
利用单碱基编辑系统定点编辑哈萨克羊MSTN基因的研究
2021-08-23 07:00:56姚旭东蒙亚琦任秀美奥郭延华张译元王立民
畜牧兽医学报 2021年8期
姚旭东,蒙亚琦,任秀美奥,郭延华,唐 红,张译元,王立民*,周 平*
(1.石河子大学动物科技学院,石河子 832000;2. 新疆农垦科学院 省部共建绵羊遗传改良与健康养殖国家重点实验室,石河子 832000)
哈萨克羊是新疆最古老的地方绵羊品种之一,具有耐粗饲、适合放牧、肥育性能好等特点,很好地适应了当地特殊的自然环境[1-2]。近年来,随着圈养规模的增加以及人们对肉品质要求的提高,增加产肉量,提高瘦肉率成为哈萨克羊的育种目标。肌肉生长抑制素(myostatin, MSTN)又称生长分化因子-8(growth differentiation factor-8, GDF-8),是转化生长因子-β(transforming growth factor-β, TGF-β)超家族的一员,参与抑制肌肉分化和生长[3-4]。MSTN基因突变导致其蛋白功能丧失,从而增加肌肉细胞的数量以及增大肌肉纤维的直径,使肌肉过度发育[5]。在家畜中,MSTN基因存在着一些有益的自然突变,使其表现出双肌性状,如双肌肉牛品种比利时蓝牛的MSTN基因第三外显子有11个碱基(937~947 bp)缺失,使MSTN蛋白翻译提前终止;皮埃蒙特牛第3外显子发生g.938G>A替换,这种突变破坏了TGF-β超家族的典型半胱氨酸结构,并导致MSTN蛋白失活[6];双肌肉羊品种特塞尔绵羊(Texel)MSTN基因3′UTR区存在g.6723G>A突变,为骨骼肌中高度表达的microRNAs(miR1和 miR206)创建了靶点,引起MSTN翻译抑制[7]。人为敲除MSTN基因会导致绵羊[8-9]、牛[10]和山羊[11]等动物的肌肉增加、体脂率降低。因此,运用基因编辑技术敲除MSTN基因培育双肌品种成为畜牧业育种的一个新目标。
目前,应用最普遍的基因编辑技术是CRISPR/Cas9(clustered regularly interspersed short palindromic repeats(CRISPR)-associated protein 9)技术,在引导RNA(small guide RNA, sgRNA)的作用下,Cas9酶可以在靶位点造成DNA双链断裂(double-strand break, DSB),然后利用细胞自身的2种修复机制——非同源末端连接(nonhomologous end joining, NHEJ)和同源性定向修复(homology directed repair, HDR)机制实现基因编辑。其中,NHEJ修复机制会引入碱基对插入和缺失(indels)[12];而HDR修复机制结合外源模板可以实现基因组精确编辑,但HDR机制较为复杂,且对细胞周期(在G2/M中更具活性)有较强的依赖性,其效率通常很低[13]。因此,在特定位点引入精确的遗传变异仍然具有挑战性。
碱基编辑器(base editors, BEs)是近年来发展迅速的基因编辑工具之一,其中的胞嘧啶碱基编辑器(cytidine base editor, CBE)主要由胞苷脱氨酶(cytidine deaminase)、尿嘧啶DNA糖基化酶抑制剂(uracil DNA glycosylase inhibitor, UGI)、Cas9蛋白和sgRNA组成,可以在无需双链断裂的情况下靶向诱导胞嘧啶(C)向胸腺嘧啶(T)或鸟嘌呤(G)向腺嘌呤(A)的精准编辑[14]。腺嘌呤碱基编辑器(adenine base editor, ABE)是在CBE的基础上开发的,其结构和碱基编辑机制均类似于CBE,只是腺苷脱氨酶(adenine deaminase)取代了胞嘧啶脱氨酶,可以靶向诱导胞嘧啶A向G或T向C的转变[15]。CBE可以通过将4个密码子(CAA、CAG、CGA或TGG)精确地转换成终止密码子阻止基因翻译,这为基因敲除提供了一种新方法[16],并且不会产生因双链DNA断裂导致的过度DNA损伤和细胞死亡[17]。另外,与CRISPR/Cas9依赖HDR实现精确编辑基因组序列相比,BEs不需要外源供体模板并且不依赖于细胞周期,因此,可以广泛用于不同的细胞类型。近年来,BEs被不断优化改进,并且已经产生了具有更高效率、更窄编辑框和更宽泛PAM识别能力的众多变体,使其应用范围更广[18-19]。迄今为止,BEs已在多种动物(如小鼠、兔子、猪、绵羊和猴子等)中获得成功[20-24]。
经检测,哈萨克羊自然情况下不存在MSTN有益突变,因此,可以利用单碱基编辑技术编辑该品种MSTN基因,定向引入突变,以期提高产肉量。本试验利用CBE系统在MSTN基因编码区提前引入终止密码子,建立敲除MSTN基因的单碱基编辑体系,为培育MSTN基因编辑哈萨克羊奠定基础,为生产产肉率高、体脂率低的哈萨克羊新品种提供新材料。
1 材料与方法
1.1 材料与试剂
pCMV-AncBE4 max-P2A-GFP质粒购自上海吉荧生物技术有限公司;pGL3-U6-sgRNA-PGK-puromycin为省部共建绵羊遗传改良与健康养殖国家重点实验室提供。100 bp DNA Ladder和高保真酶(Pyrobest DNA Polymerase)均购自宝生物工程(大连)有限公司;制性内切酶BsaI和T4 DNA Ligase购自美国NEB公司;CruiserTM酶检测试剂盒(Cat.No.GP0105)购自江苏吉锐公司;DMEM BASIC培养基购自赛默飞世儿科技(中国)有限公司;胎牛血清购自Biologcal Industries公司;细胞裂解液(QuickExtractTMDNA Extraction Solutio, Cat.No.QE09050)购自美国Epicentre公司;T载体购自美国Promega公司;嘌呤霉素购自美国Sigma公司;无内毒素质粒小抽提取试剂盒II型(D6950-01)及Gel & PCR Clean Up Kit购自美国OMEGA公司。葡萄糖、MgCl2·6H2O、NaHCO3、KH2PO4均购自日本和光株式会社。
1.2 试验方法
1.2.1 哈萨克羊胎儿成纤维细胞的原代培养 通过手术法取出妊娠30 d的哈萨克羊胎儿,放置于含有200 IU·mL-1青霉素与链霉素的生理盐水中带回实验室,取出胎儿用1%PBS清洗3次,然后用75%的无水乙醇消毒30 s,再用1%PBS清洗3次,放入培养皿中,剥取胎儿身体的皮肤剪碎,均匀铺在10 cm培养皿中,在培养箱放置3 h后加入8 mL完全培养基(DMEM+15% FBS+1%双抗),放入5% CO2,37 ℃的培养箱中培养,每2 d换一次液,细胞晕形成后除去组织块(记为d1代)。
1.2.2 sgRNA及检测引物设计 针对绵羊MSTN基因外显子序列(序列号:DQ530260.1)设计sgRNA(图1)。pCMV-AncBE4 max-P2A-GFP质粒识别间隔相邻基序(proto-spacer adjacent motifs, PAM)为NGG,sgRNA长度设为20 bp,为便于连接到载体中,在正义链5′端添加ACCG,反义链5′端添加AAAC,共设计2条sgRNAs(表1)。用Primer Premier 5.0设计靶位点检测引物,STOPsg-1位点检测引物为Exon-2F、Exon-2R;STOPsg-2位点检测引物为Exon-3F、Exon-3R(表1)。
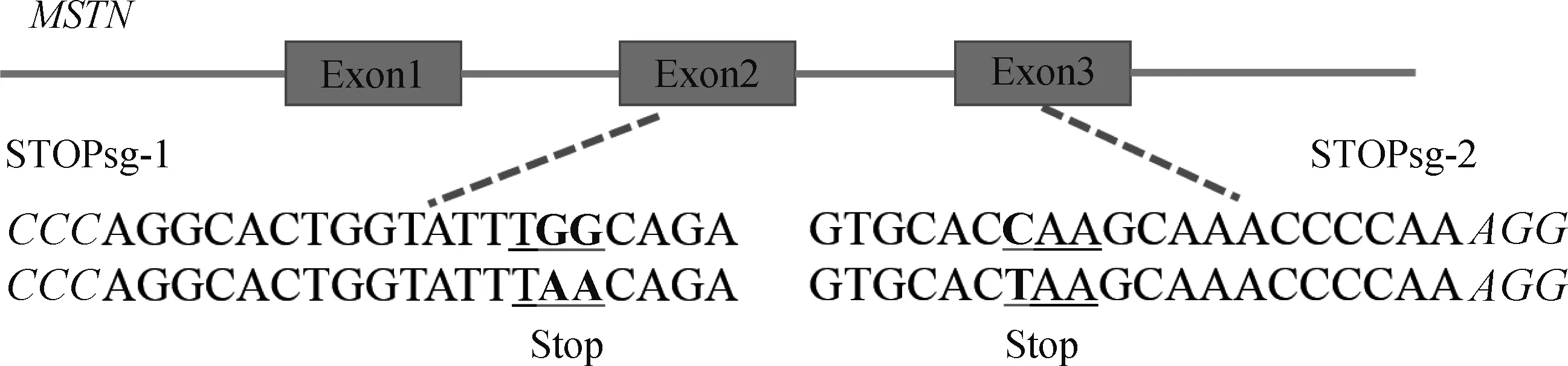
PAM区域标为斜体,靶位点标为加粗黑体,突变氨基酸带下划线Protospacer adjacent motif (PAM) region in italic, target sites in bold, and mutant amino acids in underscore图1 哈萨克羊MSTN基因靶位点的示意图Fig.1 Schematic diagram of the target site of MSTN gene in the Kazakh sheep
1.2.3 MSTN-sgRNA表达载体构建 合成的sgRNA引物分别稀释成100 μm·L-1后形成双链,反应体系如下:正义链和反义链各1 μL,10×T4 Ligation Buffer 1 μL,T4 PNK 0.5 μL,ddH2O 6.5 μL。反应条件:95 ℃ 5 min,95~85 ℃(-2 ℃·s-1),85~25 ℃ (-0.1 ℃·s-1),4 ℃保存。pGL3-U6-sgRNA-PGK-puromycin质粒经内切酶BsaI线性化后胶回收纯化。然后将退火后的sgRNA连接至pGL3-U6-sgRNA-PGK-puromycin质粒,反应体系:双链sgRNA 2 μL,线性化pGL3-U6-sgRNA-PGK-puromycin质粒50 ng,10×T4连接酶buffer 1 μL,T4连接酶1 μL,补加ddH2O至10 μL,置于恒温干浴锅中16 ℃过夜连接。连接好的MSTN-sgRNA表达载体转化后用PCR进行鉴定,上游引物用A-F(表1),下游引物用sgRNA互补链,选取3个 PCR鉴定阳性的质粒送测序进一步鉴定。
1.2.4 电转液的配制 Solution 1: MgCl2·6H2O 1.2 g,ATP-Na22 g,溶于10 mL ddH2O中,0.22 μm滤器过滤除菌,分装后-20 ℃保存。 Solution 2:葡萄糖0.02 g,NaHCO30.06 g,KH2PO40.6 g,溶于50 mL ddH2O中,用NaOH将pH调到7.4,0.22 μm滤器过滤,分装后4 ℃保存。使用时按Solution 1∶Solution 2为1∶50的体积比配制成电转液。

表1 本研究所用引物
1.2.5 电转染哈萨克羊胎儿成纤维细胞 哈萨克羊胎儿成纤维细胞传代后培养于6 cm培养皿中,当细胞融合度为80%~90%时消化细胞,离心5 min 后用PBS清洗细胞,再离心5 min,尽量吸干净液体后用100 μL电转液悬浮细胞,每组细胞的细胞量约为1×106个,将 6 μg pCMV-AncBE4 max-P2A-GFP质粒和2 μg 连接好的pGL3-U6-sgRNA-PGK-puromycin质粒共转染哈萨克羊胎儿成纤维细胞,电转完成后将电转杯放入培养箱中静置10 min, 接种至6 cm培养皿,37 ℃、5% CO2饱和湿度条件下培养6 h后更换新鲜完全培养基。继续培养24 h后观察荧光表达效果,运用Image J计数视野范围内的细胞数,计算转染效率,同时加入嘌呤霉素进行药筛。
1.2.6 CruiserTM酶酶切检测及测序分析 药筛72 h后运用数控显微操作系统(Eppendorf Transfer-Man)收集200个左右带绿色荧光的阳性细胞,将收集到的细胞用20 μL细胞裂解液裂解后直接作为PCR模板,使用检测引物(表1)扩增目标DNA序列,PCR产物纯化后进行CruiserTM酶检测。反应体系如下:PCR纯化产物取500 ng,补加ddH2O至7 μL,98 ℃变性3 min后关掉干浴锅,待温度下降到40 ℃后取出加2 μL Curiser Buffer和1 μL CruiserTM酶,45 ℃孵育20 min后加入2 μL Stop Buffer,用2%的琼脂糖凝胶电泳分离检测,并将PCR纯化产物送测序检测。同时将PCR纯化产物TA克隆后进一步测序鉴定,反应体系如下:2× Rapid Ligation Buffer 5 μL,pGEM®-T Easy Vector 1 μL,PCR纯化产物2 μL,T4 DNA Ligase 1 μL,ddH2O 1 μL。放置于干浴锅中16 ℃恒温过夜连接,连接完成后转化送测序。
2 结 果
2.1 哈萨克羊胎儿成纤维细胞的培养
采用组织块贴壁法建立哈萨克羊胎儿成纤维细胞系。剪碎的胎儿皮肤组织在培养2 d后即可以明显的观察到细胞生长,培养4 d后在显微镜下可以观察到细胞生长晕,细胞以梭形为主,形态不均一,生长力旺盛,无污染,此时取出组织块(图2A)。培养7 d后传代,由于成纤维细胞贴壁较快,因此在传代2 h后换液,以纯化胎儿成纤维细胞。继续培养3 d 后再次传代,细胞形态均一,呈梭形,边缘清晰,生长力旺盛(图2B),证明成纤维细胞已纯化,成功获得哈萨克羊胎儿成纤维细胞。
2.2 构建MSTN-sgRNA载体并转染哈萨克羊胎儿成纤维细胞
测序结果显示,MSTN-U6-STOPsg-1和MSTN-U6-STOPsg-2质粒构建成功(图3)。
采用电转染将pCMV-AncBE4max-P2A-GFP质粒和MSTN-sgRNA质粒共转染哈萨克羊胎儿成纤维细胞,24 h后用荧光显微镜观察绿色荧光表达情况(图4)。使用Image J计算得出STOPsg-1和STOPsg-2的转染效率分别为45.3%、44.7%。
2.3 MSTN基因靶位点突变检测
CruiserTM酶能对不完全配对的双链DNA切割造成双链DNA断裂,其识别范围最小为1个碱基,最大可达上百个碱基的不配对序列。靶位点的PCR纯化产物经CruiserTM酶酶切,如果靶位点有突变,则STOPsg-1酶切后出现402和306 bp的片段,STOPsg-2会出现305和231 bp的片段。酶切结果显示,STOPsg-1和STOPsg-2的PCR产物均被切开(图5),说明STOPsg-1和STOPsg-2靶位点可能存在突变,需进一步测序验证。
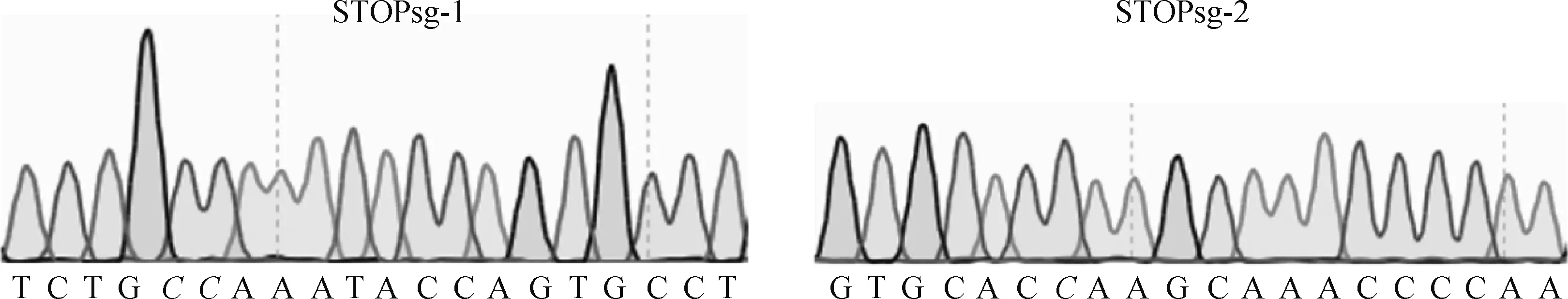
靶位点标为斜体The sgRNA target site is labeled in italic图3 sgRNA质粒测序分析Fig.3 Sequencing analysis of sgRNA plasmid

图4 电转染24 h的荧光表达情况Fig.4 Green fluorescence expression at 24 h after electrotransfection
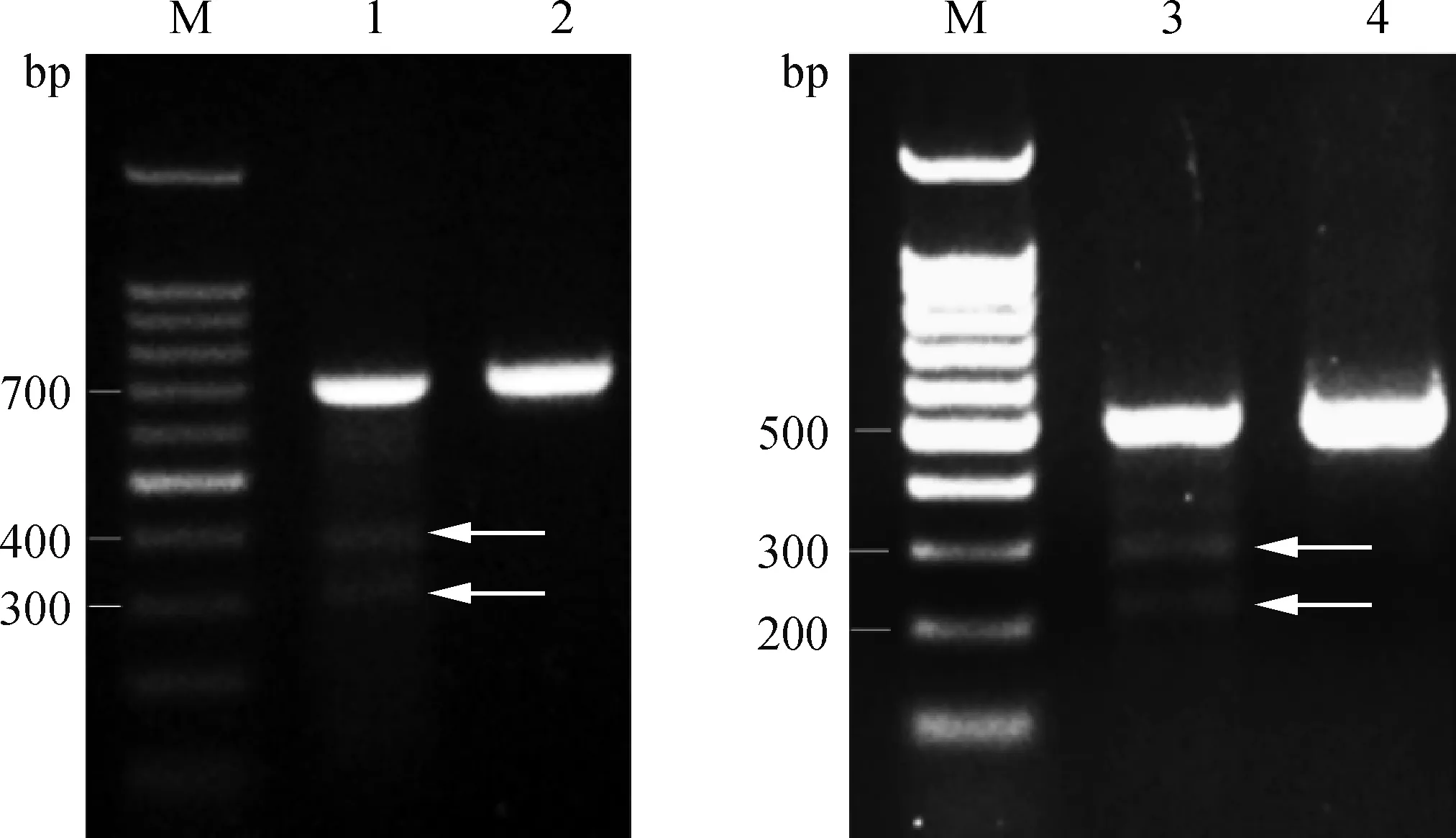
M. DNA相对分子质量标准; 1.STOPsg-1; 2. STOPsg-1阴性对照; 3.STOPsg-2; 4.STOPsg-2阴性对照; 箭头表示CruiserTM鉴定的条带M. 100 bp DNA ladder; 1.STOPsg-1; 2. STOPsg-1 negative control; 3.STOPsg-2; 4.STOPsg-2 negative control; The arrows indicate the bands identified by CruiserTM图5 CruiserTM酶鉴定结果Fig.5 Mutation detection by CruiserTM
2.4 MSTN基因靶位点测序结果分析
AncBE4 max可以使活性窗口内的C转换为T,活性窗口为sgRNA PAM位点远端数4~8位。STOPsg-1和STOPsg-2的PCR纯化产物送测序检测,结果显示,STOPsg-1和 STOPsg-2靶位点均有重叠峰(图6),表明STOPsg-1和STOPsg-2靶位点存在突变。然后对STOPsg-1和STOPsg-2的PCR纯化产物TA克隆后进一步测序验证,结果显示,STOPsg-1和STOPsg-2靶位点均发生C>T或G>A的突变,证明成功提前引入终止密码子,效率分别为26.7%(4/15)(图7A)和6.7%(1/15)(图7B)。同时,在靶位点附近观察到非目的突变(表2),未观察到C到A/G的非预期的突变和随机的indels。
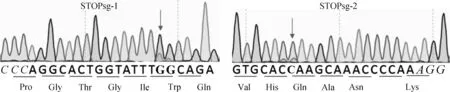
PAM区域标为斜体,靶位点标为加粗黑体Protospacer adjacent motif (PAM) region in italic, target sites in bold图6 靶位点测序峰图Fig.6 PCR product sequencing map of target sites
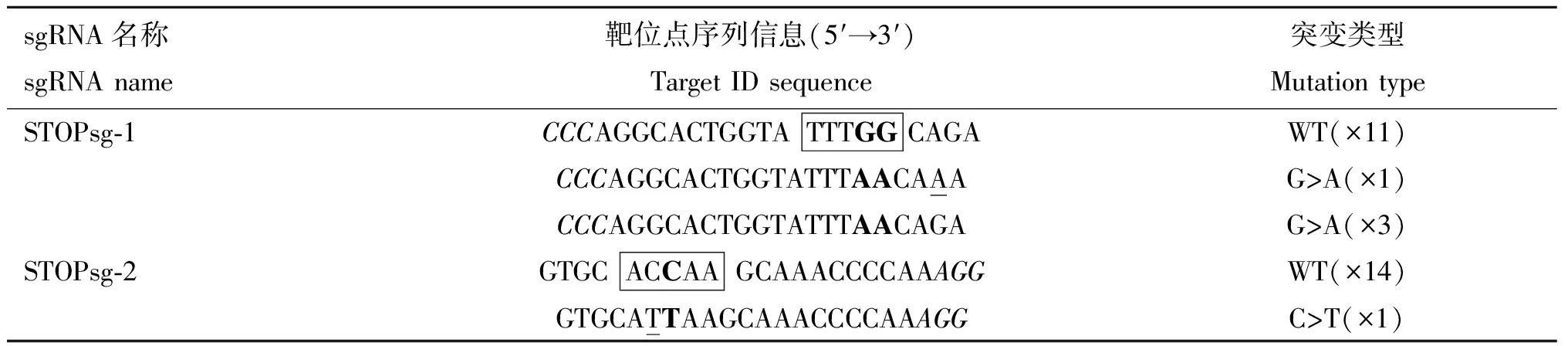
表2 MSTN基因TA克隆测序结果
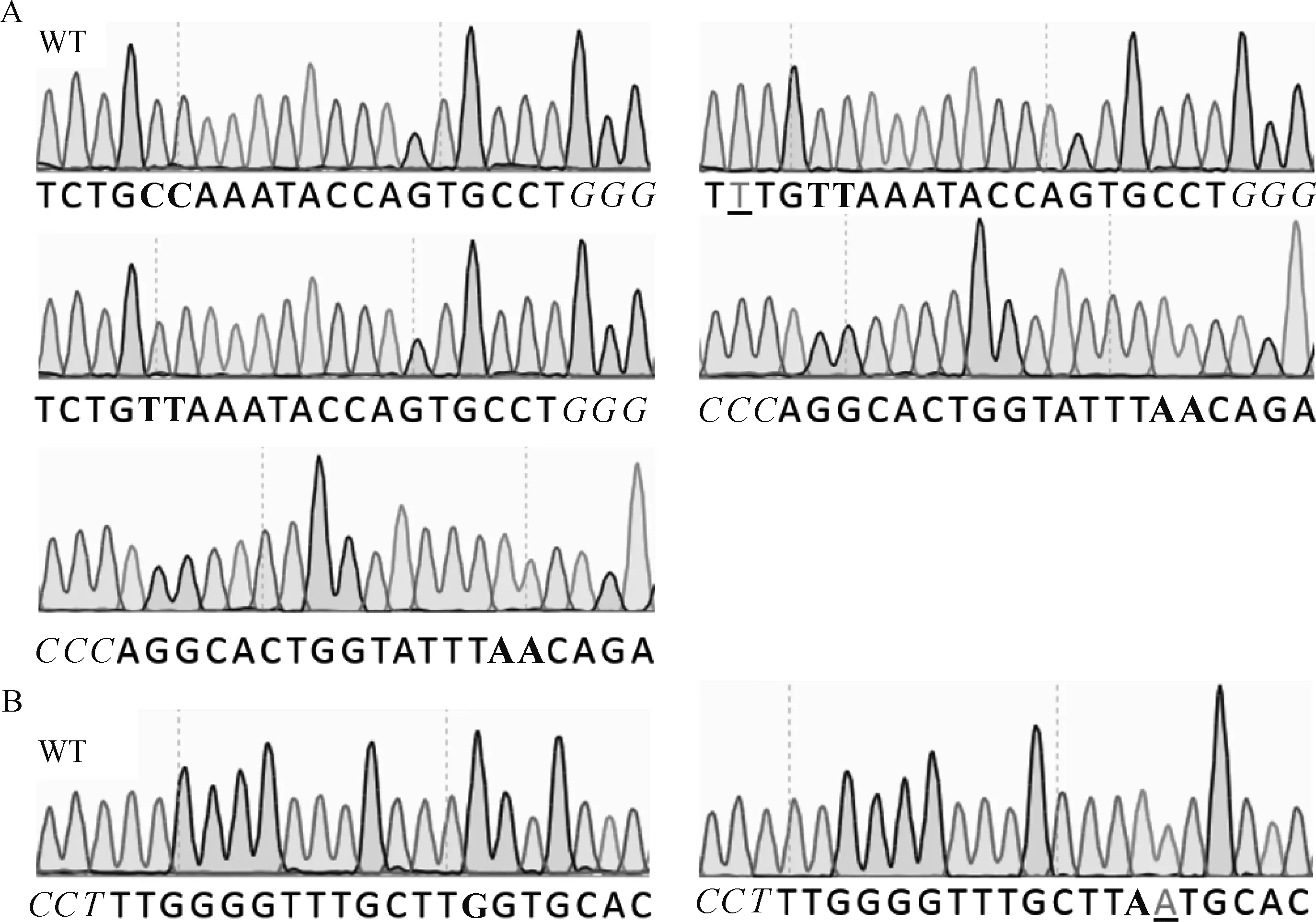
A.STOPsg-1; B.STOPsg-2。PAM区域标为斜体,靶位点标为加粗黑体,下划线表示靶点附近非目的突变A.STOPsg-1; B.STOPsg-2. Protospacer adjacent motif (PAM) region in italic, target sites in bold, non-target mutations near the target in underscore图7 TA克隆Sanger测序峰图Fig.7 Sanger sequencing peak map of the TA clone
3 讨 论
目前,大多数基因敲除动物是通过CRISPR/Cas9技术实现的,这需要形成DSB以使基因产生indels,从而达到敲除基因的目的。但是,并非所有indels都会导致基因沉默,有些甚至可能导致目标蛋白质具有新功能[25]。并且当CRISPR/Cas9靶向高拷贝数区域时,可能会导致过度的DNA损伤并降低适应性[26-27]。相比之下,BEs系统不需要引入DSB就能使基因沉默。本研究运用CBE(AncBE4 max)系统编辑哈萨克羊胎儿成纤维细胞MSTN基因,经Sanger 测序可知,成功筛选出在MSTN基因第二和第三外显子能提前引入终止密码子的2条sgRNAs,未发现indels。Liu等[28]运用BE3在兔MSTN基因提前引入终止密码子使其失活,获得的7只兔子中6只是纯合子突变,并且对潜在脱靶位点进行检测后未发现indels和脱靶突变。迄今为止,获得特定碱基突变动物的主流方法是CRISPR/Cas9系统与供体DNA结合,通过HDR途径实现的,但效果较差,并且会产生许多不需要的indel突变[29-30]。与之相比,BEs系统可以更简单有效地编辑目标碱基,不依赖细胞周期,不产生DSB或外源DNA模板导入,并且引入随机indels很低。Komor等[14]的研究表明,BE3编辑人类细胞时突变效率为37%,但indels频率很低,平均为1.1%,相反,供体DNA介导的HDR仅有0.5%的效率,但indels的百分比更高(4.3%)。
本研究使用AncBE4 max系统在哈萨克羊MSTN基因中提前引入终止密码子,以敲除MSTN基因,筛选出STOPsg-1和STOPsg-2两条可以实现有效编辑的sgRNA,编辑效率分别为26.7%和6.7%,略低于其他文献的报道[21,31]。这可能与sgRNA的设计有关,本研究设计的两个sgRNA靶核苷酸前面的核苷酸均为C,胞嘧啶核苷脱氨酶 APOBEC1偏好编辑靶核苷酸C的前面是T的序列,编辑效率遵从TC>CC≅AC>GC的规律[14]。本研究还观察到靶点附近出现非目的突变(表2),但未观察到C到A/G的非预期突变和随机的indel。出现C到A/G的非预期突变是因为C脱氨基后的U会被细胞碱基切除修复(BER)机制视为错误或破坏,然后由无处不在的尿嘧啶DNA糖基化酶(UDG)去除[32],UDG去除U会导致无碱基位点的形成,并随后通过易错的聚合酶进行修复,从而导致随机核苷酸掺入。另外,偶尔的链断裂可能是indel产生的原因[33],迄今为止,很少有研究报道CBE形成大量插入缺失[34-35]。靶点附近出现非目的突变可能是由于APOBEC1的高活性引起多个Cs脱氨基[36]。尽管当使用BEs来破坏启动子、剪接位点或者过早引入的终止密码子使基因沉默时,靶点附近非目的突变可能并不是特别重要,但会对探索基因内特定点突变产生影响。基于工程化人胞苷脱氨酶APOBEC3A(eA3A)所构建的CBE具有编辑特定胞苷的特性,能显著降低靶点附近的非目的突变[37]。AncBE4 max与工程化APOBEC3A结合是降低靶点附近非目的突变的有效措施。经不断改进,BEs在动植物品种改良、动物模型构建和疾病治疗等方面将展现出巨大的潜力。
4 结 论
本研究运用CBE(AncBE4 max)系统成功编辑了哈萨克羊胎儿成纤维细胞MSTN基因,筛选出能有效在第二和第三外显子提前引入终止密码子的2条sgRNAs,STOPsg-1靶位点编辑效率为26.7%,STOPsg-2靶位点的编辑效率为6.7%。为后期生产产肉率高并具有生物安全性的哈萨克羊奠定基础。
猜你喜欢
丝绸之路(2023年2期)2023-07-10 21:34:47
生物技术通报(2023年2期)2023-03-07 12:54:58
心声歌刊(2021年2期)2021-07-16 07:05:56
教学考试(高考生物)(2020年6期)2020-11-23 05:25:56
食品与生物技术学报(2020年8期)2020-01-06 08:00:56
科学24小时(2019年5期)2019-06-11 08:39:38
发明与创新(2019年9期)2019-03-26 02:22:48
卫拉特研究(2018年0期)2018-07-22 05:47:28
中国民族医药杂志(2016年9期)2016-05-09 07:51:52
中国药业(2014年21期)2014-05-26 08:56:45
