
贝莱斯芽孢杆菌zk1对鹰嘴桃果肉组织的影响及酶学特性
2021-07-28 07:00陈少先曾瑶英
食品与机械 2021年7期
陈少先 - 刘 悦 曾瑶英 - 余 倩
(仲恺农业工程学院食品学院,广东 广州 510225)
贝莱斯芽孢杆菌(Bacillusvelezensis)属于芽孢杆菌属(Bacillus)的一个亚种,是革兰氏阳性好氧细菌,菌体成杆状,内生孢子,在自然界中广泛分布[1]。近年来,贝莱斯芽孢杆菌主要作为生防菌在报道,国内外相关研究大多集中在促进动植物生长、拮抗病原菌、诱导系统抗性、抑菌物质及其基因簇鉴定、拮抗作用机制等方面[2-5],在抑制病原菌和生物防治方面具有显著优点。实验室在腐败的鹰嘴桃中分离并鉴定出一株贝莱斯芽孢杆菌,将其命名为贝莱斯芽孢杆菌zk1(Bacillusvelezensiszk1),该菌对鹰嘴桃的致腐能力极强,而关于贝莱斯芽孢杆菌作为腐败菌的报道极少。
果蔬的腐烂通常是由病原菌分泌的细胞壁降解酶[6]通过对细胞壁间多糖物质降解,破坏细胞结构,使果实软化,引起致腐。宋喜霞[7]判断果胶甲基反式消除酶和多聚半乳糖醛酸酶在锈腐菌侵染人参过程中起了重要作用;李宝聚等[8]证明果胶酶、纤维素酶在黄瓜黑星病菌致病中起着重要作用,是主要的致病因子之一;也有文献[9]报道灰葡萄孢菌株的致病力与β-葡聚糖苷酶具有相关性;李喜玲[10]证明了灰葡萄孢菌株的致病力与纤维素酶活力呈正相关,灰霉菌果胶酶活力高低与致病力强弱有一定的相关性。以上研究都表明了腐败菌分泌的酶类是侵染宿主的关键致病因子,此外,病原菌在侵入宿主初期会遇到细胞壁屏障,病原菌释放细胞壁降解酶,从而有利于其入侵,也为其他微生物的入侵提供便利,加速寄主的腐败。
国内外对核果果实采后病害的病理生理和采后病害控制技术方面已经做了大量的研究[11-12],但对降解细胞壁酶类的酶学性质研究较少,且在关于贝莱斯芽孢杆菌对水果的致腐作用目前尚未报道。研究拟检测菌株Bacillusvelezensiszk1致病后导致鹰嘴桃成分和果肉超微结构的变化,有针对性地开展其中相关酶的酶学特性研究,以期得到该菌株中与植物致病相关酶的最适反应温度、pH值和金属离子信息,为进一步解释其致病的机制提供理论依据。
1 材料与方法
1.1 材料与试剂
贝莱斯芽孢杆菌zk1(Bacillusvelezensiszk1,zk1):中国普通微生物菌种保藏管理中心;
鹰嘴桃:广东省河源市连平县三角湖果园;
营养琼脂(NA)和营养肉汤(NB):青岛海博生物技术有限公司;
木聚糖:分析纯,美国Sigma试剂公司;
无水葡萄糖、半乳糖醛、D-木糖、淀粉、羧甲基纤维素钠等:分析纯,国药集团化学试剂有限公司。
1.2 试验试剂和仪器
生化培养箱:SPX-150型,上海悦丰仪器仪表有限公司;
立式自动电热压力蒸汽灭菌锅:LX-C35L型,合肥华泰医疗设备有限公司;
高速冷冻离心机:H2500R-2型,上海卢湘仪离心机仪器有限公司;
冻干机:HX-18型,上海沪析实业有限公司;
超净工作台:SW-CJ-IF型,上海博迅实业有限公司医疗设备厂;
电热恒温干燥箱:DHG-942D0A型,浙江赛德仪器设备有限公司。
1.3 试验方法
1.3.1 菌悬液制备 贝莱斯芽孢杆菌zk1先用营养琼脂培养基复活,再用营养肉汤培养液在37 ℃、120 r/min震荡培养24 h,传代两次,比浊法调整菌数为104CFU/mL。
1.3.2 鹰嘴桃的成分检测
(1) 样品前处理:挑选同一批次的健康鹰嘴桃,采用菌液刺伤接种法,在赤道部位接种6个点/果,常温保湿(RH≥85%)培养48 h。用无菌刀具削取每果赤道部位组织(带果皮),修切成10 mm×10 mm×10 mm的小方块,冷冻干燥72 h,打粉(2 800 r/min,5 min),过60目筛备用。以注射无菌蒸馏水的鹰嘴桃为对照组。
(2) 成分检测:淀粉含量按GB 5009.9—2016的酸水解法执行;纤维素、半纤维素和果胶含量按NY/T 3165—2017的滤纸袋法执行。
1.3.3 贝莱斯芽孢杆菌zk1致病鹰嘴桃的透射电镜(TEM)观察 用菌液刺伤感染的方式,将菌株zk1接种到健康的鹰嘴桃,喷无菌蒸馏水保湿,37 ℃培养24 h。然后在超净工作台切取病健交接部位的组织,用刀片修整为1 mm×1 mm×3 mm的样品块,整个取样过程不超过2 min,即刻放入含2.5%戊二醛的磷酸—柠檬酸缓冲液溶液(pH 7.4)固定24 h。对照组用无菌蒸馏水接种健康鹰嘴桃果实并在同样条件下培养,参照病果的取样部位,切取对照组的桃果肉组织样品。
按照常规TEM样品制备方法[13]略作改动。1%四氧化锇将待测样品块固定6 h,含2.5%戊二醛的磷酸—柠檬酸、酸缓冲液溶液(pH 7.4)漂洗2 h,用30%,50%,70%,80%,90%,95%,100%乙醇浸泡10 min进行梯度脱水,氧化丙稀和丙稀环氧树脂稀释液浸透,Epon812包埋;超薄切片机定位并切成70~100 nm超薄切片。然后用柠檬酸铅和醋酸铀电子染色,最后用透射电镜观察。
1.3.4 标准曲线制作 淀粉酶活测定采用YOO改良法[14];果胶酶活力测定参照文献[15];纤维素酶活力测定按GB/T 23881—2009执行;木聚糖酶活力测定按GB/T 23874—2009执行。以每分钟降解底物释放1 μmol葡萄糖(或D-半乳糖醛酸、木糖)所需的酶量定义为一个酶活单位(U)。
根据以上方法制作标曲。葡萄糖标准曲线线性回归方程为y=12.46x+0.248 4,R2=0.996 7;半乳糖醛酸标准曲线线性回归方程为y=1.463x+0.072 9,R2=0.997 4;木糖标准曲线线性回归方程为y=1.579x+0.067 8,R2=0.998 3。
1.3.5 粗酶液制备及酶活力测定 吸取1 mL菌悬液加入100 mL含有1.00 g淀粉的营养肉汤培养基,37 ℃,120 r/min震荡培养12 h,诱导菌在培养基中产酶。5 000 r/min离心20 min后得到的上清液即为淀粉酶粗酶液。4 ℃保存备用,在48 h内使用。将淀粉分别替换成羧甲基纤维素钠、果胶、木聚糖,制得纤维素酶粗酶液、果胶酶粗酶液、木聚糖酶粗酶液。
试验前先将待测粗酶液、底物溶液(1.0 g/100 mL)于37 ℃预热10 min。在25 mL具塞比色管中依次加入1 mL 底物溶液、1 mL柠檬酸—磷酸氢二钠缓冲液、0.5 mL 粗酶液和1.5 mL去离子水,振荡3~5 s。于40 ℃ 水浴锅水浴30 min,再加入2 mL DNS试剂终止反应,沸水浴10 min,立刻用水冷却至室温,加水定容至25 mL。在540 nm处测定吸光值A1。每个试样进行3次平行试验,结果取平均值,保留3位有效数字。淀粉酶活性的测定底物为1%淀粉溶液,纤维素酶活性的测定底物为1%羧甲基纤维素钠溶液,果胶酶活性的测定底物为1%果胶溶液,木聚糖酶活性的测定底物为1%木聚糖溶液。
空白测定:粗酶液沸水浴灭活5 min,其余步骤同上,吸光值记为A0。
淀粉酶、纤维素酶、果胶酶、木聚糖酶酶活性按式(1)计算:
(1)
式中:
X——粗酶液的酶活力,U/mL;
A1——粗酶液反应的吸光值;
A0——空白样反应的吸光值;
K——标准曲线的斜率;
C0——标准曲线的截距;
t——酶解反应时间,min;
M——摩尔质量。
1.3.6 贝莱斯芽孢杆菌zk1酶学性质研究
(1) 酶促反应最适温度:在pH 5.0的条件下,将粗酶液与柠檬酸—磷酸氢二钠缓冲液混合,分别以30,40,50,60,70,80,90 ℃为反应温度,反应30 min,测定相应的酶活力。每处理做3个平行,取平均值计算酶活数值,最高酶活力值记为100%,计算出各温度下的酶活力。
(2) 酶促反应最适pH值:在最适温度条件下,将粗酶液分别与用pH 3.0,4.0,5.0,6.0,7.0,8.0,9.0,10.0的缓冲剂调配的底物混合,作用30 min,测定相应的酶活力,每处理做3个平行,取平均值计算酶活数值,以酶活力最高值为100%,计算各个pH条件下的酶活力。
(3) 金属离子对酶活性的影响:参照崔海洋等[16]的方法并略作修改,在最适温度和pH条件下,在粗酶液中分别加入0.15 mmol/L金属离子(Na+、K+、Ca2+、Cu2+、Mn2+、Fe2+、Ba2+、Mg2+)化合物、氯化物及硫酸盐,测定相应的酶活力,每处理做3个平行,取平均值计算酶活数值,以未添加金属离子的粗酶液酶活数值为100%,计算各金属离子下的酶活力。
2 结果与分析
2.1 鹰嘴桃成分检测
在植物体内,质体中包含的淀粉粒是多糖的主要贮藏形式,先前有研究[17]发现桃果肉细胞中淀粉粒还存在于细胞间隙。而果胶会与半纤维素、纤维素形成交联结构,起到维持细胞骨架形态的作用[18]。由图1可知,鹰嘴桃被菌株zk1侵染后,果实中的淀粉、纤维素、半纤维素和果胶含量都出现了一定程度的减少,推测菌株zk1具备较强的糖苷水解和多糖裂解能力。以鹰嘴桃果肉为营养来源时,该菌在生长过程中逐渐产生相关酶类,用来消耗培养环境中的淀粉、纤维素、半纤维素和果胶等营养物质。当其附着在鹰嘴桃等水果表面,与之接触的部位就成为了营养供给体,接触部位的果肉组织随着营养物质的消耗而逐渐坍塌腐烂,最终果肉大面积腐败。
2.2 贝莱斯芽孢杆菌zk1致病鹰嘴桃的扫描电镜观察结果
如图2(a)所示,鹰嘴桃接种无菌水培养24 h后,果肉组织结构未受到破坏,果肉细胞完整,排列整齐。细胞之间有小间隙,大液泡占据细胞内部的主要面积,其余细胞器被液泡挤到边缘,丰富的细胞内含物紧靠细胞壁分布。果肉细胞壁紧密,细胞壁之间的中胶层是高密度的结构,细胞壁与中胶层在透射电镜下成像明暗分明,可清楚辨别。液泡膜将液泡和其他细胞内含物隔开,靠近细胞壁的细胞内含物中包括了叶绿体、线粒体、质体和淀粉颗粒。所有细胞内含物均保持了正常的形态。叶绿体为两端较尖长条形细胞器,长度约1 μm。线粒体是圆形的细胞器,可以看到其内有突起嵴。
如图2(b)所示,鹰嘴桃果肉被菌株zk1侵染后,果肉组织则遭到了严重的损坏。细胞壁致密性降低,部分结构降解导致细胞整体结构松散,细胞壁与中胶层之间的明暗对比度淡化。中胶层溶解并与细胞壁剥离,细胞间隙加大,细胞壁出现漂移。部分细胞壁弯曲,细胞内含物进入细胞壁,相邻的细胞之间的聚合度明显下降。细胞器已出现降解,细胞壁空腔化,叶绿体与线粒体发生变形,大部分线粒体破裂,叶绿体片层结构不清晰。液泡膜消失,液泡与质体互溶。大淀粉颗粒被分解成零散小颗粒。
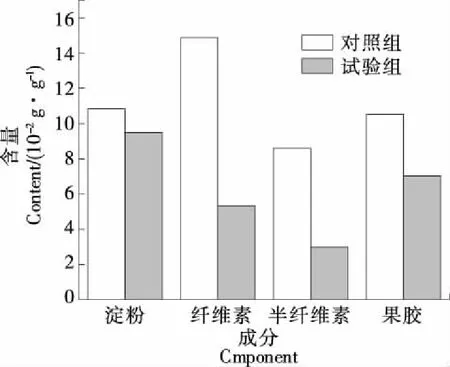
图1 鹰嘴桃成分检测结果
从图2可以看出,细菌zk1对鹰嘴桃的细胞有极强的破坏能力,可以在短短的24 h内将健康的鹰嘴桃果肉组织瓦解。病变果肉细胞具体表现为细胞壁、中胶层、淀粉颗粒等细胞内含物和结构组织的损坏。细胞壁中存在木聚糖、甘露聚糖、半乳聚糖和阿拉伯聚糖等聚多糖[19]30-33,果胶和纤维素分别是中胶层和微纤丝的主要成分[20-21],结合鹰嘴桃被侵染前后的成分检测,推测细菌zk1应该具有分解这些成分的能力。
2.3 酶学性质
2.3.1 酶促反应最适温度 如图3所示,菌株zk1能针对不同的外源诱导物产生相应的酶(粗酶液),且这些酶对温度的适应能力不同。木聚糖酶在40 ℃下的相对酶活数值为0.036 1 U/mL。果胶酶的最适温度为50 ℃,达到0.102 U/mL。反应温度为60 ℃时,纤维素酶和淀粉酶的相对酶活力最优,分别为0.831 0,0.336 2 U/mL。由此可见,此菌株在较高的反应温度条件下,纤维素酶和淀粉酶依然存活。半纤维素是植物细胞壁中广泛存在的杂聚多糖,而木聚糖是半纤维素的主要组成部分[19]9。将菌株zk1接种鹰嘴桃后,主要是在37 ℃培养,接近试验中的40 ℃。根据酶促反应最适温度的结果,该菌株对鹰嘴桃的致病作用可能是依赖于木聚糖酶对桃果细胞壁中木聚糖的水解功能,从而破坏了桃果细胞壁的半纤维素结构网络。值得注意的是,在40 ℃下,淀粉酶依然保持了较好的酶解能力,酶活力有0.694 6 U/mL。而此温度下,纤维素酶的活力为0.294 6 U/mL,与一株拥有高效纤维素酶基因的枯草芽孢杆菌工程菌的酶活力检测数值接近[22]。

CW. 细胞壁 CC. 细胞内含物 V. 液泡 ICS. 细胞间隙 P. 质体 M. 线粒体 Chl. 叶绿体 ML. 中胶层 S. 淀粉粒 T. 液泡膜

图3 温度对酶活性的影响
2.3.2 酶促反应最适pH值 如图4所示,与最适温度结果(图3)类似的是,果胶酶对反应条件的变化最敏感。随着外部反应条件因素的改变,果胶酶活力的变化幅度最大。与果胶酶的最适pH值一样,纤维素酶的最适pH值亦为6。在最适pH值下,纤维素酶活力为0.248 3 U/mL,而果胶酶的活力为0.150 6 U/mL。淀粉酶在中性反应条件(pH 7)下有最高酶活力,达到0.458 7 U/mL。木聚糖酶则表现出偏向碱性条件(pH 8)的耐受能力,这与一些菌株的木聚糖酶学特性研究结果不同。孙超[23]研究发现,桔青霉的木聚糖酶最适pH为5.5。其中的原因除了菌株之间存在物种基因差异,还可能是因为研究的菌株zk1分泌的木聚糖酶具有独特的酶结构。
2.3.3 金属离子对酶活性的影响 如图5所示,除了Mn2+外,其余金属离子能促进淀粉酶的酶活力。同时,Mn2+也是纤维素酶的抑制剂。由此推测,菌株zk1所分泌的淀粉酶和纤维素酶具有相似的结构,Mn2+能通过与氨基酸分子—SH基结合,使酶分子失活或变性[24]。但对于果胶酶而言,Mn2+则是一种激活剂,可令其相对酶活力超过114%。这可能是因为金属离子与酶分子氨基酸侧链的氧原子相结合,通过稳定的键作用力增强了酶活性区域的构象结构,或者形成了酶与底物的复合体[25]。试验中,不同的金属离子并未对木聚糖酶的酶活力造成负面影响,但林小洪等[26]研究发现,Mn2+和Cu2+对木聚糖酶具有明显的抑制作用;杨颖等[27]研究发现,Mn2+、Ba2+、Mg2+、Cu2+对来源于AT24的木聚糖酶具有激活作用,而对来源于AT22-2的木聚糖酶却具有抑制作用,这可能与不同木聚糖酶的结构差异有关,具体机理还需要进一步的研究论证。

图4 pH值对酶活性的影响
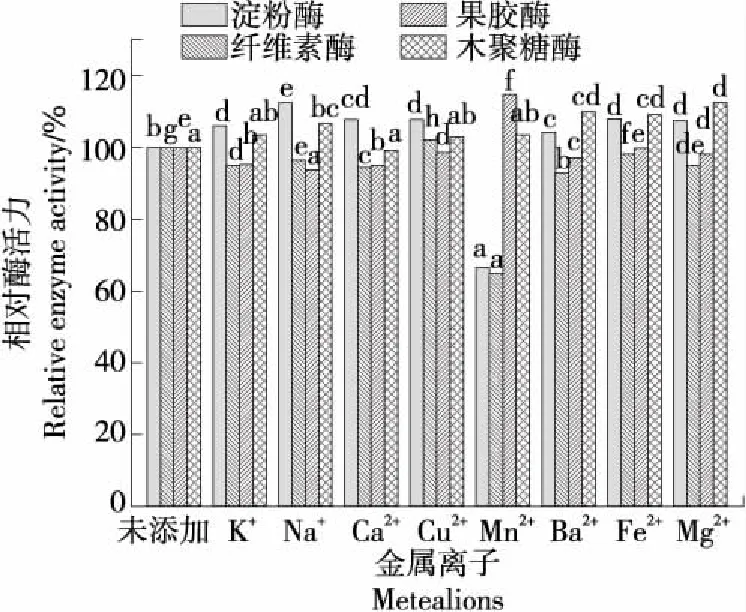
同一种酶的小写字母不同表示邓肯(D)法检验在P<0.05水平差异显著
3 结论
贝莱斯芽孢杆菌zk1侵染鹰嘴桃后,鹰嘴桃果肉细胞结构被破坏,出现变形、细胞器破裂、溶解等现象。推测该菌通过分泌淀粉酶、纤维素酶、木聚糖酶和果胶酶来降解鹰嘴桃的果皮的纤维素和果肉中的淀粉、果胶等,从而使鹰嘴桃腐烂。酶学特性研究的结果显示该菌分泌的淀粉酶最适反应温度为60 ℃,最适反应pH值是7,0.15 mmol/L 的Mn2+可抑制淀粉酶的活力;纤维素酶最适温度为60 ℃,最适反应pH值是6,Mn2+也可对其起抑制作用;木聚糖酶最适反应温度为40 ℃,最适反应pH值是8,多种金属离子均是激活剂;果胶酶最适反应温度为50 ℃,最适反应pH值是6,Mn2+对其激活效果最优。试验探究了贝莱斯芽孢杆菌zk1对鹰嘴桃的致病机理及其酶学性质,但未具体掌握该菌株降解植物细胞壁时各种多糖水解酶的结构,之后的研究方向可以考虑在这些方面继续挖掘。
猜你喜欢
农业科技通讯(2022年2期)2023-01-05
林产化学与工业(2022年4期)2022-09-13
实用手外科杂志(2022年2期)2022-08-31
粮食加工(2021年5期)2021-11-01
中国饲料(2021年2期)2021-02-23
食品与机械(2020年8期)2020-09-18
湖南饲料(2019年5期)2019-10-15
中外葡萄与葡萄酒(2018年6期)2018-11-14
食品安全导刊(2018年24期)2018-09-26
广东第二课堂·小学(2017年1期)2017-02-20
