
基于根系年轮的晋西北土壤侵蚀速率研究
2021-08-31 06:18杨晨欣苏常红李宗善
山西大学学报(自然科学版) 2021年4期
杨晨欣,苏常红*,李宗善
(1. 山西大学 黄土高原研究所,山西 太原 030006;1*. 天津师范大学 地理与环境科学学院,天津 300387;2. 中国科学院 生态环境研究中心,北京 100085)
0 引言
树木根系年轮对周围环境变化的响应十分灵敏,自然环境的突变可能会导致树木根系发生不同程度的暴露。通过树木根系年轮判断微地貌变化发生的时间、位置、强度等信息,可以弥补可达性差的地区因历史资料匮乏造成的数据获取的不便[1],为水土保持研究提供了一种全新的方法与思路。某些具有灾害性的地貌活动会给树木带来毁灭性的打击,但如果强度较小,很多树木会存活下来,但都会受到不同程度的损伤,比如泥石流冲击的痕迹,树木倾斜或折断,而这些损伤都会通过树木年轮的异常变化表现出来,树木年轮的变化为重建自然灾害或人为影响造成的土壤侵蚀事件提供了可行的方法[2-3]。降水、风速、植被覆盖度等自然环境因素及其综合影响,都是造成土壤侵蚀的潜在原因[4]。土壤侵蚀会降低土壤肥力,对下游水库造成泥沙淤积,对人类福祉造成严重的影响[4-5]。
目前基于根系年轮的土壤侵蚀研究相对较少,Ballesteros-Cánovas 等综述了用于土壤侵蚀测量的树木地貌学,重点研究了植被对地貌过程的响应[6]。Stoffel 等回顾了暴露根系对理解侵蚀过程及地貌演化的贡献,认为暴露根系的细胞结构变化是根系失去上覆土层保护后正常的生理反应,即根系暴露后,细胞结构会发生变异以适应新的生存策略,且这种变异普遍存在[7]。孙丽萍等在金沙江干热河谷龙川江流域的研究表明,根系导管面积是反演侵蚀过程的敏感指标,早材导管面积的变异是确定根系暴露时间的重要依据之一[8]。此外研究人员还对不同生境下土壤侵蚀的原因进行了分析识别,张雨以南方暴露马尾松根系为研究对象,探讨了土壤侵蚀强度与各环境因子间的关系。结果表明,区域内以乔灌草复合植被覆盖条件下的样地侵蚀速率最小,人类活动强度对土壤侵蚀剧烈程度有一定的影响,但不及植被覆盖类型的影响大[9]。
黄土高原位于黄河流域中游,土质疏松、土壤颗粒细小,加上地面坡度大、夏季多发暴雨,加速了水土流失,是世界上水土流失最严重的地区之一。晋西北地区在长期流水侵蚀下地面植被稀疏,水土流失严重[10]。本文以黄土高原晋西北地区的天峰坪镇为研究区,通过踏查将小叶杨(Populus simonii)根系定为研究对象,通过将植物解剖结构与土壤侵蚀信息结合起来,对基于树木根系年轮在土壤侵蚀分析的可行性进行探讨,拓展土壤侵蚀研究的方法与思路,为生态脆弱区生态修复在理论与实践方面提供可供借鉴的案例。
1 研究区域概况
研究区位于山西省忻州市偏关县天峰坪镇。偏关县西邻黄河,北接长城,全县总面积为1 685.4 km2,处于山西省西北部与内蒙古自治区的交界处[11]。研究区位于东经111°22′~11°01′,北纬39°12′~39°40′,海拔在900 m~1 000 m。该区域地貌复杂多山,丘陵沟壑纵横,总体呈现东高西低的地势,土壤贫瘠,主要类型为固定风沙土,属北温带大陆性气候,年平均气温5 ℃~9 ℃,年平均降水量为415.3 mm[12]。
研究区树种较为单一,以小叶杨为单一优势物种。通过踏查,选择天峰坪镇2 个较大的有侵蚀痕迹的样地,分别是井沟和蹄子沟,进行调查与采样(图1),样地内杨树根系侵蚀呈现局部暴露、全部暴露、和浅埋多种形式。
2 研究方法
2.1 样本采集
2019 年7 月19 日―2019 年7 月21 日对研究区进行调查采样,共采集小叶杨根系样品13 个。用手锯在距离树干水平距离大于0.5 m 之处采样[13-14],每个根系样本的平均长度约为12 cm ~17 cm,按照根系不同暴露方式分3 种情况进行采样:①正常生长根系(对照样),采集距地表1、3、5 和7 cm的根系(图1a);②局部暴露根系,根系约暴露出2/3 左右(图1b);③浅露根系,根系暴露程度小于根系直径的1/3(图1c)。同时详细记录采样点的地貌特征、采样点距离树干的距离和根系上表皮到土壤表面的垂直距离等。
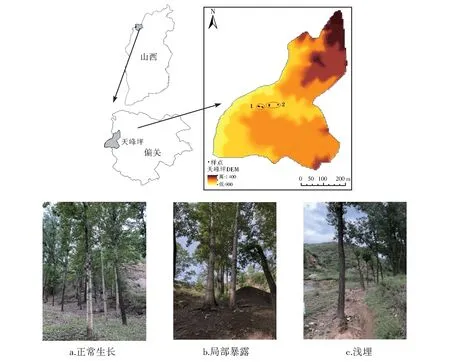
图1 研究区域位置图及采样点实景图Fig. 1 Study area and photograph of sampling sites
2.2 样本处理及分析
将采集的根系样品在干燥阴凉处晾置1 个月左右,晾干后取厚度为3 cm 的根系圆盘。用不同粒径(p240、p400、p800、p1200)的砂纸依次打磨根系圆盘横截面,至年轮清晰可见。处理后的样品使用数码显微镜观测,并沿圆盘直径连续拍摄显微照片。将显微照片用Adobe Photoshop CS6 软件拼接后使用Image-Pro Plus 图像处理软件进行处理,计算平均导管面积、平均早材导管面积和年轮内的导管数量等。小叶杨早晚材之间的分界线不明显,取年轮内前50%导管面积的平均值为平均早材导管面积,根据根系早材导管面积的变化确定根系首次暴露时间。
2.3 土壤侵蚀速率估算方法
本文以小叶杨早材导管面积显著减小、导管排列紧密作为判定根系暴露年份的指标。通过树木根系解剖结构变化计算土壤侵蚀速率需确定2 个变量,即根系暴露厚度和根系暴露年份。参照前人研究方法[15-17],建立基于根系的土壤侵蚀速率估算方程如下(公式(1)、(2),图2):
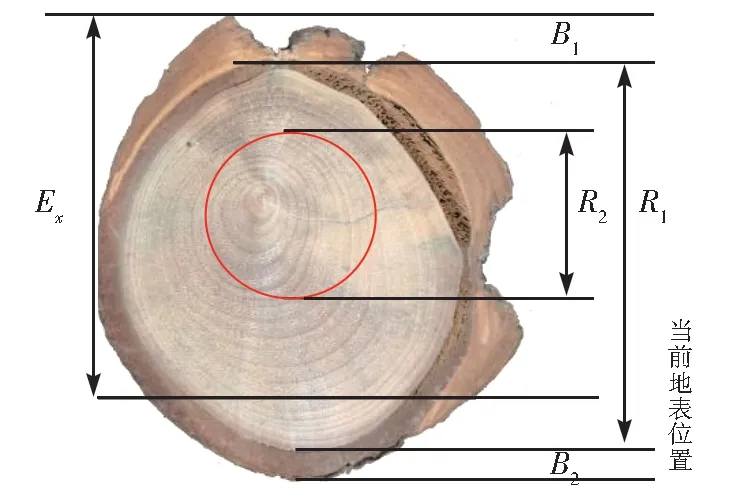
图2 土壤侵蚀速率计算方程示意图Fig. 2 Schematic diagram of equation for calculating soil erosion rate

式中,Er为侵蚀厚度;Ex为根系上表皮距地面的高度;Era为年侵蚀率;R1为当前轮径;R2为树根被侵蚀当年的轮径;B1为根系上表皮的树皮厚度;B2为根系下表皮的树皮厚度;NRex为自暴露至采样时的年轮数目。
2.4 影响因素分析与数据来源
本文对导致研究区土壤侵蚀的因素进行分析,主要分析了降水和风速这两个因素,气象月值数据均下载于中国气象科学数据共享服务网(http://data.cma.cn/),并基于Arcgis 克里格插值法插值得到研究区年平均降水和风速气象数据。
3 结果与分析
3.1 暴露根系年轮解剖结构分析
树木根系早材导管主要是输送水分和无机盐。根据其输送营养物质的效率,可以间接反映根系生长状况、受环境影响程度等信息,且导管对土壤厚度变化的感应十分敏感,可通过其细胞面积变化确定根系开始暴露的时间[18-19]。图3a 为3 号样本的显微照片和早材导管细胞面积的变化图。树木根系2007-2009 年为正常生长,根系显微结构为典型的散孔结构,早晚材导管面积无显著性差异。2010 年早材细胞面积开始显著减小,为2 253 μm2,而该年的降水量达411.74 mm,据此判定该根系样本2010年开始暴露。从2010-2012 年早材导管面积持续减小,直到2012 年减少至1 447 μm2。

图3 暴露根系细胞结构变化图Fig. 3 Structural changes of exposed root cells
图3b 为7 号样本的显微照片和早材导管细胞面积的变化图。根据显微照片和下方折线图可见,2011-2012 年间,早材细胞的面积显著减小,与2011 年相比,平均面积减少了2 454 μm2,因此可判断该根系样本在2012 年暴露,且2012 年之后面积持续下降,土壤持续受到侵蚀。
图3c 为4 号样本的显微照片和早材导管细胞面积的变化图。由显微照片可见,2005-2007 年树木根系为正常生长,根系显微结构属正常散孔结构。折线图表明,2007 年到2008 年,根系导管面积发生骤减,从上一年的5 825 μm2降至3 913 μm2,2008 年之后,根系导管面积保持逐年下降趋势,据此判定该根系样本是在2008 年暴露。
图3d 为8 号样本的显微照片和早材导管细胞面积的变化图。显微照片显示此根系的早材导管细胞面积较其他根系偏大,与4 号样本类似,折线图显示,此样本的早材导管细胞面积也在2007 年到2008 年间发生了骤减,从上一年的21 607 μm2降至11 607 μm2,可判断该根系样本在2008 年暴露。
图3e 为11 号样本的显微照片和早材导管细胞面积的变化图。折线图显示,2008 年之后细胞面积一直维持小幅度波动,2016 年根系导管面积从17 929 μm2骤然减小至8 159 μm2,判定此根系样品在2016 年开始暴露。
3.2 土壤侵蚀速率估算结果
早材导管面积的变异可确定根系首次暴露时间,计算各样本侵蚀量及土壤侵蚀速率如表1 所示,2 号样本的侵蚀量最低,7 号样本的侵蚀量最高,6 号样本的侵蚀速率最低,9 号样本的侵蚀速率最高。

表1 各样本参数及测算结果Table 1 Parameters and measurement results of sample
如表2 所示,虽然研究区两个采样点相距不远,但其受到侵蚀的剧烈程度不同,井沟比蹄子沟的平均土壤侵蚀速率要大。
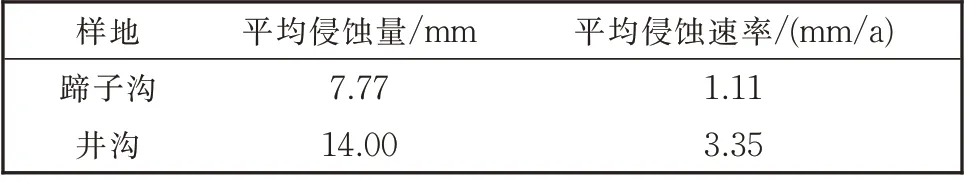
表2 两个样地的土壤侵蚀速率测算结果Table 2 Calculation results of soil erosion rate of two sample plots
3.3 影响因素分析
3.3.1 根系导管细胞面积与降雨量的相关性分析
降水是造成土壤侵蚀的潜在因素[20-21]。根系导管面积越小表示根系暴露程度越大,而根系暴露量越大,则表明土壤侵蚀的程度越重。分析研究区14年的平均降雨量与对应年份的导管细胞面积相关性可知,13 个样本中,除2 号和11 号外,其余样本均呈一定负相关趋势,其中1 号和9 号显著负相关(P<0.05)(表3)。因此,降水是导致研究区根系暴露和土壤侵蚀的主要因素。

表3 根系导管细胞面积与降雨量相关性Table 3 Correlation between root vessel cell areas and rainfall
3.3.2 根系导管细胞面积与风速的相关性分析
风是导致土壤风蚀的直接动力,而风蚀的强弱则取决于风速的大小[22]。表4 为天峰坪镇从2005年到2018 年的导管细胞面积与风速的相关性分析结果,13 个样本中有11 个样本的导管细胞面积与风速呈负相关(P<0.05),其中3 号、4 号、6 号和7 号样本与降雨量呈显著负相关关系,根系导管面积随风速变大而减小,土壤侵蚀程度增加,表明风速也是导致根系暴露和土壤侵蚀的重要因素。

表4 根系导管细胞面积与风速相关性Table 4 Correlation between root vessel cell areas and wind speed
4 讨论
本研究表明,降雨量越高,风速越大,根系暴露越严重,侵蚀量相应也越高,可见降雨和风蚀是造成研究区土壤侵蚀的重要原因。但与降雨量相比,导管细胞面积与风速显著相关的样本数量更多,表明研究区土壤侵蚀的影响因素中,风蚀的影响更为突出。目前在黄土高原土壤侵蚀相关研究中,降雨的侵蚀效应研究较多,对风蚀的研究少见。风蚀对土壤侵蚀的影响是迄需加强的研究方向。
通过比较蹄子沟和井沟两个样地的土壤侵蚀程度,发现蹄子沟的侵蚀度要弱于井沟。其原因在,蹄子沟坡度较小,植被好于井沟,植被冠层可滞留降水,有效阻挡雨水对土层的直接破坏,且林下草本和枯枝落叶层较多,凋落物亦可增加土壤团粒结构稳定性,减少降雨和风蚀对土壤的直接冲刷和刮削;与蹄子沟相比,井沟地形复杂,坡度更大,植被稀疏,灌木和草本植物零星分布,土壤裸露面积大,抗蚀能力弱,导致蹄子沟的土壤侵蚀程度整体小于井沟。
与传统的土壤侵蚀分析方法相比,基于根系年轮的土壤侵蚀测算法有其独有的优势。首先,基于根系年轮的土壤侵蚀分析可以确定根系开始暴露的时间,为探索土壤侵蚀随时间的变化提供了新的思路;此外,根系年轮宽度的变化与环境变化息息相关,为土壤侵蚀驱动机制的深入挖掘提供了可能。另外,与传统方法相比,树木根系年轮的测量和分析相对省时省力[23]。不足之处在于,各种气候或灾害事件的发生会影响树木根系年轮的生长,使其出现缺年、伪轮的情况,影响对树木定年的精确度,进而影响侵蚀率的估算精度[24-25]。因此需要更多的生态水文数据来提高定年准确性,在应用此方法估算土壤侵蚀速率值的同时,降低误差,精确定年。
本研究对研究区土壤侵蚀状况及其影响因素进行了分析,但仍有不足,如对地形、植被、人类活动等对土壤侵蚀影响的分析仍需要进一步探讨,对降雨格局、土壤质地在抗蚀性方面的潜在因素未进行分析,这也是我们未来需要拓展的内容。目前当地对暴露根系的长期监测十分缺乏,这会直接影响我们对研究区土壤侵蚀影响因素的定量评价,因此对该区树木根系生理生长状况的监测工作十分必要,以期通过监测得到精确数据,为生态治理措施的实施提供坚实可信的理论依据。
5 结论
偏关县蹄子沟和井沟的土壤侵蚀量分别为7.77 和14.00 mm,平均土壤侵蚀速率分别为1.11、3.35 mm/a,侵蚀度较高。影响因素分析表明,根系导管细胞面积随降雨量和风速的增大而减小,说明降水和风蚀是导致树木根系暴露和土壤侵蚀的主要因素。
猜你喜欢
现代园艺(2022年7期)2022-11-19
林业科技(2022年5期)2022-10-08
红蜻蜓·低年级(2021年12期)2022-01-19
中国农村水利水电(2021年10期)2021-10-28
化工设计通讯(2021年1期)2021-01-20
魅力中国(2020年23期)2020-12-08
湖北农业科学(2019年22期)2019-12-23
绿色科技(2018年24期)2019-01-19
绿色科技(2018年18期)2018-11-05
小天使·一年级语数英综合(2018年7期)2018-09-12
