
结核分枝杆菌感染中凋亡相关microRNA的研究进展
2021-08-10 12:25竺婷婷师红铃郭爱珍陈颖钰
中国兽医学报 2021年7期
苏 莉,竺婷婷,师红铃,郭爱珍,3,陈颖钰,3*
(1.华中农业大学 农业微生物学国家重点实验室,湖北 武汉 430070;2.华中农业大学 动物医学院,湖北 武汉 430070;3.华中农业大学 国家动物结核病专业实验室(武汉),湖北 武汉 430070)
结核病是全球重大传染病之一。据世界卫生组织(World Health Organization,WHO)最新统计,2019年全球新发结核病患者约1 000万,病死人数达到148万,其造成的影响对国家经济发展和公共卫生危害重大[1]。
结核分枝杆菌(Mycobacteriumtuberculosis,MTB)为结核病主要病原菌,当MTB感染宿主后,宿主调动一系列固有免疫和适应性免疫来抵御细菌入侵,如巨噬细胞吞噬病原体,产生抗菌肽、活性氧和活性氮等抗菌分子,使胞内形成一种酸性抗菌环境或通过TLR信号和相关炎性因子促进炎症应答以达到清除细菌的目的[2-3]。而结核分枝杆菌在与宿主互作的过程中,也产生一系列免疫逃避机制,包括抑制代谢相关反应、调节细胞凋亡、自噬、炎性因子产生等各种细胞生命进程来规避机体的免疫清除[4]。
细胞凋亡是机体为对抗病原而主动选择的一种程序性死亡(programmed cell death,PCD)。当宿主感染MTB后,巨噬细胞可以通过细胞凋亡破坏有利于细菌复制的胞内环境,从而清除胞内菌,且较之强毒株,弱毒株能更好的促进细胞凋亡[5]。miRNA是一类微小的非编码RNA,可通过降解mRNA或抑制翻译来调节转录后蛋白编码基因的表达。当MTB感染宿主后,机体巨噬细胞、树突状细胞中miRNA的表达谱发生显著变化。MTB通过上调或下调靶向凋亡相关基因的miRNA,抑制凋亡蛋白的合成最终改变细胞凋亡的发生。因此,miRNA介导的细胞凋亡是影响结核分枝杆菌在机体内存活的重要机制。现对结核分枝杆菌感染宿主细胞过程中凋亡相关的miRNA进行总结,以期为在RNA水平上了解结核分枝杆菌的感染机制和其作为结核病治疗靶标的能力提供参考。
1 miRNA概述
miRNA是长度约为22个核苷酸片段的非编码RNA,最初于1993年由LEE等[6]在秀丽隐杆线虫中发现。目前,在人类基因中已鉴定出超过2 500种成熟的miRNA(miRBase.org)。miRNA的生物发生通常是在细胞核中开始,经过RNA聚合酶、Dicer酶等加工后最终在细胞质内成为成熟的miRNA,进而被加载到由Ago2蛋白、Tar RNA结合蛋白(TRBP)以及KH型剪接调节蛋白(KSRP)等组成的RNA诱导沉默复合物(RISC)中[7]。RISC中miRNA 5′端的6~8个核苷酸“种子区”与靶基因mRNA转录物的3′-UTR中的互补序列结合,最终抑制靶基因翻译或导致其降解,使相应蛋白质合成受损。但是,近年来的研究发现,miRNA除了与mRNA的3′-UTR结合外,也可通过其他非经典途径参与基因的表达调控,如与mRNA的5′-UTR结合以促进降解作用[8]。在miRNA与靶基因相互结合的过程中,多种mRNA可以被同一个miRNA靶向结合,并且几个不同的miRNA也可以靶向同一mRNA,这种特性进一步增强了miRNA对靶基因的调控多样性。
2 MTB感染中调控凋亡的miRNA
miRNA在细胞凋亡的发生发展中发挥着重要的功能,通过调节凋亡相关基因如Bcl-2、Bfm、caspase-3/7、FOXO3(叉头样蛋白O3,forkhead box protein O3)等表达,以不同的途径对凋亡产生抑制或促进作用。如在小鼠巨噬细胞和MTB互作过程中,MTB通过激活TLR-2/MyD88/NF-κB信号通路诱导表达miR-27b,继而靶向Bag2基因最终控制细胞凋亡[9]。或是过表达miR-20b-5p,可负调控Mcl-1基因,促进细胞凋亡,抑制MTB的胞内生存等[10]。结核病中凋亡相关的miRNA见表1。
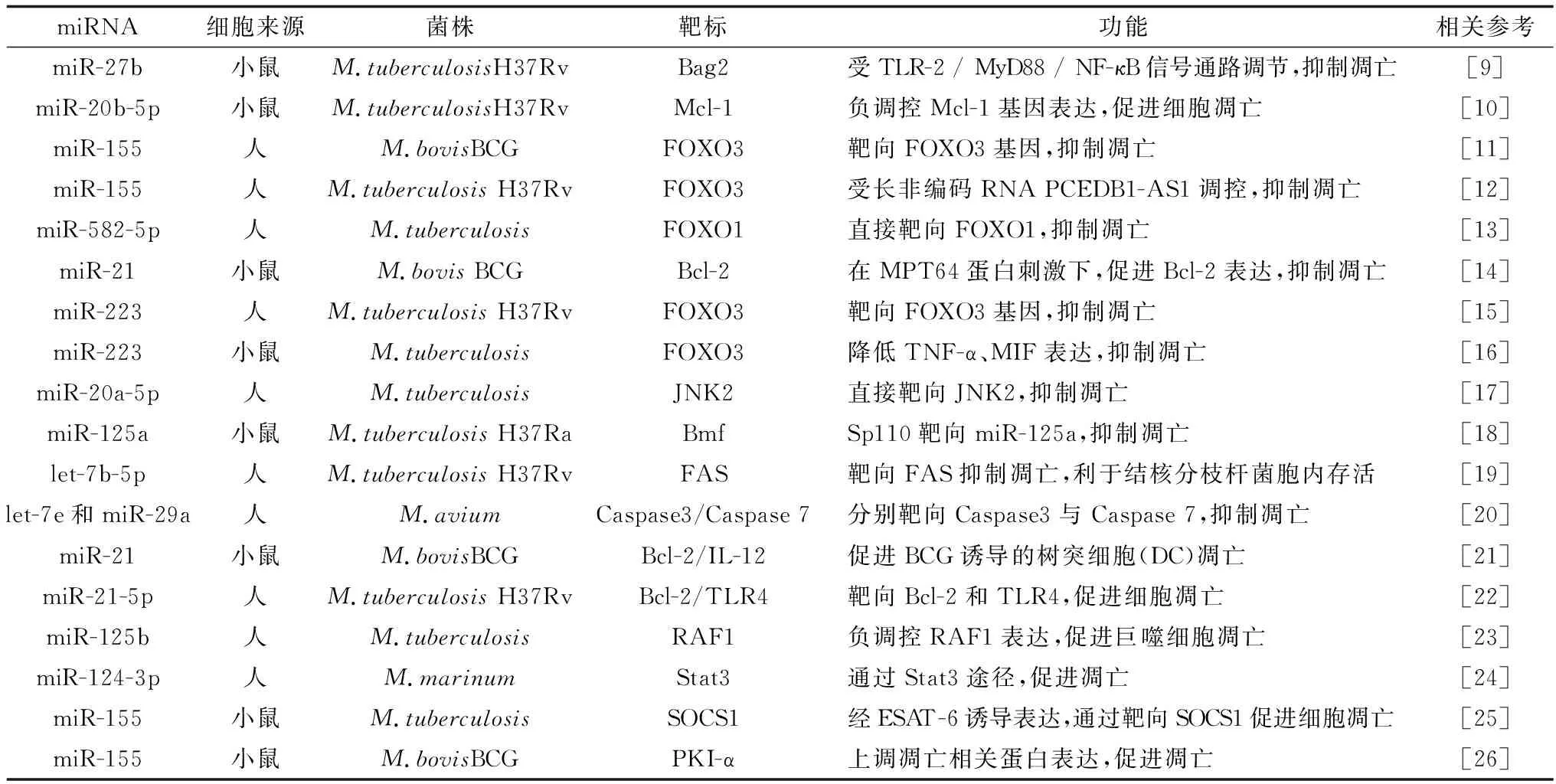
表1 结核分枝杆菌感染过程中与凋亡相关的miRNA
2.1 抑制细胞凋亡的miRNA
2.1.1miR-155 miR-155是一个多功能的miRNA,如可靶向SCOS1使MAPK信号通路失活,抑制肝脏星状细胞增殖[27],或靶向FOXO3调节妊娠高血压[28],其在多个疾病研究领域中的调控作用得到了广泛关注。FOXO3是参与调节细胞周期与细胞凋亡的重要转录因子,卡介苗(bacillus Calmette-Guérin,BCG)介导的巨噬细胞凋亡依赖于FOXO3激活。研究发现,在BCG感染的THP-1细胞中,miR-155表达量上调并靶向FOXO3基因,抑制了细胞的凋亡,使细菌逃避机体免疫应答[11]。此外,LI 等[12]证明在活动性肺结核患者中下调表达的长非编码RNA PCEDB1-AS1可直接靶向miR-155调控细胞凋亡,进一步证明了miR-155对凋亡的抑制作用以及减弱细胞清除MTB的抗菌能力。
2.1.2miR-582-5p miR-582-5p 在癌症疾病中有大量报道。最新研究发现,miR-582-5p可通过靶向ROCK1降低丙泊酚诱导的神经元凋亡,从而减轻对儿童的神经毒性作用[29]。但当前结核病中关于miR-582-5p调控细胞凋亡的研究不多。LIU等[13]发现,在活动性肺结核患者外周血单核细胞中miR-582-5p表达量上调。将miR-582-5p模拟物转染THP-1细胞后,凋亡率显著低于对照组,表明miR-582-5p可以抑制单核细胞的凋亡。叉头样蛋白O1(forkhead box protein O1,FOXO1)受Akt信号通路控制,可通过调节Fas配体、Bim蛋白或TNF凋亡相关的配体参与细胞的凋亡。研究者进一步对FOXO1的分析结果显示,miR-582-5p可靶向FOXO1下调其表达最终抑制凋亡,使MTB得以在细胞内生存。
2.1.3miR-21 miR-21在感染MTB的宿主细胞内差异表达,对机体抵抗病原体的免疫应答起着至关重要的作用。HACKETT等[30]发现,miR-21靶向PFK-M抑制IL-1β表达以及糖酵解代谢,进而减弱宿主对结核分枝杆菌的抵御。MPT64是结核分枝杆菌分泌的重要抗原蛋白之一,可剂量依赖性地抑制卡介苗纯蛋白衍化物诱导的RAW264.7细胞凋亡,促进Bcl-2的表达。此外,MPT64通过激活NF-κB信号通路,上调表达miR-21。与经典的miRNA抑制靶标表达不同,miR-21通过靶向Bcl-2的3′-UTR促进Bcl-2的表达以抑制凋亡[14]。这一通路的发现有助于更好的理解结核分枝杆菌的免疫逃避机制。
2.1.4miR-223 研究发现,相对于健康者,活动性结核病患者外周血巨噬细胞的凋亡率降低,并且感染MTB H37Rv株的巨噬细胞中miR-223表达显著增加,miR-223能够通过靶向FOXO3负反馈调节进而抑制凋亡,过表达FOXO3则显著逆转了miR-223对凋亡的抑制作用[15]。这一现象在小鼠的体内试验中也得到了证实[16]。这些结果揭示,结核分枝杆菌通过调节miR-223进而干预宿主巨噬细胞的凋亡,帮助其在体内扩散。
2.1.5miR-20a-5p/miR-125a ZHANG等[17]在活动性肺结核患者的CD14+单核细胞中发现14个下调的miRNA,通过抗结核治疗后,miR-20a-5p的降低可以逆转。进一步的试验证实,JNK2是miR-20a-5p的新型靶标,调节Bim蛋白表达触发巨噬细胞凋亡,有效清除细胞内的MTB。另有研究发现,细胞敲除miR-20a后,利用结核分枝杆菌H37Ra或H2O2进行刺激,与对照组相比,细胞凋亡率显著增加,说明miR-20a-5p对凋亡有抑制作用[31]。核体蛋白(Sp110)主要调节针对MTB感染宿主细胞的先天性免疫应答,过表达该基因可增强细胞的凋亡。在结核分枝杆菌H37Ra感染的情况下,Sp110影响多个miRNA的表达,其中miR-125a被显著下调。此外,Sp110可通过下调miR-125a促进Bmf的表达,调控细胞凋亡,更有效的清除胞内结核分枝杆菌[18]。
2.1.6let-7b-5p/let-7e/miR-29a let-7b-5p在MTB感染的THP-1细胞中可上调表达。抑制let-7b-5p表达显著提高THP-1细胞凋亡率[19]。FAS基因编码的FAS受体是细胞重要的死亡受体,与配体结合后形成死亡诱导信号复合物,在细胞凋亡中起调节作用。研究发现,let-7b-5p可靶向FAS基因使细胞凋亡能力减弱,增强MTB的胞内存活。SHARBATI等[20]通过与未感染的对照组相比,证实鸟分枝杆菌(Mycobacteriumavium,MAV)感染的巨噬细胞中半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,Caspase)活性显著降低,而let-7e与miR-29a表达量增多。靶标预测分析与荧光素酶报告试验指出Caspase3和Caspase7分别为has-let-7e和miR-29a的靶标。两者分别通过靶向作用对细胞凋亡的调节产生干扰。
2.2 促进细胞凋亡的miRNA
2.2.1miR-21 虽然有研究显示miR-21可作为细胞凋亡的负调控因子,但miR-21也存在促进凋亡的发生,这可能与不同的试验背景有关。WU等[21]比较了在正常和BCG疫苗接种的小鼠中miR-21的表达,发现与未感染组相比,BCG感染小鼠的肺中miR-21表达呈时间剂量依赖性增加。miR-21模拟物可增强BCG诱导的树突细胞(DC)凋亡。进一步试验发现,miR-21靶向Bcl-2促进树突细胞凋亡。此外,在MTB感染的THP-1和RAW264.7细胞模型中miR-21-5p表达量增加并通过靶向Bcl-2和TLR4基因,也可诱导细胞凋亡[22]。以上结果揭示,miR-21作为一个双向调控凋亡的重要RNA,在结核病临床药物靶点与标志物的选择上拥有巨大的潜力,但同时也说明其作用机制更为复杂,有待于进一步深入挖掘。
2.2.2miR-125b miR-125b在多个癌症发展中均参与调节,如靶向STAT3在皮肤鳞状细胞癌中充当肿瘤抑制因子,或调控肾癌细胞的侵袭和凋亡,亦或是通过抑制PI3K/AKT通路靶向HK2调节膀胱癌。在儿童肺结核研究中发现,外周血单核细胞中miR-125b表达下调。过表达miR-125b,则其靶基因RAF1表达降低且巨噬细胞凋亡率显著增加。RAF1是凋亡相关基因之一,通过激活MEK/ERK信号通路的方式抑制细胞的凋亡。据报道,抑制miR-125b时,RAF1表达升高,细胞活性降低[23]。这一结果表明,miR-125b有助于宿主对MTB的清除。
2.2.3miR-124-3p ZHOU等[24]发现,感染海洋分枝杆菌(Mycobacteriummarinum,MMA)的小胶质细胞中miR-124-3p的表达显著降低,过表达miR-124-3p,可上调Caspase-3或下调Bcl-2和Bcl-xl促进小胶质细胞凋亡。Stat3亦是miR-124靶标之一,研究发现其可通过Stat3相关途径调节细胞凋亡。这些为开发高效的抗结核性脑膜炎药物奠定基础。
2.2.4miR-155 miR-155在细胞凋亡调控中也具有两面性。ESAT-6是结核分枝杆菌早期分泌的抗原蛋白,在ESAT-6处理后的巨噬细胞中Caspase-3表达增加,且诱导miR-155表达,两者皆呈时间依赖性。SOCS1是细胞因子信号转导(SOCS)家族抑制剂中重要成员之一。在巨噬细胞中激活TLR2/NF-κB通路后,ESAT-6通过靶向miR-155-SOCS1途径促进巨噬细胞凋亡[25]。另据报道,感染了BCG的小鼠巨噬细胞中miR-155上调表达,而促凋亡基因PUMA、NOXA、BID、BAK1和SMAC的转录水平上调且细胞色素c与Caspase-3蛋白表达量增多。过表达miR-155可使细胞凋亡呈时间依赖性增强,表明miR-155促进凋亡[26]。
miR-155是当前MTB感染细胞后相关调控通路研究中较为全面的一个miRNA,由于不同感染模型能产生不同效应,因此在调控细胞凋亡中也具有双向作用。虽然尚未对其在巨噬细胞内的调控网络完全了解,但对于结核病的诊断治疗及预后仍具有重要意义。
3 miRNA在结核病中的诊断与治疗
在结核病防控过程中,随着越来越多miRNA机制被了解,将其应用到临床的研究也日益增多。首先,miRNA作为潜在生物标志物有效区分潜伏性结核患者、活动性结核患者和健康者已被广泛报道。在一项活动性肺结核患者和健康供体的外周血样本检查研究中发现,miR-199b-5p和miR-582-5p表达明显失调,进一步通过ROC曲线评估诊断性能表明这些miRNA有望成为活动性结核病诊断生物标志物[32]。LATORRE等[33]提出了一种诊断结核病的新型全血miRNA标志(hsa-miR-150,hsa-miR-21,hsa-miR-29c和hsa-miR-194),且具有91.21%的敏感性(90.8%~91.62%)和87.95%的特异性(87.37%~88.52%)。对于儿童结核病,也有研究表明miRNA可作为其早期诊断的有效生物标志物。其次,在疾病治疗方面,对于异常表达的miRNA可利用其抑制物或模拟物恢复正常表达。这种新型的干预策略通过对单个基因或者整个细胞进程产生影响从而达到治疗疾病的目的。MOORE等[34]通过利用聚乳酸-乙醇酸纳米粒子体内递送miR-223模拟物,抑制了巨噬细胞融合的形成,证明负载miR-223模拟物的纳米颗粒为可作为巨噬细胞融合的治疗抑制剂。此外,将外泌体样纳米囊泡携带miRNA-150传递至效应T细胞,也可以抑制小鼠接触敏感性[35]。因此,miRNA通过外源操作以增强宿主抵抗微生物感染的免疫力并提供潜在新型抗菌方法具有极大潜能。但是,关于miRNA最适合的传递途径以及药物安全性仍有待研究。
4 结束语
当前结核病的诊断治疗对全球公共卫生依旧是一个巨大的挑战,需要更为高效精准的治疗方案。miRNA的差异表达作为诊断结核病的生物标志物的潜力以及其靶向宿主基因调节宿主蛋白合成作为新型治疗靶标的研究日益增多,但依旧存在一定问题。有研究通过将结核病、结节病以及健康患者的全血进行miRNA表达谱分析,发现与健康对照组比较,结核病、结节病组的miRNA差异表达显著,但是结核病、结节病2组内的miRNA表达谱相似度高[36]。故而利用miRNA作为诊断的生物标志物时要注意病理或病原相似的疾病。另一方面,许多miRNA共同对细胞凋亡有抑制或促进作用,能否将多个功能类似的miRNA联合应用,以及联合应用中共同调控网络的研究目前较少。有些miRNA如miR-155或miR-21在凋亡调控中还具有两面性,需结合实际,具体问题具体分析。因此,作为治疗结核病的新兴领域,未来miRNA的临床应用需要我们进一步更深入的探索与研究。
猜你喜欢
保健医苑(2022年5期)2022-06-10
医学概论(2022年4期)2022-04-24
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
特别健康(2018年4期)2018-07-03
中国医学装备(2016年6期)2016-12-01
实用皮肤病学杂志(2015年4期)2015-12-22
中国卫生(2015年1期)2015-11-16
医学研究杂志(2015年12期)2015-06-10
中国当代医药(2015年36期)2015-03-11
