
小鼠原发性胆汁性胆管炎模型及其在疾病研究中的作用
2021-09-08 07:22关艳玲
中国药理学通报 2021年9期
李 浩,关艳玲,张 雨,刘 倩,魏 伟,马 旸
(安徽医科大学临床药理研究所,抗炎免疫药物教育部重点实验室,安徽抗炎免疫药物协同创新中心,安徽 合肥 230032)
原发性胆汁性胆管炎(primary biliary cholangitis,PBC)是一种慢性胆汁淤积性肝病,其特征是肝内小胆管的破坏,疾病最终可进展为肝纤维化和潜在的肝硬化。目前,PBC发病原因尚未明确,可能由环境、细菌感染、基因等诸多因素引起。人群中年发病率为 0.9-5.8人/10万人,92%的患者为女性,目前除部分晚期患者接受肝脏移植外,针对PBC中早期的治疗手段有限。目前FDA仅批准了两种治疗药物——熊去氧胆酸(ursodeoxycholic acid,UDCA)和奥贝胆酸(obeticholic acid,OCA),但约有40%的患者对UDCA无应答[1]。因此,研究PBC的发病机制、寻找药物的作用靶点对于开发治疗PBC的药物十分重要。动物模型是阐述人类疾病病因和病理机制、开发治疗药物的重要工具,对于PBC的发病机制的探索以及药物的研发有着重要意义。本文综述了近年来国内外报道的PBC动物模型研究进展以及各种模型在疾病研究中的应用,为PBC模型的选择提供参考。
1 PBC的发病机制
PBC的发病机制十分复杂,受遗传易感性、环境因素及免疫耐受异常的影响。在遗传易感性方面,人类白细胞抗原和白介素-12在PBC的发生中起重要作用;环境因素可通过分子模拟打破机体对线粒体抗原的自身耐受, 启动自身免疫反应;机体的免疫耐受缺失(如T细胞、B细胞的免疫调控失衡),可导致胆管上皮细胞(biliary epithelia cell,BEC)损伤。
自身抗体在PBC的发病中可能起着关键的作用,也是PBC的临床诊断的重要标准之一。在PBC的发生过程中,巨噬细胞吞噬抗原,提呈给T细胞,T细胞分化成效应T细胞并产生一系列的细胞因子,同时B细胞在效应T细胞的辅助下,增殖分化为浆母细胞,最终分化为浆细胞,产生一系列的抗体,其中包括特异性自身抗体——抗线粒体抗体(anti-mitochondrial antibody,AMA)。AMA是PBC的特征性自身抗体,见于95%以上的PBC患者, 且随着检测技术的提高, AMA的阳性检出率也明显升高,但目前认为其水平高低与疾病严重程度关系不大。此外,1/3以上的PBC患者可检出抗核抗体(antinuclear antibody,ANA), 其中抗gp210、p62、sp100、PML等抗核抗体对诊断AMA阴性的PBC患者具有较高的诊断价值,可能与疾病的预后有关[2]。
T细胞在PBC的发病过程中起着关键的作用,自身反应性CD4+T细胞和CD8+T细胞被认为是参与PBC的主要机制之一,此前的研究发现PBC患者受损胆管周围存在大量致病性CD4+T细胞和CD8+T细胞,并且证实CD4+T细胞和CD8+T细胞是主要的致病因素。其中,PBC患者肝组织炎症部位中浸润的淋巴细胞主要是CD8+T细胞,CD8+T细胞是患者胆管破坏的效应细胞[3],主要通过表达FasL及分泌穿孔素和颗粒酶B引起BEC的凋亡。辅助性T细胞(T helper cell, Th)的活化是自身免疫病的重要致病机理之一,有报道称,Th 17/调节性T细胞(regulatory T cell,Treg)比例失衡,在类风湿关节炎等自身免疫病中起着关键作用[4],而PBC患者中Treg数量下降, Th17细胞数量增高, 打破两者之间的动态平衡, 维持自身免疫应答的稳态遭到破坏,可能是引发PBC病症的原因。
PBC患者的血清B淋巴细胞活化因子水平相比健康人群及丙型肝炎患者均显著升高, 并且PBC的肝脏组织亦可见B细胞浸润。提示B淋巴细胞可能在PBC的发病过程中起重要作用,但其具体的作用机制仍有待进一步研究。研究表明[5],采用利妥昔单抗(rituximab)耗竭B细胞,可以显著减少B细胞所产生的AMA,降低血清AMA滴度,同时降低血浆免疫球蛋白(IgA、IgM、IgG)的水平以及碱性磷酸酶(alkaline phosphatase,ALP)的水平,提示耗竭B细胞可能成为治疗PBC的新靶点。
自然杀伤细胞(nature killer cell, NK)和自然杀伤性T细胞(nature killer T cell,NKT)与PBC的发病过程紧密相关。研究发现,肝脏的NK细胞具有不同的表型,在dnTGFβRⅡ(dominant-negative transforming growth factor-β receptor type Ⅱ)小鼠模型中,肝脏的DX5—NK细胞作为CD4+T细胞的负性调节因子,可以通过抑制肝脏的CD4+T细胞反应来抑制肝脏炎症,但是整体的NK细胞对于适应性免疫的调节是双向的[6]。NKT细胞是打破免疫耐受的始动者, 分泌多种细胞因子, 激活CD4+T细胞、CD8+T细胞等多种细胞, 诱发PBC。
单核细胞也是PBC患者胆管周围常见的浸润细胞,促炎表型的单核细胞在PBC患者的外周血中显著增多,在PBC中其促炎功能显著升高,并且与肝脏的损伤程度有关[7],此外,PBC患者外周血中的单核细胞对外界的刺激更加敏感,在病原相关分子模式刺激下,经介导其分泌的促炎因子增多。
BEC是PBC中免疫破坏的靶细胞,在PBC的发病机制中起着关键的作用。在相应刺激下能分泌炎性因子以及趋化因子,可能和PBC的发病过程相关。此外,BEC表达HLA-I、HLA-II和多种共刺激分子,促进T细胞的免疫应答。有报道称,多种原因造成的BEC自噬通量下降,会使得肝内中、小胆管反复受到免疫系统攻击,不能通过自噬维持稳态,最终导致肝纤维化和肝硬化[8],提示,维持细胞的自噬功能,增加其自噬通量,有可能成为治疗PBC的新方向。
PBC主要由自身免疫驱动,胆汁酸和肠道菌群同样参与了疾病的进展。胆汁酸能调节胆管反应和胆汁淤积性损伤,并且胆汁酸会使得肝脏的炎症持续,并且随着时间的推移导致纤维化。研究表明,对胆汁酸受体使用激动剂、拮抗剂或补充特定的胆汁酸可以调节胆管细胞的增殖和功能[9]。肠道菌群和细菌移位被认为是粘膜免疫反应和耐受的重要原因,肠道微生物群的移位也被认为可以促进PBC[10]。
总之,PBC的发病受到多种因素的影响,其中,自身免疫耐受失衡和胆汁酸循环障碍是导致PBC发生的直接因素。胆汁酸循环障碍可能会导致胆汁淤积症,而胆汁淤积将会对BEC产生毒性作用,损伤BEC;而自身免疫耐受失衡可能会导致免疫细胞持久性的攻击BEC,导致BEC的进一步损伤以及疾病的进一步恶化。
2 PBC小鼠模型
在过去的几十年中,一系列的PBC模型被相继报道。这些模型加快了进一步研究PBC的发病机制的速度,极大地促进了PBC治疗药物的研发。本综述介绍多种PBC小鼠模型,描述了该模型的形成原因、组织病理学变化及利用PBC小鼠模型对疾病发病机制和药物开发的研究进展。
2.1 手术模型手术制备PBC小鼠模型的方法主要是胆管结扎模型(bile duct ligation, BDL),通过结扎小鼠的胆总管,造成胆总管阻塞,胆汁淤积,从而引起一系列的炎症反应,引发肝纤维化和肝硬化。该方法于实验小鼠腹腔切口,在十二指肠起始端胆总管处进行胆总管结扎。此前的研究表明,对C57BL/6J雌性小鼠胆总管结扎,可导致小鼠肝脏发生炎症反应,汇管区可见嗜中性粒细胞、肥大细胞以及淋巴细胞浸润[11]。此外,也有采用大鼠进行胆管结扎并成功引起手术大鼠的胆管阻塞,胆汁淤积,总胆汁酸(total bile acid, TBA)和谷丙转氨酶(alanine aminotransferase,ALT)、谷草转氨酶(aspartate aminotransferase,AST)水平升高的报道[12]。尽管该模型手术方法成熟、成模率高、实验成本低、可造成胆总管阻塞、肝内胆汁淤积,但并未涉及引起PBC相关的特异性抗体AMA等因子升高,因此,该模型多用于胆汁淤积性疾病的研究。
2.2 药物诱导模型
2.2.12-OA-BSA诱导模型 2-辛炔酸偶联牛血清白蛋白(2-octynoic acid coupled to bovine serum albumin,2-OA-BSA)诱导的小鼠模型,可以很好地模拟人类PBC的疾病特征。在完全弗氏佐剂的存在下,用包含结合分歧杆菌株H37Ra(10 g·L-1)的2-OA-BSA(100 g/25 L)腹膜内接种8-9周龄的C57BL/6J雌性小鼠,随后每隔2周在不完全弗氏佐剂的存在下,对小鼠进行1次腹膜内接种以加强诱导模型。
2-OA-BSA小鼠模型具有许多和人类PBC类似的血清学和病理学特征。用2-OA-BSA诱导小鼠,第4周时,小鼠血清中的抗丙酮酸脱氢酶复合体E2亚基(E2 subunits of pyruvate dehydrogenase,PDC-E2)IgG和IgM抗体显著升高,第8周时血清抗PDC-E2的IgG和IgM、IgA抗体均升高。在2OA-BSA免疫4-12周时,血清肿瘤坏死因子α(tumor necrosis factor α,TNFα)以及干扰素γ(interferon γ,IFN-γ)的水平升高,免疫12周后,小鼠的肝脏CD8+T淋巴细胞的比例增高,CD4+T淋巴细胞的比例降低,肝脏CD4+/CD8+T淋巴细胞的比例倒置,而脾脏中CD8+、CD4+T淋巴细胞比例均降低。汇管区可见大量的淋巴细胞或单核细胞浸润于受损的胆管周围,主要是CD4+和CD8+T淋巴细胞,其中CD8+T淋巴细胞占主导地位。此外,还可以观察到肝实质和汇管区的脂肪减少,上皮样肉芽肿的形成,肝实质也可见小面积的轻度局灶性坏死,这些都与人类的PBC相似。
该模型小鼠也有许多与不同于人类PBC的血清学和病理学的特征,在2-OA-BSA诱导第24周时,未见肝脏的脂肪变性、嗜酸性粒细胞增多以及胆汁淤积的现象。此外,在该模型小鼠的其他组织器官,包括甲状腺、唾液腺、肺、肾、小肠和结肠未产生免疫损伤,这与人类PBC患者的肝外器官也出现炎症反应的特点不相符[13]。
采用2-OA-BSA诱发小鼠发生类PBC疾病的方法,可能对人类PBC的病因学提供了新的证据,即环境因素在人类PBC发病中的重要性。
2.2.2DDC诱导模型 采用0.1%的1,4-二氢-2,4,6-三甲基-3,5-吡啶二甲酸二乙酯(3,5-diethoxycarbonyl-1,4-dihydrocollidine,DDC)对2月龄的雄性瑞士白化病小鼠饮食喂养8周,即可制备具有类似人类PBC特征的DDC小鼠模型。小鼠暴露于含有DDC的饮食中,可诱导卟啉积累、胆汁淤积、免疫反应和类似肝卟啉症和硬化性胆管炎的肝胆损伤,出现类似人类PBC的胆管损伤和肝纤维化,但与人类PBC不同的是该模型小鼠的胆管损伤与肝纤维化是平行发生的。此外,该模型小鼠出现一定的胆汁流量的减少,但是无统计学意义,即未出现类似人类PBC的胆汁淤积症状。此外,DDC诱导了TNF-α以及骨桥蛋白的表达,骨桥蛋白的表达也偶见于人类PBC[14]。
虽然,该模型小鼠表现出类似人类PBC的胆管和胆管周围的病理损伤,但该模型小鼠中未观察到AMA升高等人类PBC的特异性血清学指标,因此该模型小鼠一般用于观察外源性因素诱导的胆道疾病和胆汁性肝纤维化。
2.2.3polyI:C诱导模型 肌苷酸聚胞苷酸(polyinosinic polycytidylic acid,poly I:C),是一种1型干扰素(interferon, IFN)诱导剂,采用腹腔注射poly I:C(5 mg·kg-1)诱导6-8周龄雌性C57BL/6J小鼠,每周注射两次,共28周,即可制备具有类似人类PBC特征的poly I:C小鼠模型[15]。
在poly I:C注射后小鼠血清IFN-α水平升高,注射后3 h内达到最高,注射后24 h后逐渐下降至无法检测,除了IFN-α外,poly I:C还诱导了其他的促炎细胞因子,包括IL-12 p70、IL-10、单核细胞趋化蛋白-1以及IFN-γ,其中,IL-6的水平在24 h后仍然较高。Poly I:C诱导4周后,小鼠出现唾液腺炎,胰腺炎以及间质性肾炎。注射8周后,汇管区出现大量炎性细胞浸润,主要是CD4+以及CD8+T淋巴细胞,其中CD8+T淋巴细胞主要定位于胆管附近,肝脏组织中还检测到CD11b+巨噬细胞以及CD11c+树突细胞。此外,该模型小鼠血清ALT以及ALP的水平均升高,在polyI:C诱导第24周时,所有小鼠都表现出一种或两种自身抗体。
Poly I:C诱导模型虽然在AMA的阳性率以及肝纤维化等方面与人类PBC特征有一定差异,但该模型在研究IFN-α在PBC的病因学中具有重要作用。
2.2.42-OA-BSA联合polyI:C诱导模型 由于2-OA-BSA模型不具有人类PBC患者的肝脏嗜酸性粒细胞浸润增多以及肝纤维化等特点,而polyI:C模型存在AMA阳性率过低等问题,将2-OA-BSA与polyI:C联合应用于C57BL/6J小鼠,制得的联合模型更好的模拟了人类PBC。选用5-6周龄的雌性C57BL/6J小鼠,在2-OA-BSA首次注射免疫后的3 d注射polyI:C(5 mg·kg-1),每3 d注射1次,即可制得2-OA-BSA联合polyI:C小鼠模型。
该模型相较于自然病史和疾病严重程度较低的2-OA-BSA模型,表现出了更严重的胆管疾病,肝脏CD8+T细胞的浸润显著增加,IFN-γ、TNF-α、IL-12p40以及IL-6等多种炎性细胞因子的水平均增加。此外,该模型汇管区和肝实质嗜酸性粒细胞浸润增多,肝脏纤维化形成的特点更加符合人类PBC的病理特点。
2.3 抗原免疫模型
2.3.1新鞘氨醇杆菌免疫模型 新鞘氨醇杆菌(Novosphingobium aromaticivorans)是一种革兰阴性菌,通常存在于土壤、水和海岸平原沉积物中,其和PBC患者体内的线粒体自身抗原PDC-E2、侧链二氧酸脱氢酶复合体E2亚基(2-Oxo acid dehydrogenase,BCOADC-E2)以及2-氧戊二酸脱氢酶复合体E2亚基(2-Oxo-glutarate,OGDC-E2)具有高度的氨基酸同源性,并且与PBC患者体内的PDC-E2表达相同的鞘糖脂CD1,可被NKT细胞识别并激活该细胞,将新鞘氨醇杆菌以5×107cfu静脉注射C57BL/6J、NOD以及SLJ品系小鼠,两周后所有品系小鼠均出现胆管损伤、汇管区大量炎性细胞浸润,以及肉芽肿的形成等类似人类PBC的疾病表现。此外,该模型小鼠还表现出PDC-E2以及AMA的聚集、嗜酸性粒细胞的浸润,80%的小鼠检出AMA抗体阳性[16]。
虽然该小鼠模型制备周期短,操作便捷,但所有的小鼠均未检测到人类PBC中所见的严重肝纤维化,并且在其他的组织器官,包括甲状腺、肾脏、关节以及肠道中未发现慢性炎性,这也与人类PBC的疾病表现不相符。
2.3.2大肠杆菌免疫模型 除了新鞘氨醇杆菌外,大肠杆菌肽序列中有6-8个氨基酸残基与人类PBC的PDC-E2具有同源性。将大肠杆菌以5×107cfu通过眶周静脉丛注射入♀ 6周龄的NOD.B6Id10/Idd18小鼠体内,14 d后采用同样方法再次注射,即可制备大肠杆菌诱导模型。
大肠杆菌诱导模型相比于新鞘氨醇杆菌诱导模型,小鼠表现出更严重的胆道疾病,而且血清AMA滴度更高,大肠杆菌感染4周后,小鼠血清AMA滴度达到峰值,然后逐渐下降至与新鞘氨醇杆菌感染小鼠相同水平。大肠杆菌感染26周后,小鼠肝脏出现明显的汇管区炎症反应,并且伴有肉芽肿的形成,此外小鼠出现程度不一的胆管损伤,轻度表现为淋巴细胞的聚集,重度胆管上皮细胞几乎完全消失[17]。
虽然大肠杆菌诱导模型和新鞘氨醇杆菌诱导模型都未能使得小鼠表现出肝脏纤维化等人类PBC特征,且该类模型在制备时会造成小鼠合并发生尿路感染,降低模型制备的特异性,但是其为研究感染因素在PBC发病中的作用提供了新的思路。
2.3.3胆管蛋白免疫模型 BDP胆管蛋白(bile duct protein,BDP)模型,采用同系胆道抗原有效的破坏受体小鼠的免疫耐受。对野生型的8-12周龄C57BL/6J小鼠进行BDP分离,然后对7-8周龄的小鼠进行BDP免疫,即采用BDP乳液进行多点皮下注射,即可制得BDP免疫小鼠模型。
研究表明,同基因的胆道抗原能有效地破坏受体小鼠的免疫耐受,并且在BDP免疫小鼠中表现出PBC的几个关键特征,包括肝脏汇管区的特异性炎症浸润,增加肝脏和脾脏的CD4+和CD8+T细胞数目和激活状态。小鼠的AMA呈100%阳性,此外,在该模型中,肝脏和脾脏的Treg百分比都降低,这可能与T细胞的进一步激活有关[18]。
BDP模型突破了以往PBC模型的局限,以往并没有研究集中在突破耐受性的经典模型,即用自体组织进行免疫。因此,BDP模型强调胆管抗原在PBC发病机制中的作用。此外,BDP模型的建立主要依赖胆管抗原,而人体内的胆管抗原可能反映分子拟态介导的环境因素,这与2-OA-BSA诱导的模型以及大肠杆菌诱导的模型相似。BDP模型强调的是通过激活自身免疫淋巴细胞耐受性崩溃的理论,这与基因突变所诱发的肝脏疾病不同。BDP模型也有着一些缺点和不足,在该模型中并没有观察到有肉芽肿的形成和胆管损伤,此外,该模型是否存在性别偏倚,仍有待进一步研究。
2.3.4纯化的AMA-M2免疫模型 AMA是PBC的特异性抗体,9个亚型中以AMA-M2的特异性最强。而AMA-M2主要针对线粒体内膜上的PDC-E2抗原,这种抗原广泛存在于体内的有核细胞中,并且AMA仅作用于肝胆管线粒体抗原,但机制目前仍不清楚。
将纯化的PDC-E2、OGDC-E2、BCOADC-E2三联体抗原,无菌盐水稀释(1 g·L-1)后,乳化于等体积的完全弗氏佐剂,腹腔注射(200 μL/只)接种于C57BL/6J小鼠,即可制备AMA-M2诱导模型。免疫66周后,100%模型小鼠体内出现AMA-M2抗体、血清ALP水平显著升高,但模型小鼠肝脏并未出现淋巴细胞浸润以及纤维化,虽然引起部分胆管损伤,但在其他组织器官中,如肾脏、胃和肌肉中也未观察到淋巴细胞浸润[19]。
该模型制备周期长,与人类PBC相似的血清学和病理学特征有限,但该小鼠模型稳定性高,可用于研究特异性抗体AMA-M2在PBC的作用。
2.4 基因自发模型
2.4.1NOD.c3c4小鼠模型 非肥胖糖尿病(Non-obese diabetic,NOD)小鼠可以自发自身免疫性Ⅰ型糖尿病,NOD.c3c4小鼠模型是第一个被报道的PBC自发小鼠模型,是在识别糖尿病的基因片段中偶然发现的。NOD.c3c4小鼠是在NOD小鼠的背景下,在其3号和4号染色体(c3/c4)上分别插入B6/B10来源的胰岛素依赖性糖尿病(insulin dependent diabetes,Idd)的抗性等位基因。在B6和B10抗性等位基因与NOD基因组发生交互作用之后,导致了一种新型的可以遗传控制的自身免疫性肝病,具有类似人类PBC的病理和生理特征。
NOD.c3c4小鼠模型表现出许多的肝脏组织病理学的异常,包括肝脏淋巴细胞浸润、汇管区炎症反应,以及受损胆管周围嗜酸性粒细胞浸润和上皮肉芽肿的形成。此外,50%-60%的小鼠在9-10周龄自发产生针对PDC-E2的自身抗体[20],80%-90%会产生抗核抗体,胆道上皮受累部位出现CD3+、CD4+、CD8+T细胞浸润。此外,有报道称对NOD.c3c4小鼠采用CD3单克隆抗体处理,可以使其免受自身免疫性肝病的侵害,但是在脾脏的过继转输之后仍然会发生自身免疫性肝病[21]。这表明,T细胞在该疾病的发生中起着关键作用。
该模型与人类PBC不同的一个方面是胆道上皮细胞的增生,在该模型中,胆管损伤和淋巴细胞的浸润先于胆管上皮细胞的增殖[3]。该模型具有比人类PBC更广泛的胆管增生改变和自身免疫攻击部位,NOD.c3c4小鼠以胆总管和肝内胆管为靶标,而PBC主要攻击肝内中小胆管。
尽管NOD.c3c4小鼠模型具有许多和人类PBC相似的组织病理表型,但该模型仍存在AMA阳性率较低,血清ALP无法测得等缺点,此外该模型小鼠发病不存在性别偏倚。
2.4.2dnTGFβRⅡ小鼠模型 在NOD.c3c4小鼠模型造模成功后,研究者发现TGFβⅡ型受体显性抑制(dominant-negative transforming growth factor-βreceptor type Ⅱ,dnTGFβRⅡ)转基因小鼠模型可自发产生和人类PBC相似的组织病理学表现[22]。TGFβⅡ型受体是TGFβ在信号转导的过程中至关重要的受体,它能够调节淋巴细胞的活化。dnTGFβRⅡ小鼠在CD4启动子的作用下过度表达TGFβ受体Ⅱ型显性抑制基因,最终导致小鼠部分T细胞中的TGFβ信号缺失。而T细胞中的TGFβ信号维持了周围Treg细胞,并以细胞自主的方式抑制T细胞的增殖、激活和分化,缺乏这种信号将会导致小鼠严重的自身免疫和早期死亡。
该模型小鼠针对人类PBC中的主要自身抗原PDC-E2、OGDC-E2以及BCOADC-E2自发产生AMA,并且呈100%阳性。dnTGFβRⅡ小鼠的肝脏组织学表明100%的小鼠在肝实质和门静脉区有类似于人类PBC的CD4+、CD8+以及CD19+T淋巴细胞的浸润。并且25%-50%的22周龄的小鼠,会产生胆管损伤,这一点也可见于人类的PBC[23]。此外,与人类PBC患者相似,dnTGFβRⅡ小鼠的肝脏中CD8+/CD4+T细胞的水平升高,血清中IFN-γ、TNF-α、IL-6和IL-12 p40水平均显著增加[22]。dnTGFβRⅡ小鼠表现出和人类PBC相似的主要血清学和组织学特征,这表明TGFβ信号转导在PBC的发病中可能起着至关重要的作用。
dnTGFβRⅡ小鼠也具有许多与人类PBC的不同之处,该模型小鼠不存在类似于人类PBC的性别偏倚,肝内无嗜酸性粒细胞浸润,肉芽肿形成也不明显。
此前有报道,将dnTGFβRⅡ小鼠脾脏的CD4+以及CD8+T细胞过继转移到Rag1-/-小鼠体内,结果Rag1-/-小鼠出现了类似人类的PBC疾病表现,表明脾脏的T细胞以及B细胞的耐受性的丧失导致了PBC。为了进一步研究T细胞在PBC发病中的作用,分别将CD4+以及CD8+T细胞转移到Rag1-/-小鼠体内,结果CD4+T细胞转移的Rag1-/-小鼠出现了结肠炎,而CD8+T细胞转移的Rag1-/-小鼠出现了类似于人类PBC的疾病表现,这表明,CD8+T细胞在胆管损伤中起着至关重要的作用[23]。将dnTGFβRⅡ小鼠与B细胞缺陷小鼠(Igμ-/-小鼠)进行杂交,用以评估B细胞在肝脏的炎症浸润以及伴随的结肠炎中的作用。结果表明,Igμ-/-dnTGFβRⅡ小鼠表现出更严重的胆管损伤以及结肠炎,提示B细胞在dnTGFβRⅡ小鼠中具有抑制炎性浸润的作用[24]。为了进一步确认B细胞在dnTGFβRⅡ小鼠组织病理中的作用,对幼龄小鼠(4-6周)以及成年小鼠(20-22周)分别进行B细胞耗竭治疗。结果表明,幼龄小鼠在经过治疗后,其肝脏的炎症浸润情况以及血清AMA水平明显降低,但是结肠炎有加重倾向,同样对于已经发病的成年小鼠进行治疗则无效,提示,B细胞在PBC的发病过程中起着双向的作用[25]。除了在T、B细胞的研究上提供了新的思路外,dnTGFβRⅡ小鼠也被应用于CD-1d限制NKT细胞、IFN-γ和IL-12在PBC病理过程中的作用的研究,为PBC的病理学和免疫学研究中提供了新的思路和方法。
2.4.3IL-2R-/-小鼠模型 白细胞介素-2(interleukin-2,IL-2)对于CD4+、CD25+Treg细胞亚群的发育和外周扩张起着至关重要的作用,而CD4+、CD25+Treg细胞可以通过抑制T细胞的体内反应提高机体的自我耐受性。
将C57BL/6J背景来源的小鼠敲除IL-2R基因后,IL-2R-/-小鼠产生与慢性非化脓性破坏性胆管炎相似的血清学和病理学特征,这也可见于人类PBC。IL-2R-/-小鼠血清学检测呈现出100%的AMA的阳性,此外,IL-2R-/-小鼠的汇管区炎性细胞浸润增多,并且CD8+/CD4+T细胞的比例也有所升高。与dnTGFβRⅡ小鼠的过继转移实验所表明的结果相同,敲除CD4+T细胞的IL-2R-/-实验小鼠表现出结肠炎的发生以及严重程度的降低,而敲除CD8+T细胞的IL-2R-/-结肠炎的小鼠则表现出缺乏胆管损伤的病理学表现,同样提示CD8+T细胞是PBC发病过程中的关键细胞[26]。
值得注意的是,IL-2R-/-小鼠表现出严重的自身免疫性胆管炎的同时,也伴随发生全身的肠道疾病,25%-50%的小鼠在8-20周龄的时候死于严重的溶血性贫血。
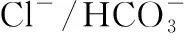
AE2a,b-/-小鼠表现出和人类PBC相似的疾病表现,血清学检测发现除AMA升高外,IgG、IgM以及ALP的水平均升高。此外,该模型小鼠还表现出脾肿大,IL-12 p70和IFN-γ的水平升高,CD4+Foxp3+Treg表达减少。约1/3的小鼠会发生胆管损伤,损伤胆管周围以及汇管区可见CD4+、CD8+T细胞浸润[27]。
虽然AE2a,b-/-小鼠在研究PBC的发病机制方面起到了一定的作用,但是该模型仍存在发病迟缓、不存在性别偏倚、难以繁衍以及AMA阳性率低等问题,为其应用带来了一定的限制。
2.4.5ARE-Del-/-小鼠模型 ARE-Del-/-小鼠模型和人类PBC具有高度相似的疾病表现,该模型是通过敲除IFN型基因3'非翻译区中具有162 nt的腺嘌呤尿嘧啶富含元件(adenylate uridylate-rich element,ARE)而形成的[28]。
ARE-Del-/-小鼠存在着类似人类PBC发病的女性偏倚现象,20周龄的雌性小鼠的肝脏可观察到中度到重度的淋巴细胞浸润,而同样条件下的雄性小鼠只有轻度到中度的淋巴细胞浸润。此外,雌性ARE-Del-/-小鼠的汇管区和肝小叶炎症、小胆管的破坏较严重,也可观察到明显的肉芽肿的形成,雌性小鼠还可见轻度的肝纤维化,而雄性小鼠则无法观测到。在血清学特征上,8-10周龄的雌性小鼠可观察到抗PDC-E2、OGDC-E2以及BCOADC-E2等自身抗体,其中主要是抗PDC-E2自身抗体。而在20周龄时,雌性小鼠AST、ALT以及总胆汁酸水平较雄性小鼠均明显升高。
报道称,将ARE-Del-/-小鼠CD4+T细胞转移至B6/Rag1-/-小鼠体内,引起受体小鼠发生汇管区以及肝实质的炎症浸润,并且胆管损伤和肉芽肿的形成降到最低,此外,在CD4+T细胞转移后,受体小鼠血清IFN-γ、TNF-α以及IL-6的水平均显著升高,而采用相同方法将ARE-Del-/-小鼠的CD8+T细胞转移至B6/Rag1-/-小鼠后,受体小鼠仅发生轻度的汇管区和肝实质炎性浸润,无肉芽肿形成,IFN-γ、TNF-α以及IL-6等血清学指标也无显著变化,提示CD4+T细胞在ARE-Del-/-小鼠的发病机制中起着关键的作用。性别偏倚是ARE-Del-/-小鼠模型的优势之一,在雌性小鼠中,差异表达的基因具有更强的Ⅰ型和Ⅱ型干扰素信号和淋巴细胞介导的免疫反应,可能是导致该模型小鼠产生类似人类PBC的女性偏倚的原因。总之,改变IFN-γ表达在PBC的发病机制中至关重要[29]。
3 总结
PBC是一种以AMA的存在和肝内胆管进行性破坏为特征的肝脏特异性自身免疫性疾病,其发病过程可能涉及环境、遗传易感性、免疫耐受异常等多种因素。由于PBC的发病涉及多种因素,其临床过程可能较为复杂,因此,利用动物模型对阐明PBC的发病机制具有一定的价值。

Tab 1 Comparison between human PBC and murine models of PBC
本文综述了手术模型、药物诱导模型、抗原免疫模型以及基因自发小鼠模型,虽然目前针对PBC的动物模型种类繁多,但也正因PBC的发病机制的复杂性,现有动物模型还不能够完全模拟人类PBC的疾病表现,每一种动物模型在模拟人类疾病上都有其独特的优势和不足。手术模型制备简单、成本低廉,在研究胆汁淤积方面方法成熟、成模率高;药物诱导模型通过药物来诱导小鼠自身免疫耐受失调,最终导致发生类似人类PBC的疾病表现,该类模型的制备验证了环境因素在PBC发病中的关键作用;而抗原免疫模型利用自身抗原或和自身抗原具有氨基酸同源性的异种抗原免疫小鼠,强调了在PBC发病过程中,自身抗原以及感染因素的作用;基因自发模型均可自发产生类似人类PBC的疾病特征,在研究天然免疫和适应性免疫在PBC发病过程中的作用有着独特的优势,但该类小鼠成本较高,并且大多数的基因自发小鼠存在着难以繁衍等问题。事实上,每一种动物模型都各有优劣,由于PBC发病机制的复杂性,任何单一的动物模型都很难完全模拟其疾病表现,但也正是这些各有优劣的动物模型的共同使用研究,让我们对PBC有了更进一步的了解。而正因为我们目前对PBC的了解,将会促进我们在未来开发更加合适匹配的动物模型,也将促进对PBC发病机制的进一步探索和治疗药物研发。
猜你喜欢
传染病信息(2022年2期)2022-07-15
昆明医科大学学报(2022年4期)2022-05-23
中国典型病例大全(2022年12期)2022-05-13
生物医学工程学进展(2018年1期)2018-03-26
西南医科大学学报(2015年1期)2015-08-22
医学研究杂志(2015年6期)2015-07-01
癌变·畸变·突变(2015年3期)2015-02-27
现代检验医学杂志(2015年6期)2015-02-06
郑州大学学报(理学版)(2014年2期)2014-03-01
中国医学科学院学报(2013年2期)2013-03-11
