核受体超级家族等多个家族相互作用;3种在线软件预测CEBPA 5'调控区-2 000 bp序列上共有13个启动子;预测该区域共有一个CpG岛且位于CEBPA 5'调控区-2 000 bp序列的-265 bp - -1 895 bp之间,大小为1 640 bp;qRT-PCR结果表明,CEBPA在胸肌和腿肌的表达量显著低于腹脂、皮下脂肪、肝脏的表达量;固始鸡胸肌中CEBPA的表达量在6周龄、12周龄、22周龄、30周龄、55周龄成U型结构,22周龄的表达量相对于6周龄和55周龄表达量显著降低;6周龄固始胸肌的表达量显著低于罗斯肉鸡的表达量。CEBPA可能参与调控胸肌的脂质沉积。
猜你喜欢
理财周刊(2022年4期)2022-04-30
昆明医科大学学报(2022年1期)2022-02-28
特产研究(2021年6期)2021-12-08
现代装饰(2021年5期)2021-12-02
畜禽业(2021年11期)2021-11-22
数学小灵通·3-4年级(2020年5期)2020-06-24
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
学生天地·小学低年级版(2019年7期)2019-10-08
河南农业科学(2019年9期)2019-09-24
