
感染期和休眠期结核分枝杆菌脂肪酸来源和利用差异的研究进展
2021-08-10 11:36刘立英秦守涛孙江洋李健明曾范利
中国兽医学报 2021年7期
周 颖,刘立英,秦守涛,丛 薇,李 东,孙江洋,时 坤,李健明,宗 颖,赵 丹,曾范利*,杜 锐*
(1.吉林农业大学 动物科学技术学院,吉林 长春 130118;2.辽宁农业职业技术学院,辽宁 熊岳 115009;3.吉林市动物疫病预防控制中心,吉林 吉林 132001;4.吉林省永吉县动物疫病预防控制中心,吉林 永吉 132100;5.吉林农业大学 中药材学院,吉林 长春 130118;6.吉林省梅花鹿药用资源利用关键技术研究室,吉林 长春 130118;7.吉林农业大学 农业质量标准与检测技术研究中心,吉林 长春 130118)
结核病(tuberculosis)是由结核分枝杆菌(Mycobacteriumtuberculosis)引起的一种慢性的、危害严重的人畜共患病,不仅对人和动物的健康造成危害,还会引起巨大的经济损失。2018 年,全球范围内据估约有1 000万(范围为900万~1 110万)结核病新发病例,各国的结核病负担差异较大,从每年每10万人发病例数低于5例到超过500例,而全球平均约为130例[1]。尽管全球努力解决结核病的发病率和传播问题,但每年仍约有1 000万人被诊断患有此病,导致约200万人死亡。我国是全球22个结核病高负担国家之一,同时也是全球27个耐多药结核病流行严重的国家之一。目前,结核病的治疗方法主要是依据具体病情采取2种以上药物的联合用药,短期在6~9个月,最终将活动性结核转化成非活动性结核病,痰菌呈阴性,结核菌受到药物控制进入休眠期[2]。结核分枝杆菌具有分枝杆菌属的细胞壁脂质含量较高的特性。细胞内病原体的新陈代谢与它们的宿主细胞密切相关,特别是对于结核分枝杆菌,脂质恰好是这种相互作用的主要介质,这些化合物同时作为病原体的营养源和宿主免疫反应的调节剂。有研究发现,结核分枝杆菌的脂肪酸,是使其在宿主体内幸存下来的关键所在[3]。现就不同时期结核分枝杆菌脂肪酸合成利用差异的研究进展进行综述。
脂肪酸(fatty acid)是指一端含有1个羧基的长的脂肪族碳氢链的有机物,是中性脂肪、磷脂和糖脂的主要成分。脂肪酸组成甘油磷脂的酰基链,甘油磷脂是细菌膜的主要和必不可少的组成部分[4-5]。细菌使用Ⅱ型脂肪酸合成系统合成脂肪酸,其中脂肪酸是使用离散的单功能酶从乙酰辅酶A前体通过2个碳原子的延伸来合成的[6]。脂肪酸在氧充足的情况下,可分解为CO2和H2O,释放大量能量,因此脂肪酸是主要能量来源之一。结核分枝杆菌主要位于巨噬细胞吞噬体内以及发生肉芽肿病变的各种组织和区域的细胞外[7-8],这些细胞和组织富含脂肪酸和胆固醇[8]。
1 感染期脂肪酸的合成和利用
1.1 感染期的结核分枝杆菌结核分枝杆菌进入宿主体内后,通过多种感染机制在宿主体内扩散,其中之一就是通过抗原递呈细胞递呈来感染巨噬细胞。在感染期间,结核分枝杆菌在巨噬细胞内大量复制,激活相关免疫应答反应,从而杀死结核分枝杆菌。但结核分枝杆菌内存在的一些毒力因子能够有效抑制巨噬细胞凋亡,抑制巨噬细胞的免疫应答,巨噬细胞无法将结核分枝杆菌抗原提呈给T细胞,阻止效应T细胞的激活[9],促进胞内结核菌的存活或持留[10]。
在感染期,结核分枝杆菌通过相关毒力因子促进巨噬细胞的凋亡,使结核菌更容易扩散和传播,造成更严重的感染[11],如通过肉芽肿的扩展和传播来增强肉芽肿的形成;当细胞内结核分枝杆菌负荷达到阈值时,巨噬细胞发生凋亡,将结核菌递呈给未感染的巨噬细胞,形成新的肉芽肿。结核分枝杆菌通过促进巨噬细胞死亡和被未感染的巨噬细胞吞噬[12]的途径,在感染期初期间在宿主体内大量扩散,并形成有利于其的生存环境。
1.2 脂肪酸在感染期的来源结核分枝杆菌通过所处环境获得脂肪酸,如通过水解宿主细胞膜脂质从宿主细胞获得脂肪酸[13]。同时,结核分枝杆菌感染巨噬细胞,感染后形成的脂质体与吞噬小体迅速结合并融合,为细菌提供了丰富的脂质来源。稳定的脂肪酸合成来源,也为脂肪酸的利用提供基础。
1.3 感染期脂肪酸的利用
1.3.1结核分枝杆菌利用脂肪酸供能 研究表明,结核分枝杆菌在感染期间主要使用脂肪酸而不是碳水化合物作为碳底物[3]。生长在小鼠肺中的结核分枝杆菌的呼吸受到脂肪酸的强烈刺激,但对碳水化合物没有反应。多条证据表明,宿主脂质(脂肪酸和胆固醇)是感染期间结核分枝杆菌的关键碳源[14]。在20世纪50年代,确定在哺乳动物组织中繁殖的结核分枝杆菌优先代谢脂肪酸[15],此后的大量研究反复证明,脂质代谢促进结核分枝杆菌在感染期间的存活[16-18]。
在感染期间,结核分枝杆菌可以通过水解宿主细胞膜脂质从宿主细胞获得脂肪酸[13],再通过β-氧化产生能量。乙醛酸循环作为结核分枝杆菌产生能量的重要途径,结核分枝杆菌通过β-氧化,将脂肪酸降解为乙酰辅酶A单位,如果糖酵解底物不存在或稀少,则为不完整的乙醛酸循环。
1.3.2脂肪酸与结核菌毒力和细胞壁相关性 结核分枝杆菌利用脂肪酸产生所需代谢中间产物,其与结核菌的毒力相关。结核分枝杆菌利用脂肪酸产生毒力所需的重要生物合成前体、聚酮类脂(PDIM)、聚酰化海藻糖(PATs)、磺化脂类(SL)和真菌酸。脂肪酸合成酰基AMP引物,通过聚酮合成酶反复修饰酰基链以最终产生聚酮类脂,聚酰化海藻糖和磺化脂类[19]。研究表明,富含脂质的分枝杆菌细胞壁由海藻糖二醇酸组成,可诱导肿瘤坏死因子-α的反应,从而引起肉芽肿性炎症反应[20]。诱导过量的肿瘤坏死因子-α反应是结核分枝杆菌毒力的另一种机制,有利于其在宿主中的存在而不被检测到。同时,细胞膜最外层中PDIM的存在可保护结核分枝杆菌免受巨噬细胞产生的活性氮中间体的杀灭作用;另外,由于细胞膜最外层中的大量PDIM影响免疫活性成分的表面暴露,导致小鼠巨噬细胞和树突状细胞分泌促炎细胞因子IL-6和肿瘤坏死因子-α的下调[21]。
脂肪酸也是结核分枝杆菌细胞壁重要成分,并参与合成结核分枝杆菌细胞壁。结核分枝杆菌的细胞壁拥有高含量的分枝菌酸、脂类和多糖。分枝菌酸(mycolic acid,MA)是具有α-烷基-β-羟基的C60-90的长链脂肪酸,是分枝杆菌属细胞壁主要的疏水性脂肪酸,占细胞壁干重的40%~60%[22]。MA在结核分枝杆菌的菌落形态、细胞壁的通透性、抗生素的敏感性、存活能力、致病性作用以及抵御宿主的免疫反应中扮演着重要的角色[23-26]。其中,酮-分枝菌酸和甲氧基分枝菌酸的有无可以明显改变结核分枝杆菌细胞壁的通透性,若细胞壁的通透性降低,会导致结核分枝杆菌的寿命和毒力都降低。同时,C16-26脂肪酸通过专门的脂肪酸合成酶Ⅱ复合物(FASⅡ)产生霉菌酸[27]。霉菌酸即分枝菌酸是长链α-烷基-β-羟基脂肪酸,负责细胞壁的流动性以及其渗透性[28],并在很大程度上被认为是结核分枝杆菌对脱水和化学处理的存在抵抗力的原因之一。
1.3.3结核菌利用脂肪酸产生磷脂和甘油三酯 结核分枝杆菌将脂肪酸同化到细胞膜磷脂中或转化为甘油三酯。磷脂是维持细胞质膜完整性所必需的,结核分枝杆菌内的大多数短链不饱和脂肪酸位于磷脂中,而饱和脂肪酸作为霉菌酸的生物合成前体,或复杂脂质的取代基,被用于细胞包膜的外层。同时,当感染巨噬细胞并在其中生长繁殖时,结核分枝杆菌可以代谢整合脂肪酸和利用胆固醇,其中结核分枝杆菌利用脂肪酸来平衡胆固醇衍生的代谢中间体,使其加入细菌细胞壁脂质的生物合成[29]。脂肪酸转化甘油三酯后则作为一种碳源能量储存,当营养物质受到限制时可被分解供能[30]。同时,脂肪酸转化为甘油三酯促进结核分枝杆菌的药物耐受性[31],这是一种结核分枝杆菌在体内感染期间通过免疫压力进一步增强的表型耐受[32]。
2 休眠期的脂肪酸合成和利用
2.1 休眠期的结核分枝杆菌目前,结核病的治疗药物以化学药物为主,主要是将活动性结核转化成非活动性结核病,结核菌受到药物控制进入休眠期,在休眠期内,结核菌不会复制,但可以保持活性并内化宿主脂质。但是,由于结核分枝杆菌为胞内寄生菌,可以长期在细胞内处于休眠状态,直到宿主免疫系统被削弱,然后重新激活。结核病的病理原因复杂,很大程度上归因于结核菌为建立初级感染而采取的免疫逃避策略,它在宿主中持续存在,并在有利的条件下重新激活致病性[20]。
根据定义,结核分枝杆菌的休眠期即潜伏期是指在宿主内持续存活多年而不引起临床疾病或在结核菌素试验中显示反应性的生物体的临床病症。在休眠期间,结核分枝杆菌局限于由宿主免疫细胞聚集组成的肉芽肿中,肉芽肿防止结核菌扩散,此时结核菌的特征是体外生长缓慢,代谢途径减少,染色特征改变,不能在固体培养基上培养,以及对抗结核病治疗药物的抗性。通过这种休眠状态逃脱宿主的免疫反应从而在宿主细胞中存活,并在有利条件下重新恢复活性。
休眠期的结核分枝杆菌处于脂质负载的巨噬细胞内,结核菌的复制受到严重抑制。有研究表明,在多种压力下,转录/翻译装置中总体活性的降低。例如,rhlE(ATP依赖性RNA解旋酶同源物)被阻遏,表明转录机制中活性降低。ATP合酶亚基的基因表达被抑制,表明细胞中ATP生成的主要关闭;同时,有氧呼吸被显着抑制。此外,厌氧呼吸所需的基因(frdA,narG/H/X,nirA)持续表达(图1)[33]。
在缺氧刺激下,脂质负载的巨噬细胞内的结核菌积累中性脂质,失去耐酸性,同时产生抗生素耐药性。抗生素耐药性的发展被称为“表型耐药”或“耐药”,通过细菌对抗生素的通透性下降,抗生素的外排,抗生素的修饰以及多个基于靶标的耐药性突变的逐步积累等产生的[34]。当结核分枝杆菌进入休眠非复制状态的能力,具有非常低的代谢活性和对现有抗结核药物的表型耐药性[35-36]。
2.2 脂肪酸在休眠期的来源休眠期的结核杆菌位于肉芽肿组织包裹中,环境中脂肪酸和胆固醇富集,并处于缺氧环境下。有证据表明,脂肪酸是病原体在休眠期长期存活所使用的能源[36-37],因此,在通过模拟缺氧和不治疗、应激等条件下,研究细菌内脂肪酸来源。通过放射性标记宿主脂肪酸,观察到其从宿主细胞转移到结核分枝杆菌中,并合成甘油三酯。脂质负载巨噬细胞内结核分枝杆菌内积累的甘油三酯主要是通过结核分枝杆菌tgs1基因产物将宿主脂源性脂肪酸重新酯化成甘油三酯合成的[38]。同时,在多重压力下,编码乙醛酸循环酶的基因,例如异柠檬酸裂合酶(ICL或aceA/Rv0467)和柠檬酸合酶(gltA1/Rv1131c)表达显著增加(图1)[33],表明脂肪酸合成增加,脂肪酸为休眠期结核菌的重要能量来源。
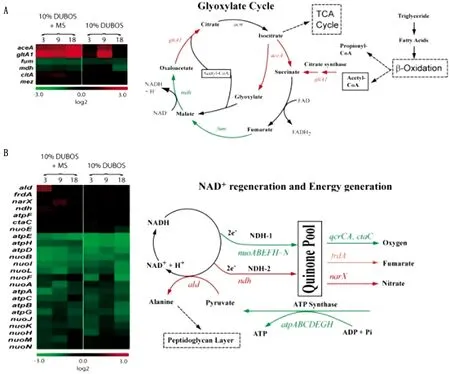
图1 微阵列分析显示乙醛酸循环、NAD再生和能量代谢相关基因表达的变化[33]
2.3 休眠期脂肪酸的利用在休眠期,结核分枝杆菌将脂肪酸作为重要碳源,将宿主来源的脂肪酸用于3个主要方面。
2.3.1支持中枢新陈代谢 与处于感染期的情况类似,休眠状态下的结核分枝杆菌也利用脂肪酸作为其重要碳源之一。由于处在低氧高脂质下,造成了肉芽肿内部其他营养物质缺乏,利用脂肪酸维持中枢系统的新陈代谢更加重要。处于休眠状态下的结核分枝杆菌基本不进行复制,其生长水平及分裂状态都非常缓慢,使休眠期的结核分枝杆菌的代谢出现一定程度的改变。同时,以脂肪酸为基底之一合成的甘油三酯作为休眠期结核分枝杆菌生长的能量来源[39]。同时,胆固醇也是菌体的重要能量源,但胆固醇代谢会对菌体代谢造成很大负担,结核分枝杆菌利用增加脂肪酸的含量来作为缓冲,利用脂肪酸和胆固醇共同代谢来确保菌体代谢平衡[40]。
2.3.2作为酰基引物产生与毒力相关的次生代谢物 与处于感染期的结核分枝杆菌相似,休眠期的病原体利用脂肪酸产生毒力所需的重要生物前体。例如脂质小体,是一种普通培养基中通常存在的脂肪物质,含有多聚β-羟基烷酸酯、甘油三酯和蜡酯。脂质小体更常见于结核分枝杆菌的休眠期,这也是休眠期结核菌表达脂质小体被认为是导致表型耐药的原因[41]。
2.3.3支持细胞膜完整性和创建碳储存分子[42]与感染期相似,脂肪酸通过降解、延伸和与其他物质合成结核分枝杆菌所必须的成分,维持细胞膜完整性。同时,转化为甘油三酯,作为能量储存。
在感染期,结核分枝杆菌利用脂肪酸作为碳源进行分裂增殖,侵染更多的巨噬细胞,同时也是细胞毒力的来源之一。但在休眠期,结核分枝杆菌分裂增殖基本停止,对脂肪酸的利用也转变为在巨噬细胞体内持续存活,提供结核分枝杆菌产生表型耐药的所需物质。结核分枝杆菌同时利用脂肪酸产生甘油三酯,并消耗甘油三酯为菌体供能。
3 脂肪酸作为结核分枝杆菌治疗新靶点的相关进展
3.1 脂肪酸是生物被膜的主要机制成分脂肪酸作为结核分枝杆菌治疗靶点的探究方向的提出,主要基于研究者们对生物被膜的研究和发现。细菌通过形成一个细胞群体的策略在不同环境条件下存活,这样的细胞群体就是生物被膜。超过80%的细菌感染存在生物被膜过程,病原菌通过这种生长方式长期留在宿主体内,躲避免疫系统和各种微生物治疗的清理作用,发展成为持续感染状态[43]。生物被膜作为人类感染的重要致病因素[44],越来越引起人们的重视,也是近年来慢性疾病致病机制不可忽视的研究要点。而结核分枝杆菌也能够形成生物被膜。目前已经得知,结核分枝杆菌的生物被膜基质成分主要包括多糖、eDNA、蛋白和脂质等[45]。脂肪酸作为细菌细胞壁脂质的主要组成部分和主要供能来源,对其的的研究足以作为药物靶点是新的治疗策略。
3.2 以脂肪酸为主体作为结核分枝杆菌靶点治疗的研究结核杆菌的细胞壁组成成分包含肽聚糖、阿拉伯半乳糖和MA。MA作为自然界重最大的脂肪酸,不仅在细胞壁干重占到了40%~60%的比例,也是结核杆菌细胞膜外层的基础。分枝菌酸的合成主要是在FASⅠ和FASⅡ 的相互作用之下。
此外,MA结构功能的改变也能影响结核杆菌在巨噬细胞中的生长状态。MA的合成与装配是可以为结核病的化学药物治疗提供许多潜在功能靶位的。烯酰ACP是MA合成中必经步骤的组成部分,已经被证明是抗结合治疗的重要药靶[46]
目前,人们采用的抗结核一线药物异烟肼,其主要的作用靶点是MA生物合成酶InhA,多聚磷酸盐polyP调控分枝杆菌的生理过程。其中也包括结核分枝杆菌生物被膜的形成。polyP能够影响到分枝杆菌对异烟肼的敏感性[47]。研究表明,poly P的缺失能够影响分枝杆菌生物被膜的形成能力,主要是能改变细菌表面的脂质成分[48]。不难推断出polyP或许影响分枝菌酸的代谢进一步影响了分枝杆菌生物被膜的形成。此外,polyP的缺失可导致C16和C17这些短链脂肪酸,也就是FASⅠ合成酶系统产生的中间产物,在生物被膜成熟时期呈现显著性缺乏。很多研究证明,结核分枝杆菌的持留性也与ployP有关,其代谢酶也可作为潜在的药物靶点[49]。比如以多聚磷酸盐激酶2作为药物靶点而设计出的新型药物[50]。
结核分枝杆菌的HupB蛋白主要定位在细胞膜上,与生物被膜形成的缺失和非极性的脂质谱改变相关。FASⅡ系统中的ACP转化酶(MCAT)也是脂肪酸代谢中的重要步骤。所以对于脂肪酸作为靶点治疗重点的研究,也可以深入到以具体蛋白位点或者蛋白表达为突破点。
4 展望
结核病是有记录以来人类最古老的疾病之一,尽管目前有减毒活疫苗和几种药物治疗结核,但它仍然是一种慢性的、危害严重的人畜共患传染病。结核分枝杆菌在感染期和休眠期的生长状态,繁殖及感染性存在较大差异,这种差异造成了对结核分枝杆菌研究的困难。脂肪酸是大多数生物体内所必需的物质,也广泛存在于结核分枝杆菌内。结核分枝杆菌将脂肪酸作为较稳定的能量来源,利用脂肪酸控制细胞壁的通透性,虽然处于不同时期的结核分枝杆菌的生理特性存在明显差异,其脂肪酸产生方式也存在差异,但对脂肪酸的利用等具有相似性。同时,目前证实控制细菌所采用的Ⅱ型脂肪酸合成系统的相关基因是保守且必不可少的,其中关键步骤所使用的同工酶执行的酶促反应相似[34]。
目前,围绕脂肪酸的形成以及其中涉及到的相关生化反应过程,如FASⅠ与FSAⅡ系统中涉及到的相关蛋白表达与位点的变异,这些都可以为脂肪酸作为结核分枝杆菌治疗的新靶点提供依据,为结核病的治疗提供新的途径。此后,也可以着眼于脂质形成的过程,细菌对于脂肪酸的利用与生产过程,或者其中涉及的相关酶蛋白表达或者是控制相关蛋白表达的具体位点基因,都可以作为新靶点寻找的重点。
猜你喜欢
锦州医科大学报(2021年8期)2021-11-18
昆明医科大学学报(2020年12期)2021-01-26
祝您健康(2018年12期)2018-11-27
浙江工业大学学报(2017年5期)2018-01-22
广东农业科学(2017年5期)2017-08-29
中国油脂(2017年12期)2017-01-18
吉林大学学报(医学版)(2015年3期)2015-12-17
护士进修杂志(2015年13期)2015-03-18
食品工业科技(2014年13期)2014-03-11
食品工业科技(2014年9期)2014-03-11
