
LncRNA TMPO-AS1调控miR-143-3p的表达影响肺癌细胞增殖、侵袭、迁移和凋亡
2021-09-01 09:41:58周斌锋彭兵锋王志强喻国平张小强
中国老年学杂志 2021年15期
周斌锋 彭兵锋 王志强 喻国平 张小强
(1鹰潭市人民医院心胸外科,江西 鹰潭 335000;2南昌大学附属第二医院胸外科)
肺癌是临床常见恶性肿瘤之一,据调查研究显示全世界范围内肺癌发病率持续升高,其中非小细胞肺癌是肺癌的主要病理类型,目前临床常采用手术、化疗等手段治疗肺癌,但肺癌患者生存率仍未明显升高〔1〕。近年来,免疫治疗与分子靶向治疗成为研究热点,因而深入探究肺癌发生的分子生物过程有助于寻找新的治疗靶点以此提高患者生存率。长链非编码RNA(LncRNA)TMPO-AS1可促进非小细胞肺癌、前列腺癌等肿瘤发生及发展〔2,3〕。但TMPO-AS1在肺癌发生及发展过程中的分子机制尚未完全阐明。LncBase v.2网站预测显示微小RNA-143-3p(miR-143-3p)可能是TMPO-AS1的靶基因,研究表明miR-143-3p在结肠癌、宫颈癌、喉部鳞状细胞癌等恶性肿瘤中均呈低表达,上调其表达可抑制肿瘤细胞增殖及转移〔4~8〕。但TMPO-AS1是否可通过靶向miR-143-3p促进肺癌发生及发展尚未可知。因此,本研究主要探讨TMPO-AS1与miR-143-3p在肺癌细胞中的相互作用关系,初步分析其对肺癌细胞增殖、迁移、侵袭及凋亡的影响,为肺癌靶向治疗提供潜在靶点。
1 材料与方法
1.1材料与试剂 BEAS-2B、肺癌细胞A549、SPC-A-1、H446、PC-9均购自美国ATCC细胞库。DMEM培养基、胎牛血清、胰蛋白酶均购自美国Gibco公司;二甲基亚砜(DMSO)购自美国Sigma公司;Lipofectamine2000购自美国Invitrogen公司;Matrigel基质胶购自美国BD公司;TMPO-AS1小干扰RNA(si-TMPO-AS1)、si-con、miR-143-3p mimics、miR-con、miR-143-3p抑制剂(anti-miR-143-3p)、anti-miR-con均购自上海吉玛制药技术有限公司;pcDNA3.1购自北京合生基因科技有限公司;Transwell小室购自美国Corning公司;MTT细胞增殖试剂盒购自美国Sigma公司;Annexin V-FITC/PI双染法细胞凋亡检测试剂盒购自北京宝赛生物技术有限公司;双荧光素酶报告基因检测试剂盒购自北京全式金生物技术有限公司;Trizol、反转录及qRT-PCR试剂盒均购自大连宝生物工程有限公司;兔抗人PCNA、基质金属蛋白酶(MMP)-2多克隆抗体均购自美国Santa Cruz公司;兔抗人酶切含半胱氨酸的天冬氨酸蛋白水解酶(caspase)-3抗体购自美国Abcam公司;辣根过氧化物酶(HRP)标记的山羊抗兔IgG二抗购自武汉博士德生物工程有限公司。
1.2方法
1.2.1细胞转染及分组 BEAS-2B细胞与肺癌细胞A549、SPC-A-1、H446、PC-9均培养于含有10%胎牛血清及青霉素-链霉素混合溶液的DMEM培养基,置于37℃、5%CO2培养箱内培养,待细胞生长至80%左右时进行传代培养,取对数生长期肺癌SPC-A-1、H446细胞,0.25%胰蛋白酶消化细胞,制备细胞悬液,接种于6孔板,置于37℃、5%CO2培养箱内继续培养,待细胞生长融合至50%时,参照Lipofectamine2000试剂说明书进行转染,转染前更换为Opti-MEM减血清培养基,实验将肺癌SPC-A-1、H446细胞随机分为si-con组(si-con分别转染入SPC-A-1、H446细胞)、si-TMPO-AS1组(si-TMPO-AS1分别转染入SPC-A-1、H446细胞)、miR-con组(miR-con分别转染入SPC-A-1、H446细胞)、miR-143-3p组(miR-143-3p mimics分别转染入SPC-A-1、H446细胞)、si-TMPO-AS1 +anti-miR-con组(si-TMPO-AS1与anti-miR-con分别共转染入SPC-A-1、H446细胞)、si-TMPO-AS1 +anti-miR-143-3p组(si-TMPO-AS1 与anti-miR-143-3p分别共转染入SPC-A-1、H446细胞),各组细胞转染6 h后更换为含有10%胎牛血清及青霉素-链霉素混合溶液的DMEM培养基继续培养48 h,收集对数生长期细胞进行后续功能验证。
1.2.2qRT-PCR检测细胞中TMPO-AS1、miR-143-3p表达水平 采用Trizol法提取细胞总RNA,应用NanoDrop2000分光光度计检测RNA浓度,取2μg RNA参照反转录试剂盒说明书将其反转录为cDNA,qRT-PCR检测TMPO-AS1、miR-143-3p的表达水平,反应体系:2×SuperReal PreMix Plus 10 μl,上下游引物各1 μl,cDNA模板2 μl,ddH2O补足体系至20 μl;反应条件:95℃ 2 min(循环1次),95℃ 30 s,60℃30 s,72℃ 30 s(循环40次),检测miR-143-3p表达时以U6为内参,TMPO-AS1以β-actin为内参,采用2-ΔΔCt法计算TMPO-AS1、miR-143-3p相对表达量。
1.2.3噻唑蓝(MTT)检测细胞增殖 转染后各组肺癌SPC-A-1、H446细胞按照每孔5 ×103个细胞的密度接种于96孔板,同时设置空白孔(培养液、无细胞),每孔加入20 μl MTT溶液(5 mg/ml),继续培养4 h,弃培养液,每孔加入150 μl DMSO,低速振荡,充分混匀,应用酶标仪检测波长为490 nm处各孔吸光度值(OD),计算细胞存活率,细胞存活率(%)=〔(实验组OD值-空白对照组OD值)/(正常培养组OD值-空白对照OD值)〕×100%。
1.2.4克隆形成实验 收集各组肺癌SPC-A-1、H446细胞,用胰蛋白酶消化细胞,4℃,1 000 r/min转速离心3 min(离心半径12 cm),弃上清,加入无菌磷酸盐缓冲液(PBS)5 ml,洗涤细胞沉淀,相同条件下再次离心,反复洗涤3次,加入培养基,制备单细胞悬液,调整细胞浓度为1×103个/ml,单细胞悬液接种于6孔板进行克隆形成培养,每3 d更换一次培养,继续培养14 d后观察细胞克隆形成情况,实验设置3次重复。
1.2.5流式细胞术检测细胞凋亡 分别收集各组肺癌SPC-A-1、H446细胞,胰蛋白酶消化细胞,4℃,1 000 r/min转速离心5 min,弃上清,加入预冷PBS洗涤2次,每次5 min,弃上清,涡旋振荡30 s,制备细胞悬浮液,加入200 μl结合缓冲液,再加入10 μl Annexin V-FITC。振荡混匀,冰上避光孵育30 min,加入5 μl PI,涡旋振荡混匀,冰上孵育5 min,加入400 μl结合缓冲液,涡旋振荡混匀,使用300目尼龙网过滤,于1 h内应用流式细胞仪检测各组细胞凋亡情况。
1.2.6Transwell实验检测细胞迁移及侵袭 细胞侵袭实验:肺癌SPC-A-1、H446细胞培养于含有0.1%胎牛血清的DMEM培养基,培养24 h后,用不含胎牛血清的DMEM培养基重悬细胞,取2×105个细胞接种于含有基质胶的Transwell小室上室中(直径6.5 μm,孔径8 μm),取500 μl含有10%胎牛血清的DMEM培养基置于Transwell小室的下腔中,置于37℃、体积分数5%CO2培养箱机修培养24 h,去除上膜表面上的细胞,用多聚甲醛固定20 min,0.5%结晶紫染色液染色10 min,无菌水清洗后置于倒置显微镜下随机选取10个视野计数。细胞迁移实验:用不含胎牛血清的DMEM培养基重悬细胞,取2×105个细胞接种于Transwell小室上室中,其余实验步骤同细胞侵袭实验,实验均设置3次生物学重复。
1.2.7双荧光素酶报告基因检测 构建TMPO-AS1携带的3′UTR的荧光素酶报告因子,每个基因的结合位点与miR-143-3p结合位点相对应,荧光素酶测定显示TMPO-AS1与miR-143-3p存在结合位点,将结合位点与突变位点分别载入荧光素酶报告基因载体,分别为野生型WT-TMPO-AS1、突变型MUT-TMPO-AS1,WT-TMPO-AS1、MUT-TMPO-AS1分别与miR-143-3p mimics、miR-con分别共转染入肺癌SPC-A-1、H446细胞,继续培养24 h,应用双荧光素酶报告基因测定系统检测荧光素酶活性,实验重复3次。
1.2.8Western印迹检测PCNA、MMP-2、酶切caspase-3蛋白表达 分别取各组肺癌SPC-A-1、H446细胞,加入蛋白裂解液,冰上裂解30 min,12 000 r/min转速离心10 min(离心半径6 cm)提取细胞总蛋白,采用BCA法测定蛋白浓度,蛋白煮沸变性,取变性完成蛋白样品30 μg/孔加入十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)的加样孔内,经SDS-PAGE分离蛋白,转移至聚偏氟乙烯(PVDF)膜,5%脱脂奶粉封闭1 h,加入一抗(1∶1 000),4℃轻摇24 h,TBST洗膜3次,每次5 min,加入二抗(1∶3 000),室温孵育2 h,TBST洗膜3次,每次10 min,避光加入ECL显色液,曝光,显影,凝胶分析系统及ImageJ软件分析条带灰度值。
1.3统计学方法 应用SPSS21.0软件进行t检验、单因素方差分析。
2 结 果
2.1TMPO-AS1和 miR-143-3p在肺癌细胞中的表达 与BEAS-2B相比,肺癌细胞A549、SPC-A-1、H446、PC-9中TMPO-AS1的表达水平均显著升高(P<0.05),而肺癌细胞A549、SPC-A-1、H446、PC-9中 miR-143-3p的表达水平均显著降低(P<0.05),见表1。其中肺癌SPC-A-1、H446细胞中TMPO-AS1的表达水平相比于其他肺癌细胞系明显升高(均P<0.05),因而本研究选取肺癌SPC-A-1、H446细胞进行后续实验。
表1 检测肺癌细胞中TMPO-AS1和 miR-143-3p表达
2.2沉默TMPO-AS1或过表达 miR-143-3p对肺癌细胞增殖的影响 与si-con组相比,si-TMPO-AS1组肺癌SPC-A-1、H446细胞存活率及克隆形成率显著降低(P<0.05),PCNA蛋白的表达水平显著降低(P<0.05);相较于miR-con组,miR-143-3p组肺癌SPC-A-1、H446细胞存活率、克隆形成率与PCNA蛋白的表达水平均显著下降(P<0.05),见图1、表2。

图1 检测肺癌细胞中PCNA蛋白表达

2.3沉默TMPO-AS1或过表达 miR-143-3p对肺癌细胞凋亡的影响 流式细胞术检测结果显示,si-TMPO-AS1组肺癌SPC-A-1、H446细胞凋亡率均显著高于si-con组(均P<0.05),酶切caspase-3表达水平均显著升高(均P<0.05);相较于miR-con组,miR-143-3p组肺癌SPC-A-1、H446细胞凋亡数显著增多(均P<0.05),酶切caspase-3的表达均上调(均P<0.05),见图2、表3。
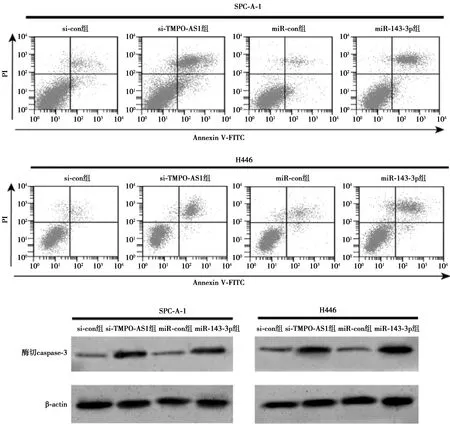
图2 流式细胞术检测细胞凋亡和Western印迹检测肺癌细胞中酶切caspase-3蛋白表达
2.4沉默TMPO-AS1或过表达 miR-143-3p对肺癌细胞迁移和侵袭的影响 肺癌SPC-A-1、H446细胞中沉默TMPO-AS1或 miR-143-3p过表达后,与si-con组比较,si-TMPO-AS1组细胞迁移及侵袭均数显著减少(均P<0.05),MMP-2蛋白的表达水平显著降低(P<0.05);与miR-con组比较,miR-143-3p组细胞迁移及侵袭数显著减少(P<0.05),MMP-2蛋白的表达水平显著降低(P<0.05),见图3、图4、表4。

图3 Transwell检测肺癌细胞迁移和侵袭(结晶紫染色,×200)

图4 Western印迹检测各组MMP-2表达
2.5TMPO-AS1靶向调控 miR-143-3p的表达 LncBase v.2网站预测显示TMPO-AS1与 miR-143-3p存在互补靶向序列,见图5。双荧光素酶报告实验结果显示,肺癌SPC-A-1、H446细胞中转染克隆有TMPO-AS1-3′UTR突变型载体质粒实验中,miR-143-3p组与miR-con组比较,荧光素酶活性差异无显著性;转染克隆有TMPO-AS1-3′UTR载体质粒实验中,miR-143-3p组荧光素酶活性明显受到抑制,与miR-con组比较,荧光素酶活性差异具有显著性(P<0.05),见表5。

图5 TMPO-AS1与 miR-143-3p存在互补靶向序列
表5 双荧光素酶报告实验
qRT-PCR检测结果显示,si-TMPO-AS1组肺癌SPC-A-1、H446细胞中miR-143-3p的表达水平显著高于si-con组,差异有统计学意义(P<0.05),pcDNA-TMPO-AS1组肺癌SPC-A-1、H446细胞中miR-143-3p的表达水平显著低于pcDNA组,差异有统计学意义(P<0.05),见表6。
表6 LncRNA TMPO-AS1调控 miR-143-3p的表达
2.6干扰 miR-143-3p表达逆转沉默TMPO-AS1对肺癌细胞的抑制作用 相较于si-TMPO-AS1+anti-miR-con组,si-TMPO-AS1+anti-miR-143-3p组肺癌SPC-A-1、H446细胞存活率与克隆形成率均显著升高,差异有统计学意义(均P<0.05),迁移细胞数与侵袭细胞数均显著增加,差异有统计学意义(均P<0.05),细胞凋亡率显著降低,差异有统计学意义(P<0.05),PCNA、MMP-2蛋白的表达水平均显著升高,差异有统计学意义(均P<0.05),而酶切caspase-3蛋白的表达水平显著降低,差异有统计学意义(P<0.05),见图6、表7、表8。

1~4:si-con组、si-TMPO-AS1组、si-TMPOAS1+anti-miR-con组、si-TMPOAS1+anti-miR-143-3p组图6 Western印迹检测肺癌细胞中酶切caspase-3、PCNA和MMP-2蛋白表达


3 讨 论
大部分肺癌患者确诊时已处于中晚期,经临床治疗后患者预后较差〔9〕。因而急需寻找早期诊断及监测肺癌的新方法。研究表明LncRNA可能通过调控miRNA而参与肺癌发生及发展过程〔10,11〕。但仍有部分LncRNA-miRNA在肺癌发生过程中的作用机制尚未完全阐明。本研究深入分析TMPO-AS1与miR-143-3p在肺癌发生发展过程中的可能作用机制。
TMPO-AS1在肺腺癌患者血清中呈高表达,其高表达量与患者预后不良密切相关,研究表明TMPO-AS1还可能竞争性吸附miRNA而参与肺腺癌发生发展过程〔12,13〕。本研究结果提示TMPO-AS1表达水平升高可能促进肺癌的发生,沉默TMPO-AS1表达可显著降低肺癌细胞增殖、迁移及侵袭能力,诱导细胞凋亡。研究表明PCNA可反映肿瘤细胞增殖,其表达水平升高预示细胞增殖能力增强,MMP-2过表达可增强肿瘤细胞迁移及侵袭能力,酶切caspase-3蛋白是细胞凋亡发生过程中的关键因子,其表达量升高预示凋亡细胞数增加〔14~16〕。本研究结果提示沉默TMPO-AS1表达可通过上调Cleaved caspase-3蛋白表达及下调PCNA、MMP-2蛋白表达而促进肺癌细胞凋亡,抑制细胞增殖、迁移及侵袭。
miR-143-3p在乳腺癌细胞中呈低表达,miR-143-3p过表达可通过靶向LIM结构域激酶1的表达而抑制乳腺癌发生及发展〔17〕。研究表明miR-142-3p通过靶向CDC25C,TGFβR1,GNAQ,WASL、RAC1抑制黑色素瘤恶性进展〔18〕。与上述研究结果相似,本研究结果表明miR-143-3p在肺癌细胞中的表达水平显著降低,miR-143-3p过表达与沉默TMPO-AS1后对肺癌细胞增殖、迁移、侵袭及凋亡的作用效果相同,同时本研究通过双荧光素酶报告实验及qRT-PCR实验证明TMPO-AS1可靶向负性调控miR-143-3p的表达,提示TMPO-AS1可能靶向抑制miR-143-3p表达而促进肺癌细胞增殖、迁移、侵袭,抑制细胞凋亡。本研究结果提示沉默TMPO-AS1表达可通过上调miR-143-3p表达而减弱肺癌细胞增殖、迁移、侵袭能力,促进细胞凋亡。
综上,TMPO-AS1可调控miR-143-3p的表达影响肺癌细胞增殖、迁移、侵袭及凋亡,为揭示肺癌发生的分子机制奠定理论基础,可为肺癌的靶向治疗提供新的靶点。
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
农业与技术(2021年22期)2021-11-30 06:41:00
天津医科大学学报(2021年4期)2021-08-21 02:14:50
国际医药卫生导报(2021年9期)2021-07-06 10:17:48
中日友好医院学报(2021年1期)2021-04-14 01:58:32
今日农业(2020年24期)2020-12-15 16:16:00
山东医药(2020年9期)2020-05-20 01:12:16
水动力学研究与进展 B辑(2016年4期)2016-10-18 01:45:25
北方药学(2016年8期)2016-08-09 02:20:45
兽医导刊(2016年12期)2016-05-17 03:51:50
