
基于SSR标记的贵州水稻纹枯病菌遗传多样性
2021-09-13 07:23谢海呈谭清群杨学辉
贵州农业科学 2021年7期
谢海呈, 谭清群, 徐 娅, 刘 霞, 杨学辉*
(1.贵州省农业科学院 水稻研究所,贵州 贵阳 550009;2.贵州省农业科学院 植物保护研究所,贵州 贵阳 550009;3.湄潭县植保植检站,贵州 湄潭 564100)
0 引言
【研究意义】由立枯丝核菌(Rhizoctoniasolani)侵染引起的水稻纹枯病是水稻产区的常发性病害,以往发生对产量的影响不大,近年来随着高产、矮秆、多蘖品种以及高肥密植栽培技术的推广应用,该病的危害程度日益加重,危害最高损失可达50%[1-2]。在我国部分地区,其危害已超过稻瘟病而居首位[3]。该病为土传病害,病菌长期存于土壤中,寄主范围广,重要寄主作物有水稻、玉米、大麦、高粱、豆类、花生等,其在田间的发病程度与初始菌量相关[4]。开展病原菌遗传多样性研究有助于了解病害在田间的流行趋势和防控策略的制定。【前人研究进展】目前,有关水稻纹枯病菌遗传多样性研究多采用分子标记技术,易润华等[5]采用RAPD标记广东省病菌群体遗传多样性在不同水平上的分布;李挺丹等[6]基于ISSR明确了福建不同水稻纹枯病菌群体间存在明显的遗传分化且与菌株的地理来源相关;王爱军等[7-8]采用rDNA-ITS序列对水稻纹枯病菌进行了融合群鉴定、融合群组成与分布等研究;王玲等[2]采用SSR分析了广东等8个省水稻纹枯病菌的遗传多样性的形成与遗传分化;朱明海等[9]采用AFLP解析了海南南繁区核心区与非核心区的遗传变异。水稻纹枯病是影响贵州水稻生产的主要病害之一,近2年田间病株率达5%~86%,局部重发或部分田块全发病。【研究切入点】目前,对该病的研究主要集中于品种选育和病害防治,对病原菌群体遗传结构特征研究较少。【拟解决的关键问题】采用SSR标记对贵州省不同地区的水稻纹枯病菌进行遗传多样性分析,旨在揭示病菌群体遗传结构的组成和群体间的遗传分化,为水稻纹枯病防控措施的制定提供依据。
1 材料与方法
1.1 供试菌株
2018—2019年从贵州省9个地区(州、市)26个县(区)采集的水稻纹枯病标样277个,其中黔南州38株,黔东南州27株,黔西南州36株,铜仁地区41株,毕节地区28株,贵阳市15株,安顺市24株,遵义市30株,六盘水市38株。
1.2 基因组DNA提取
将经过活化的水稻纹枯病菌的菌丝体接种于装有50 mL马铃薯液体培养基(马铃薯200 g,葡萄糖20 g,蒸馏水1 000 mL)的三角瓶中,28℃恒温下180~200 r/min振荡培养3 d,收集菌丝提取菌株DNA。采用索莱宝的真菌基因组DNA提取试剂盒(Cat#D2300),提取步骤按说明书进行。
1.3 SSR-PCR扩增
利用11对SSR荧光标记引物进行PCR扩增。SSR-PCR反应体系为20 μL,含有dNTP 0.4 μL,Buffer 2 μL,前后引物各0.3 μL(20 μmol/L),DNA模板2 μL,Taq 0.2 μL及ddH2O 14.8 μL。扩增程序:94℃预变性5 min;94℃变性30 s,65℃到50℃降落复性30 s,72℃延伸40 s,共35个循环;最终72℃延伸3 min。PCR产物经琼脂糖电泳抽检合格后进行毛细管电泳检测。
1.4 毛细管电泳检测
将甲酰胺与分子量内标按体积比100∶1的混匀后,取15 μL加入上样板中,再加入1 μL稀释10倍的PCR产物,然后使用ABI 3730XL测序仪进行毛细管电泳。利用Genemarker V2.2.0中的Fragment(Plant)片段分析测序仪得到的原始数据,将各泳道内分子量内标的位置与各样品峰值的位置比较分析后得到片段大小。
1.5 数据处理
每对引物检测1个等位基因位点,将每个样品在各等位基因位点的不同片段用Convert1.31转化成POPGENE能识别的格式。使用POPGENE32计算等位基因数(Na)、有效等位基因数(Ne)、Shannon’s信息指数(I)、Nei’基因多样性指数(He)、遗传分化系数(Fst)及基因流(Nm),NTSYS对Nei’s遗传距离进行UPGMA聚类分析,构建系统发育树。GenAlEx进行分子方差分析(AMOVA),明确各群体每个位点的遗传分化。
2 结果与分析
2.1 贵州水稻纹枯病菌群体的SSR位点多样性
11个SSR位点均表现出多态性,多态性位点百分率(P)为100%。全部位点在277个水稻纹枯病菌菌株中共检测到92个等位基因,分子量变异范围为124~283 bp,图1所示为毛细管电泳检测的引物RH2对2个菌株的扩增片段,在225 bp处均有1个峰值。由表1可见,每个位点的等位基因数为4~14个,平均8.363 6个;平均有效等位基因数为2.277 7个,变幅为1.115 0~3.552 9;平均Shannon’s信息指数为1.066 4。

图1 水稻纹枯病菌在RH2位点上的毛细管电泳峰型图
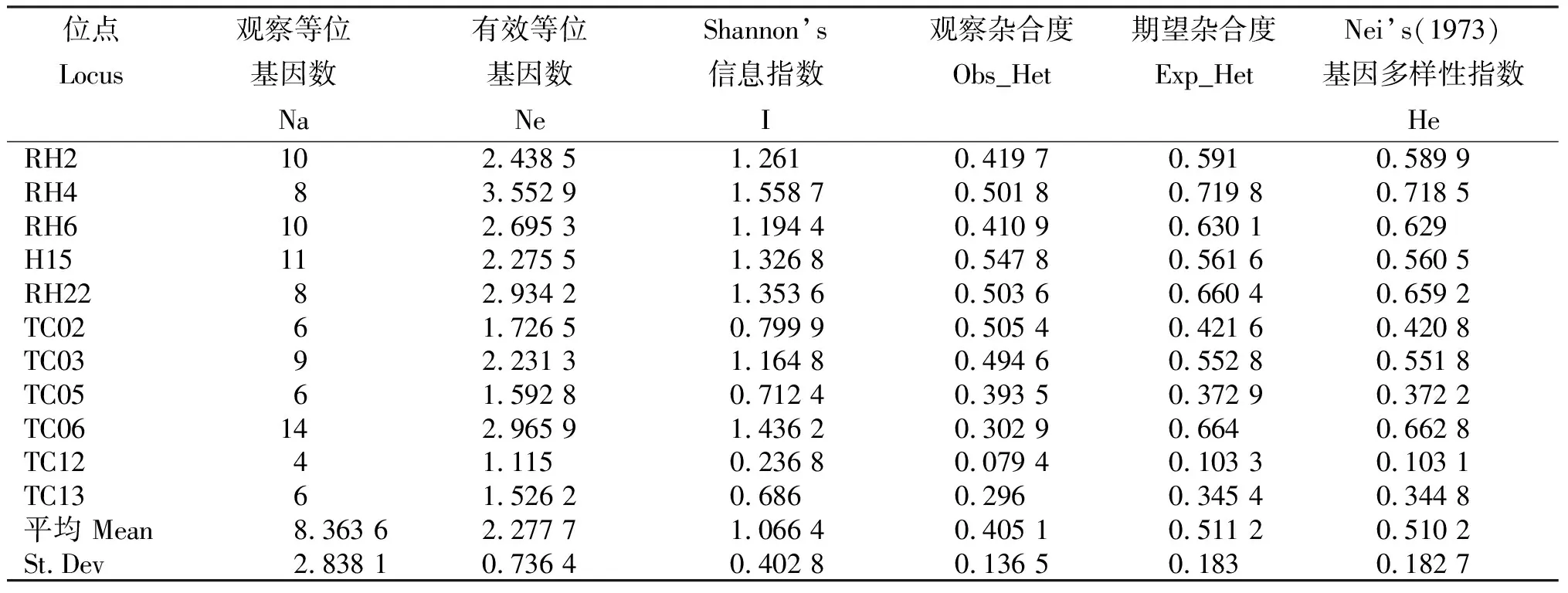
表1 水稻纹枯病菌SSR位点多样性参数
2.2 不同地区水稻纹枯病菌群体的遗传多样性
从表2可知,对于不同地区来源的水稻纹枯病菌群体,其遗传多样性有一定差异,9个地区的水稻纹枯病菌群体中,观测的等位基因数在3.090 9~5.636 4,铜仁群体最高,贵阳群体最低;有效等位基因数为1.864 3~2.611 4,最高和最低群体分别是铜仁和安顺;Shannon’s信息指数最高为铜仁群体(1.102 6),最低是安顺群体(0.762 4);而Nei’基因多样性指数的变幅为0.545 9(铜仁)~0.398 8(毕节)。综合以上4个参数分析,铜仁地区水稻纹枯病菌群体的遗传多样性最为丰富,其次是黔西南州、黔东南州和黔南州,贵阳、安顺和毕节群体的相对较低。
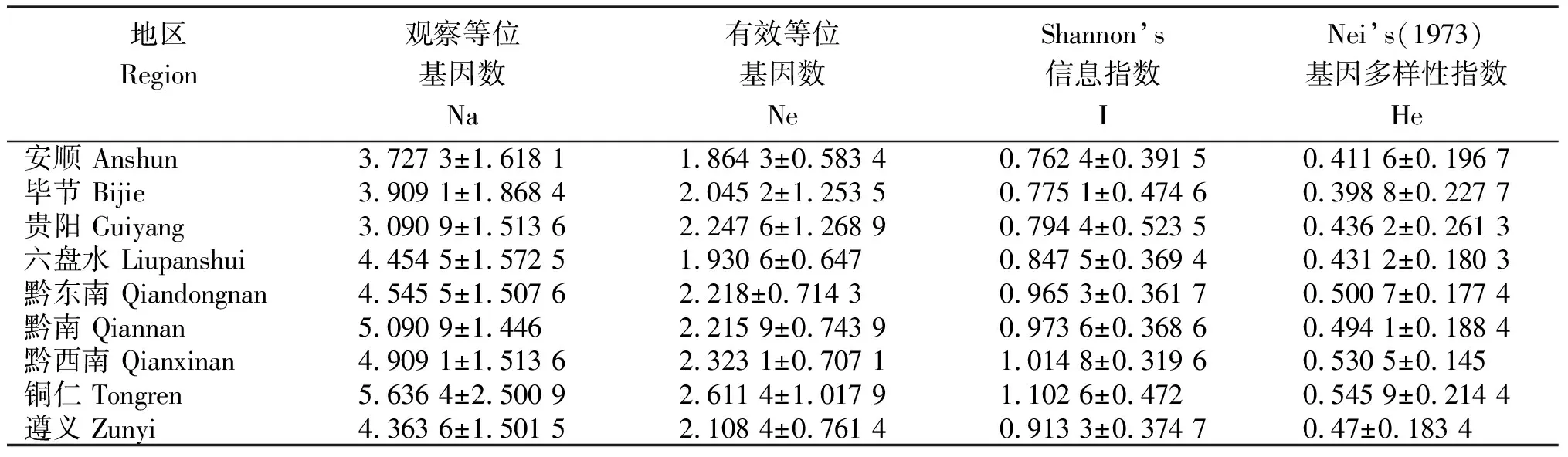
表2 贵州9个地区水稻纹枯病菌群体的遗传多样性参数
2.3 不同地区水稻纹枯病菌群体的分子方差
贵州9个地区纹枯病菌群体的分子方差分析显示(表3),不同地区群体间的变异占9%,而地区内菌株间遗传变异为91%,说明省内水稻纹枯病菌的遗传变异主要是由不同群体内菌株间差异引起。比较不同地区来源水稻纹枯病菌群体的遗传分化值(表4),群体间已有遗传分化,其中,毕节-安顺纹枯病菌群体间的遗传分化值最大,为0.176,其次是贵阳-黔东南(0.174)、毕节-黔东南(0.171)和安顺-黔南(0.170),较低的群体是六盘水-遵义(0.028)和六盘水-毕节(0.050),各群体间的遗传分化均达显著差异(P<0.05)。

表3 不同地区水稻纹枯病菌群体的分子方差分析

表4 贵州9个地区水稻纹枯病菌群体的遗传分化值及概率
2.4 不同地区水稻纹枯病菌的聚类树
由图2可见,从遗传距离和遗传相似度分析,贵州9个地区的菌株群体间遗传上均较接近,遗传相似度的变幅为0.859 5~0.973 2。两两群体间遗传相似度最大(GI=0.973 2)、遗传距离最小(GD=0.027 1)的群体是遵义与六盘水的菌株群体,而遗传相似度最小(GI=0.859 5)、遗传距离最大(GD=0.154 1)的是贵阳与黔东南的菌株群体。在遗传相似系数0.91处,9个水稻纹枯病菌群体可以分为明显的2个类群,安顺、黔东南、黔西南聚为一个类群,其余6个地区的菌株群体聚另一类群。在第二类群中,黔中的贵阳、黔北的遵义和黔西北的毕节与六盘水聚为亚群一,铜仁和黔南聚为亚群二。

图2 9个地区水稻纹枯病菌群体聚类树
对于全部菌株群体(图3),在0.58处可将277个菌株划分为9类群,其中,类群1、2包括了大部分菌株(155株、107株),其余7个类群均为仅含个别菌株的小类群,而两个大类群又各自聚为不同的亚群。第1类群分为2个亚群,各有19个、136个菌株;第2类群的2个亚群各有79个和28个菌株。经分析菌株其遗传相似性与地理来源无直接关系。总体来看,以0.37和0.53的遗传相似系数,全部群体均是划分为占绝对优势的大类群和个别菌株的小类群。可见,贵州省水稻纹枯病菌存在较为丰富的遗传多样性,既有明显的主体类群,又有多样性各异的小类群。

图3 277株水稻纹枯病菌SSR分子指纹UPGMA聚类树
3 结论
贵州近几年水稻纹枯病的危害趋于严重,但对其病原菌的遗传多样性研究至今未有报道,加强基于分子标记技术的水稻纹枯病菌群体遗传结构特征和变异特点研究,有利于了解病害的发生流行进程并对病害的防治具有重要的指导作用。研究结果表明,源于贵州9个地(州、市)的277个水稻纹枯病菌菌株在11个SSR位点上全部为多态位点,共检出92个等位基因,平均每个位点8.36个,平均期望杂合度(He)和Shannon’s信息指数(I)分别为0.511 2、1.064 6,与王玲等[2]关于中国南方八省水稻纹枯病菌群体的同类研究(He=0.472,I=0.859)相比,贵州的病菌群体具有较高的遗传多样性。
在不同地理来源上,贵州省不同地区来源的水稻纹枯病菌群体遗传多样性存在一定差异,黔东南、黔西南、黔南、铜仁等少数民族地区纹枯病菌群体的遗传多样性明显高于贵阳、安顺和遵义等地区(市),这是否与少数民族地区至今拥有较为丰富的品种类型有关有待进一步研究。从分子方差分析结果来看,不同地区的病菌群体存在遗传分化,但其变异的91%是由不同群体内菌株间差异引起,说明了贵州水稻纹枯病菌的遗传分化与其地理来源关系不明显,而与病害发生地的寄主和环境生态有着较大的关系。
4 结论
采用SSR荧光标记方法对贵州水稻纹枯病菌遗传多样性的分析表明,贵州省内水稻纹枯病菌群体具有较为丰富的遗传多样性,检测的11个位点均为多态性位点,共检测到92个等位基因,平均等位基因数、有效等位基因数、香农指数和Nei’s基因多样性指数分别为8.363 6、2.277 7、1.064 6和0.510 2。铜仁地区水稻纹枯病菌群体的遗传多样性稍高于其他地区,不同地区间病菌群体存在一定的遗传分化,但其变异主要由群体内菌株间差异引起,基于Nei’s遗传相似系数将贵州省水稻纹枯病菌划分为2个主体类群和7个小类群,不同群体的遗传分化及菌株间的亲缘关系与地理来源不明显。
猜你喜欢
科学与财富(2021年33期)2021-05-10
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
农民致富之友(2019年21期)2019-08-08
农民致富之友(2019年4期)2019-03-13
新课程·下旬(2018年9期)2018-11-14
中国医药导报(2018年14期)2018-08-30
江苏农业科学(2017年17期)2017-11-15
青少年科技博览(中学版)(2015年10期)2015-01-11
小说评论(2014年3期)2014-11-14
