
HKT1在植物耐盐机制中的研究进展
2021-08-11 02:46张永兰解莉楠
生物技术通报 2021年6期
张永兰 解莉楠
(东北林业大学生命科学学院 东北盐碱植被恢复与重建教育部重点实验室,哈尔滨 150040)
土地盐碱化是作物产量降低的主要因素之一。由于气候变化和不合理灌溉,盐碱化土地的面积不断增加,土地盐碱问题日益严重[1]。在土壤积累的盐离子成分中,Na+对植物的影响最为突出。Na+破坏蛋白质和膜的稳定性,导致细胞内活性氧信号(reactive oxygen species,ROS)的产生,使植物细胞功能紊乱[2]。为了维持细胞中的Na+稳态,植物通过不同的Na+转运机制来保护自身免受Na+积累的损害,如将根中Na+排出到土壤(盐过敏途径,salt overly sensitive,SOS)[3-4];在地上部分将Na+运输到维管组织(高亲和性钾转运蛋白1,high-affinity potassium transporter1,HKT1)[5-6],以 及 将Na+隔离到液泡中(钠离子/氢质子交换蛋白,Na+/H+exchanger,NHX)[7-8]。
HKT1是最重要的Na+转运蛋白之一[9]。在许多物种中,HKT1通常在木质部薄壁细胞中表达[10]。近期研究表明,无论在单子叶植物还是双子叶植物的耐盐机制中,HKT1都有不可或缺的重要地位[11]。这使得HKT1的研究成为植物抗逆性工程的优先目标。因此,研究HKT1参与影响植物耐盐性的生理机制,为植物抗逆提供依据。
本文综述了HKT1转运蛋白在不同植物中的主要功能,以及在植物内调控表达过程的研究进展,讨论了HKT1蛋白在植物Na+长距离运输中起到的作用和相关模型,并对利用调控HKT1转运蛋白表达实现作物耐盐育种的可行性进行了展望。
1 HKT转运蛋白的结构与分类
自1994年在小麦(Triticum aestivum)中发现TaHKT2;1后,陆续从水稻、拟南芥等植物中发现了具有不同运输特性的HKT转运蛋白[9]。这些HKT蛋白都具有典型的TrkH保守结构域。HKT类转运蛋白包含8个跨膜域,每2个跨膜域和1个孔状P-loop组成1个跨膜基序(图1-A)[10]。起初HKT家族蛋白的命名规则十分混乱,2006年,国际间就HKT家族成员的命名方式达成了共识,以蛋白质第一孔域(first pore domain,PD1)中氨基酸的差异为主要特征,根据其运输特性,将HKT1蛋白分为两类(图1-B)[11]。第一类HKT转运蛋白的成员(HKT1)在PD1位置处是丝氨酸(Ser),同其他3个孔域(pore domain,PD)上的甘氨酸(Gly)残基形成丝氨酸-甘氨酸-甘氨酸-甘氨酸(Ser-Gly-Gly-Gly)的基序,丝氨酸的存在使HKT1转运蛋白对Na+的电导超过其他阳离子,决定了HKT1是Na+的转运体[12];第二类HKT转运蛋白的成员(HKT2)在PD1位置处是甘氨酸(Gly),从而形成甘氨酸-甘氨酸-甘氨酸-甘氨酸(Gly-Gly-Gly-Gly)的基序,因此,HKT2不仅可以是Na+/K+协同转运体,还可以是Na+或K+的单向转运体,而HKT2最终选择转运哪种离子则取决于这两种离子的外部浓度[13]。后续研究表明,将拟南芥中HKT1蛋白质第一孔域68位的丝氨酸突变为甘氨酸[14],则相应的蛋白功能也从单纯的Na+转运体变为Na+/K+协同转运体。
但也有例外,蓝桉树(Eucalyptus camaldulensis)的EcHKT1;2[15]、冰叶日中花(Mesembryantemum crystallinum)的McHKT1;1[16]、盐芥(Thellungiella salsuginea)的TsHKT1;2[17]等虽然在PD1中含有丝氨酸,但也能够转运K+,表明除了PD1位置上的丝氨酸和甘氨酸之外[14],还有其他位置上的氨基酸参与决定HKT蛋白是否有运输K+的功能。值得注意的是,这些PD1上丝氨酸/甘氨酸并不能决定离子选择性规则的情况,主要存在于盐生植物中 (表1)[18]。

表1 HKT类转运蛋白的分类Table 1 Classification of HKT transporters
2 HKT1的功能特点
维持细胞内离子的动态平衡是植物应对过量离子的重要能力[36]。植物通过降低细胞质中Na+含量,增加K+含量,可以使细胞质保持合适的钠钾比[1-2],从而避免细胞损伤和营养缺乏的情况产生[37]。减少细胞质中Na+含量的方法包括:限制Na+摄取[38]、增加Na+流出[39]和对细胞内的Na+进行区域化[40]。高亲和性钾转运蛋白(high-affinity potassium transporter,HKT)是一类具有离子转运特性的膜蛋白,其中HKT1能够特异性转运Na+,通过对Na+的长距离运输和分配,维持组织和细胞内的Na+/K+平衡[41]。
植物中的HKT1蛋白主要定位于根中柱木质部薄壁细胞(xylem parenchyma cells,XPC)的质膜[42]。HKT1在该区域中的表达可以使Na+从木质部中转运到木质部薄壁细胞,避免Na+被转运到地上部分,从而保证植物的光合作用不被破坏[43]。在单子叶植物和双子叶植物中,HKT1家族成员的数量是不同的,单子叶植物的 HKT1成员比双子叶多[11]。迄今为止,通过对单子叶植物中分离出的所有HKT1转运蛋白的研究发现,所有单子叶植物的HKT1蛋白都具有对Na+的选择性运输的共同特征[11]。但是不同植物间又有着不同的特点。接下来,将分别讨论不同植物中HKT1蛋白在离子运输以及植物耐盐性中的作用。
2.1 AtHKT1在拟南芥中的功能
AtHKT1;1是从拟南芥中分离出的唯一一个HKT1家族成员[20]。AtHKT1;1在非洲爪蟾卵母细胞中进行异源表达时,显示出单一的Na+选择性运输活性[20]。在植物中,首先考虑AtHKT1;1是否可以从外部培养基中摄取Na+。但是在拟南芥野生型和athkt1;1突变中,二者的根对于Na+的吸收并没有差异[44]。虽然AtHKT1;1在根和叶木质部与韧皮部均有表达,但在根尖中没有表达,这也不利于拟南芥从外部介质摄取Na+,说明在拟南芥根中,HKT1没有将Na+从外部介质转运到细胞内的功能[44]。
虽已知AtHKT1的离子选择性与具体表达位置,但是仍不能确定HKT1在植物应对盐胁迫中的具体机制[45]。随着对拟南芥HKT1蛋白的深入研究,研究者逐步提出几种关于HKT1蛋白作用的假设模型。
Rus等[9]通过对athkt1;1突变体进行分析 发现,AtHKT1;1的突变减少了幼苗中Na+的总量,从而提出了一种假设,即AtHKT1;1是根中Na+内流过程中起主要作用。而随后的研究表明,在athkt1;1突变植株中,根中Na+的内流并不低于野生型[46],于是先前的假设被推翻。通过对athkt1;1突变株进行盐处理后,与野生型相比,经过盐处理的突变体在韧皮部中积累的Na+更少[47]。拟南芥韧皮部组织附近的茎中也存在HKT1的微量表达。综合AtHKT1;1在韧皮部的特异性表达,还有野生型与突变体中韧皮部Na+含量的不同,Berthomieu等[47]提出AtHKT1;1将地上部分多余的Na+通过韧皮部向下运输,运回到根部,即所谓的“再循环”模型。
Sunarpi等[21]研究表明,与野生型植物相比,athkt1;1突变植株在茎和木质部中积累的Na+含量更高。通过采用抗体结合和GUS检测发现,AtHKT1;1主要在拟南芥木质部薄壁细胞(XPC)中表达[21]。基于此,作者提出了“外排”模型。即,拟南芥HKT1蛋白主要通过木质部薄壁细胞从根的木质部卸载Na+,避免过量的Na+通过蒸腾流运输到地上部分[21]。此外,基于单向22Na+示踪数据表明,拟南芥HKT1蛋白没有令Na+通过韧皮部的作用[48]。目前,由于以上两点原因,“再循环”模型受到了质疑。
在木质部中,验证了AtHKT1;1卸载Na+的功能[49]。并且木质部中Na+卸载量的增加,会增强植物的耐盐性。目前,虽然AtHKT1;1在根中的功能比较明晰,但其在地上部分的功能阐述却不甚明确,所以,“再循环”模型也有存在的可能。2种模型的本质都是Na+的运输,由于从木质部转运到木质部薄壁细胞的离子可以通过共塑扩散加载到韧皮部,联系“再循环”过程,就很有可能实现Na+在植物体内的转运(图2)。Van Zelm等[1]认为AtHKT1;1在拟南芥地上部分将多余的Na+装入韧皮部,而在根中,AtHKT1;1将Na+从木质部转运到木质部薄壁细胞[48],从而达到促进Na+从茎到根的下行运输,抑制根到茎的上行运输的目的。这也从侧面同时承认,在拟南芥中,“外排”模型和“再循环”模型可能同时存在。
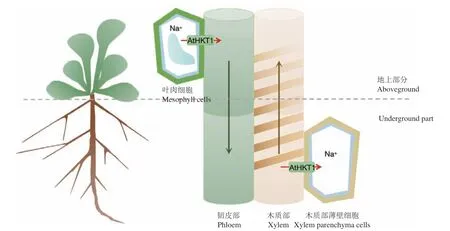
图2 HKT1在Na+长距离运输中的作用Fig. 2 The role of HKT1 in Na+ long-distance transportation
2.2 水稻中OsHKT1的类型及功能
在水稻中,与盐胁迫相关的数量性状位点SKC1(shoot K+concentrations)[23]参与维持茎中较高的K+浓度,SKC1对应的基因为OsHKT1;5(OsHKT8)。Horie等[50]研究表明OsHKT1;5在水稻中的作用与AtHKT1;1在拟南芥中的作用类似,均编码特异性Na+转运蛋白,在根中木质部薄壁细胞膜上表达[26],通过卸载木质部中Na+,限制其向地上部运输来降低地上部Na+含量[27],进而间接影响K+的含量,维持植物体内Na+/K+平衡。还有研究发现水稻OsHKT1;5除了在根中表达外,在叶鞘木质部薄壁细胞和成熟叶片韧皮部中也表达[24];22Na+放射性标记试验表明,OsHKT1;5还能够向韧皮部运输Na+,以降低Na+向幼嫩叶片的运输。
除OsHKT1;5外,OsHKT1;4在Na+运 输 过程中也起到重要作用(表1)。在盐处理条件下,OsHKT1;4在叶鞘中特异表达[22],尽管在水稻品系Pokkali和Nipponbare中存在3种不同的OsHKT1;4转录本,但只有全长转录本的数量才与叶鞘中Na+的浓度成反比[25]。表明在水稻的地上部分中,全长OsHKT1;4控制Na+从叶鞘到叶片的转移。
2.3 小麦中耐盐主效基因TaHKT1的分类及功能
在硬粒小麦(表1)(Triticum turgidum L. subsp. Durum)中,鉴定出2个QTL位点Nax1和Nax2,参与从木质部中排出Na+和降低叶片中Na+含量的过程。此外,含有基因组A、B和D的六倍体面包小麦(Triticum aestivum)比只含有基因组A和B的四倍体硬粒小麦更耐盐[51]。分析发现,D基因组中的Kna1位点能够使面包小麦在盐胁迫条件下,维持相对较低的Na+/K+比例[52]。QTL位点Nax1、Nax2和Kna1均参与将Na+从木质部转运到叶鞘中的过程。通过精细定位,Nax1和Nax2被鉴定为TaHKT1;4,Kna1被鉴定为TaHKT1;5[28]。由于2个Nax基因都起源于与硬粒小麦杂交的一粒小麦(Triticum monococcum),因此,分别将Nax1和Nax2命名为TmHKT1;4-A2和TmHKT1;5-A[28]。这两个基因显然具有相似的功能:降低小麦叶片中的Na+含量[30],与拟南芥中的AtHKT1和水稻中的OsHKT1;5类似,都能减少各自植物体叶片中的Na+。此外,含有TmHKT1;5-A的硬粒小麦的近等基因系(near isogenic lines,NIL),能够在盐胁迫条件下具有更高的籽粒产量。以上结果表明,HKT1基因在小麦耐盐性中具有至关重要的作用[53]。
2.4 其他HKT1家族成员的功能
大麦(表1)(Hordeum vulgare L.)是强 耐盐性的禾谷类作物,广泛应用于作物耐盐性研究。通过对2 671份大麦材料进行GWAS分析,发现HvHKT1;5的标记SNPs与大麦耐盐性高度关联。HvHKT1;5编码一种特异性转运Na+的蛋白,定位于根中柱细胞质膜,转运Na+的功能受外界K+浓度影响[35]。利用RNAi技术敲除HvHKT1;5的表达之后,大麦的生长呈现出与预期相反的耐盐性[54]。这表明,与常见的HKT1蛋白研究结果相反,敲除HvHKT1;5导致木质部组织中Na+浓度降低,Na+从根到地上部分的转移减少,与野生型植株相比,敲除HvHKT1;5的转基因植株的耐盐性增加。以上均表明HvHKT1;5参与了Na+被转运到木质部内的过程,这一发现与已知的AtHKT1在木质部卸载Na+的功能相反。
在番茄(表1)(Solanum lycopersicum)中,检测到2个密切相关的HKT1基因[32]。对这两个基因进行异源表达分析发现,在低K+介质中,SlHKT1;1和SlHKT1;2的表达都不能补充酵母突变体的生长;与此同时,表达SlHKT1;1的细胞能够耗尽外部Na+,这表示SlHKT1;1是Na+选择性转运蛋白[55];而在表达SlHKT1;2的酵母细胞中,没有检测到任何Na+或者K+的转运活性[56]。随后进行的基因表达定位分析表明,在番茄的根、茎、叶、花、果实中普遍存在SlHKT1;1和SlHKT1;2的表达[32]。RNAi干涉降低SlHKT1;2的表达,则改变植物叶片中的钠钾比,并降低植株耐盐性[57]。这些结果表明,SlHKT1;2在维持番茄的钠平衡和提高耐盐性方面起着更为显著的作用。
从鼠尾栗(表1)(Sporobolus virginicus)中分离鉴定出来的HKT1蛋白(SvHKT1;1)异源表达时,可以调控Na+的进出,但是不能使钾离子转运蛋白缺失的酵母突变体恢复生长[19]。而且SvHKT1;1的表达也不能恢复拟南芥athkt1;1突变体的生长,这表明SvHKT1;1和典型的HKT1转运蛋白的功能不相同。拟南芥中表达SvHKT1;1后,经过高浓度盐处理(500 mmol/L NaCl)[19],转基因拟南芥的SvHKT1;1在地上部分的表达量比根中的表达量低,这种地上部分表达量比根中表达量低的情况,在低浓度盐胁迫条件下并没有出现。在经历盐胁迫后,相比于野生型,转基因拟南芥的地上部分的Na+含量升高,耐盐性降低[19];而在盐胁迫的早期,根中的Na+的吸收率增高。这表示SvHKT1;1的表达增强了根细胞的Na+吸收,也提高了Na+向地上部分的的转运,而这些过程降低了耐盐性[19]。以上证据均表明,SvHKT1;1和其他植物中的HKT1作用不一致。在转基因拟南芥中,SvHKT1;1反而降低了植株的耐盐性[19]。
在不同植物中,AtHKT1;1的同源基因仍行使转运Na+的作用,不同的HKT1蛋白负责的Na+转运的方向不同,水稻OsHKT1;5和AtHKT1;1的功能基本一致,都是将Na+从木质部中转运出来,从而提高植株的耐盐性。但是在大麦中,HvHKT1;5介导的Na+运输方向则相反,能够促进Na+向木质部中转运,这也导致大麦HvHKT1;5的表达会降低植株的耐盐性[35]。同样,在转SvHKT1;1[19](来源于鼠尾栗)拟南芥中,SvHKT1;1的存在反而降低了植株的耐盐性。因此,HKT1蛋白在作物中的转运Na+的方向还有待进一步确定。
3 HKT1的调控机制
HKT1参与植物细胞内钠钾离子平衡的调节,但是关于HKT1基因表达调控过程和蛋白作用机制的研究还不完善。目前主要从第二信使、植物激素调控HKT1基因表达两方面进行研究。
3.1 第二信使调控HKT1的表达
利用缺乏解毒酶(detoxification enzymes)的拟南芥突变体[58]、缺乏参与产生活性氧(reactive oxygen species,ROS)酶的拟南芥突变体[59],以及参与产生ROS的酶抑制剂[60]处理的野生型拟南芥的几项研究表明,植物体内ROS积累量的高低和耐盐性的强弱有明显关系。拟南芥突变体atrbohF(Arabidopsis thaliana respiratory burst oxidase protein F,拟南芥呼吸爆发氧化酶蛋白F)的研究表明,盐胁迫下,AtRbohF增加根维管束中ROS水平,导致木质部中Na+含量的减少,从而减少Na+运输到地上部分的浓度[61]。因为AtHKT1;1参与Na+从木质部卸载的过程,科研人员提出一个假设来解释AtRbohF对木质部Na+水平的影响,即AtRbohF产生的ROS调控AtHKT1;1表达或活化AtHKT1蛋白(图3)。此外,盐胁迫增强了AtRbohF在根中维管组织的表达。受增加的ROS影响,钾离子通道SKOR在木薄壁细胞中表达活性增强[59],这也说明ROS水平对于转运蛋白的活性是至关重要的。
在钙离子调控Na+进入细胞的过程中,钙调蛋白(calmodulin,CAM)[62]以及钙调蛋白依赖的转录激活子(CaM-binding transcription activator,CAMTA)家族有着不可或缺的作用(图3)。在拟南芥中,CAMTA6突变个体呈现出更强的耐盐性[63],而且植物中积累的Na+含量变少,随后进行的生物信息学研究发现,AtHKT1;1的启动子内部有潜在的CAMTA结合位点(CACGTGTC基序),同时AtHKT1和CAMTA6在盐处理后表达水平变化趋势一致,这表示CAMTA6与AtHKT1的表达调控密切相关[64]。除此之外,经盐处理后的野生型中有1 020个上调和1 467个下调的盐响应基因。其中,638个上调基因和1 242个下调基因被归类为CAMTA6依赖基因。在大麦中,HvHKT1;1和HvHKT1;5的启动子区域都有HvCAMTA4的顺式反应元件[63]。HvHKT1;1是Na+转运蛋白,表达量与大麦的耐盐性正相关,但是HvHKT1;5是大麦中耐盐性的负调控因子。Shen等[65]研究表明HvCAM1与HvCAMTA4有互作关系,且HvCAM1、HvHKT1;1和HvHKT1;5都在维管束中表达,敲除HvCAM1的突变体植株中,HvHKT1;1表达量上升,而HvHKT1;5表达水平下降。与野生型相比,过表达HvCAM1的植株呈现出盐敏感的性状。这表明大麦中的钙调蛋白HvCAM1以及钙调蛋白依赖的转录激活因子HvCAMTA4、HvCAMTA6可以直接或间接地调节HvHKT1的表达,从而影响植物耐盐性。
3.2 细胞分裂素调控HKT1的表达
众所周知,盐胁迫改变了植物中的细胞分裂素水平,而细胞分裂素又在响应盐胁迫的过程中起着作用。所有细胞分裂素受体和几种拟南芥A型反应调节因子的表达都会受到盐处理的影响[66],细胞分裂素受体基因功能的丧失使植物耐盐性提高。在拟南芥B型调节因子(Arabidopsis type-B regulator)双突变体atarr1-3arr1-12和野生型植株之间的比较中,双突变体植株对细胞分裂素不敏感[67],与未处理对照组相比,外用细胞分裂素导致植株中Na+的积累较高,但是野生型植株地上部分中的Na+增加了46%;atarr1-12突变植物的地上部分的Na+积累仅上升21%。表明突变后,植物耐盐性提高,细胞分裂素在在植物Na+积累过程中起作用[67]。与野生型相比,atarr1-12突变体的根中AtHKT1;1表达高达6.2倍,但茎中无明显变化,这表示在拟南芥根部,转录因子ARR1-3和ARR1-12调节AtHKT1;1的 表达[21]。
异戊烯基转移酶(isopentenyl-transferases,IPT)是植物细胞分裂素从头合成的关键酶。在拟南芥异戊烯基转移酶缺陷突变体ipt1,3,5,7中,与野生型相比,AtHKT1;1表达量提高。在根维管组织中,AtHKT1;1和ARR1-3和ARR1-12的表达模式一致[67],进一步验证了细胞分裂素信号控制AtHKT1;1的表达[68]。此外,外部应用细胞分裂素处理后4 h,野生型植物中AtHKT1的表达量减少87%;atarr1-12突变体中AtHKT1;1的表达量减少21%[67],这也证实了细胞分裂素在调节AtHKT1;1表达中的作用(图3)。虽然这些结果清楚地证明了细胞分裂素对AtHKT1;1的表达的影响,但这种调节的分子基础尚不清楚。此外,通过对促进植物根生长细菌Bacillus subtillis菌株GB03的试验表明,该菌株释放的挥发物诱导了AtHKT1;1在根和茎中的表达变化[69]。在GB03所产生的挥发物中,2,3-丁二醇是在盐胁迫下刺激生长的主要效应物。后来的一项研究表明,拟南芥ein2(cytokinin/ ethyleneinsensitive,细胞分裂素/乙烯不敏感)和cre1(cytokinin receptor-deficient,细胞分裂素受体缺乏)突变体对GB03产生的挥发物没有反应,证实了2,3-丁二醇对AtHKT1;1表达的影响是通过细胞分裂素信号实现的[69]。但GB03诱导根AtHKT1;1表达是否由ARR1-3和ARR1-12介导的,目前尚不清楚。
3.3 其他调控方式
近期在拟南芥的研究中发现,蛋白磷酸酶G家族成员PP2C49参与调控盐胁迫下Na+在根和地上部分的分配(图3)[70],而且,在拟南芥根中,PP2C49通过直接抑制拟南芥HKT1蛋白的Na+转运活性,降低根中Na+的积累。PP2C49定位在细胞质和细胞核中,在根中的维管组织中高度表达,通过对PP2C49缺失和过表达的突变体的研究,发现PP2C49对植株的耐盐性进行负调控。离子含量和嫁接试验表明pp2c49突变体减少了地上部分和木质部中Na+离子含量[70]。而pp2c49突变体的耐盐性表型和Na+分布情况与AtHKT1;1中的相反,通过对PP2C49双突变体和过表达植株进行酵母双杂交、BiFC以及LUC试验,证明AtHKT1;1是PP2C49介导盐胁迫响应的下游基因,并且两者之间有物理互作。

图3 植物细胞离子响应策略以及HKT1表达调控途径Fig. 3 Plant cell ion response strategy and HKT1 expression regulation pathway
Wang等[71]研究表明,盐处理后,DNA甲基化识别酶OsSUVH7结合OsHKT1;5启动子区域甲基化的MTTE转座子上,募集分子伴侣调控蛋白OsBAG4和转录因子OsMYB106,形成稳定的转录调控复合体,激活OsHKT1;5的表达。该过程的发现为HKT1的调控启发了新思路,即盐胁迫激活表观调控通路,从而激活HKT1的表达。
4 展望
随着世界人口的增长,粮食安全问题日益凸显。由于人类活动范围的扩展,土地盐碱化问题不断加重[72],这一情况给农作物的产量带来严峻的考验,并且加重生态安全的威胁。为此,深入剖析植物耐盐性机理,以期获得高耐盐性植物资源,从而应对未来粮食安全的挑战。
HKT1作为重要的Na+转运蛋白,其离子运输作用明确,因此,对植物中各种HKT1转运蛋白的转基因研究,将提供可用于耐盐作物育种工程的重要信息[73]。例如,Moller等[38]的研究表明,在植物的根部过表达相应的HKT1蛋白,将会提高该植物的耐盐性。这是由于AtHKT1;1、SlHKT1;1、OsHKT1;4和TaHKT1;5是高度保守的同源基因,并且在植物中具有相同的功能[19,54,56]。因此,根中过表达相应的HKT1蛋白可能在耐盐作物育种工程中发挥重要作用。此外,将一个HKT1等位基因从一个相对原始的小麦品种转化到硬粒小麦[74],显著改善了硬粒小麦地上部分的Na+排出情况和田间耐盐性,提高了在盐胁迫下的籽粒产量。
在除了谷物之外的其他物种中,通过调节HKT1表达来提高耐盐性也是一种很有希望的方法。但是,由于HKT1本身在同一作物的不同品种中具有比较多的自然突变类型[75]。所以,HKT1的表达的效果是否对耐盐性有益,取决于植物的遗传背景和年龄,例如,在番茄品种Solanum lycopersicum和野生番茄品种Solanum pennellii中,野生番茄品种Solanum pennellii的耐盐性较高,可能是由于其HKT1蛋白对Na+的亲合性较低[56]。另外,从鼠尾栗中分离出来的SvHKT1;1转入拟南芥中,反而降低了拟南芥的耐盐性[19];降低大麦中HvHKT1;5的表达,会提高大麦的耐盐性[54]。因此,如果想要得到具备更高耐盐性的品种,则需要根据植物中HKT1不同的作用,针对的进行更精确的调控。
尽管在育种工程中,通过调节HKT1表达而成功改良作物耐盐性的例子并不多,但是不妨把眼界放的宽些,不仅仅聚焦在HKT1这一种离子转运蛋白上,可以发现包括番茄在内的物种中[40],过表达NHX1蛋白也可以增强相应植物的耐盐性。虽然在作物中关于HKT1表达调控过程和蛋白作用机制的研究还不甚完善,但这是进一步探索的一个有趣方向。鉴于HKT蛋白的功能与植物耐盐性密切相关,因此,在作物耐盐育种过程中,对与HKT1功能相似的离子转运蛋白,进行相关的生理作用和调控过程的研究,将会为培育耐盐作物新品种提供新思路[75-76]。
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
植物研究(2021年2期)2021-02-26
河南农业科学(2020年2期)2020-03-11
种子(2019年6期)2019-07-13
浙江农业科学(2017年8期)2017-08-30
天津农业科学(2015年11期)2015-12-03
安徽农学通报(2014年7期)2014-04-29
中国糖料(2013年4期)2013-03-02
