
小兴安岭针阔混交天然次生林主要树种NSC变异来源分析
2021-07-20 20:31王晨李耀翔王子纯孟永斌
森林工程 2021年3期
王晨 李耀翔 王子纯 孟永斌

摘 要:為探究小兴安岭针阔混交天然次生林优势树种非结构性碳水化合物质量分数随树种、器官和间伐强度的变化特征,进而了解林带间伐后的非结构性碳水化合物(nonstructural carbohydrates,NSC)分配格局、生长状况、适应策略和恢复潜力,在小兴安岭针阔混交天然次生林6种间伐强度(0%,15%,20%,25%,30%,35%)的林带中,选取5个主要树种, 采集树木枝条、树干和树叶, 测定其可溶性糖类及淀粉质量分数, 分析其NSC各组分随树种、器官和间伐强度的变化特征。结果表明:不同树种之间可溶性糖、淀粉和总NSC质量分数均存在显著性差异。所有NSC组分在不同树种中变异均较高,云杉、色木槭、青楷槭 、红松NSC各组分的变异系数均大于30%, 最高达88%。不同器官间NSC质量分数差异显著,其中可溶性糖质量分数在树叶中最高, 树干中质量分数最低,淀粉和总NSC质量分数在树干中最高, 树枝中最低。NSC各组分在不同器官中变异较低,均低于35%。间伐强度对各NSC组分影响亦显著。所有NSC组分在不同间伐强度中变异均较高, 可溶性糖变动范围为67.07%~80.99%,淀粉为44.77%~53.55%,总NSC质量分数为35.14%~42.08%。变异来源分析显示,可溶性糖、淀粉和总NSC质量分数主要受器官影响, 树种、器官和其交互作用对NSC不同组分影响均达到极为显著水平(P<0.01),间伐强度对NSC不同组分影响均达到显著水平(P<0.05)。综上,树种、器官和间伐强度是影响小兴安岭针阔混交天然次生林树木NSC各组分质量分数的主要因素,其中器官是影响各NSC组分质量分数的主导因素。
关键词:小兴安岭;抚育间伐;天然次生林;非结构性碳水化合物;可溶性糖;淀粉
中图分类号: S753;S718.5 文献标识码:A 文章编号:1006-8023(2021)03-0036-08
Abstract:In order to explore the variation characteristics of the nonstructural carbohydrate concentration of dominant tree species in the coniferous and broadleaved natural secondary forest in the Xiaoxingan Mountains with tree species, organs and thinning intensity, and to understand the distribution pattern, growth status, adaptation strategies and recovery potential of the NSC after the forest belt thinning, the coniferous and broadleaved natural secondary forest in the mixed natural secondary forest with 6 kinds of thinning intensity (0%, 15%, 20%, 25%, 30%, 35%), select 5 main tree species, collect tree branches, trunks and leaves, and determine their soluble sugar and starch concentration, analyze the variation characteristics of NSC components with tree species, organs and thinning intensity. The results showed that there were significant differences in the concentration of soluble sugar, starch and total NSC among different tree species. All NSC components have high variation in different tree species. The coefficients of variation of the NSC components of Picea asperata, Acermono, Acer tegmentosum and Pinus koraiensis were all greater than 30%, and the highest was 88%. There were significant differences in NSC concentration among different organs. Among them, the concentration of soluble sugar was the highest in the leaves and the lowest in the trunk. The concentration of starch and total NSC was the highest in the trunk and the lowest in the branches. The various components of NSC had low variation in different organs, all of which were less than 35%. The intensity of thinning also had a significant effect on each NSC component. All NSC components had high variation in different thinning intensities. The variation range of soluble sugar was 67.07%~80.99%, starch was 44.77%~53.55%, and the total NSC concentration was 35.14%~42.08%. The analysis of the source of variation showed that the concentration of soluble sugar, starch and total NSC were mainly affected by organs. The influence of tree species, organs and their interaction on different components of NSC had reached a very significant level (P<0.01), and the intensity of thinning had an effect on different components of NSC which reached a significant level (P<0.05). In summary, tree species, organs and thinning intensity were the main factors that affect the concentration of NSC components in the natural secondary forests of the coniferous and broadleaved mixed natural secondary forests in Xiaoxingan Mountains, and organs were the dominant factors affecting the concentration of each NSC component.
Keywords:Xiaoxingan Mountains; tending thinning; natural secondary forest; nonstructural carbohydrates; soluble sugar; starch
0 引言
森林光合作用和呼吸作用之间的平衡决定了森林的碳汇。光合产物在树木体内的运输和分配调控森林的生产力和碳汇功能。植物光合作用产生的碳水化合物分为结构性碳水化合物(structural carbohydrates,SC)与非结构性碳水化合物(nonstructural carbohydrates,NSC)[1]。SC主要用于构成植物的基本框架,包括木质素、纤维素与半纤维素等;NSC主要包括可溶性糖与淀粉,用于植物的生长和代谢[2]。NSC质量分数的高低可以反映植物整体的碳需求、碳供应水平以及对环境的适应能力, 是研究碳循环机理和过程中不可缺少的参数,也是表征树木碳源—碳汇平衡的重要指标[3]。
NSC的质量分数与分配特征受多种因素影响[4-5]。树种是NSC质量分数及其分配特征的主要影响因素之一,不同树种对NSC利用和存储策略不同,导致其NSC质量分数存在较大差异[6]。器官也是影响NSC质量分数及分配特征的主要因素之一,不同器官NSC质量分数是不同的,不同树木器官间的功能性一定程度上决定了NSC各组分质量分数的差异[7]。马履一等[8]、刘思泽等[9]、张甜[10]、王杰等[11]的研究结果表明抚育间伐会改变林分结构和土壤理化性质,改变森林的光照、温度、水分和CO2体积分数等条件,而光照、温度、水分和CO2体积分数等条件的改变,会对树木NSC质量分数产生影响[12]。
目前开展的碳汇研究多是基于人工林,而对天然次生林的碳汇研究较少,特别是有关树种、器官和间伐强度对天然次生林NSC质量分数影响的研究尚未见报道。因此本文通过研究小兴安岭针阔混交天然次生林树种、器官及间伐强度是否影响NSC各组分质量分数,以及树种、器官及间伐强度对NSC各组分质量分数的影响是否存在差异,从而了解树木碳水化合物的供应与转化的情况以及对生存环境的适应策略。
1 研究地区与研究方法
1.1 研究区概况
试验区域位于小兴安岭地区带岭林业实验局东方红林场414林班1—3小班。试验区平均海拔600 m,坡向为西北方向,坡度14°。气候类型为大陆性湿润季风气候,年均气温1.4 ℃, 其中,平均最低温度是在1月上旬,达到-40 ℃;最高气温是在7月,达到37 ℃, 全年年平均降水661 mm, 主要集中在7—8月份。土壤以暗棕壤为主,少量林地为谷地草甸土和沼泽土,土壤厚度约30 cm。林分类型为天然针阔混交次生林,乔木树种约20种,以冷杉(Abies fabri)、色木槭(Acer mono)、云杉(Picea asperata)、青楷槭(Acer tegmentosum)、水曲柳(Fraxinus mandshurica)、黄波椤(Phellodendron amurense)、红松(Pinus koraiensis)、杨树(Populus)和花楷槭(Acer ukurunduense)为主[10]。
2012年在生境相对一致的研究区域依据林分密度与蓄积量进行带状抚育间伐,并设置了6块100 m×100 m的试验样地,间伐强度分别为0%、15%、20%、25%、30%、35%(样地编号分别为CK,A,B,C,D,E),应用下层抚育法伐除濒死木、被压木及干形不良的林木。2019年8月进行样地复查,在6种间伐强度的样地内设置20 m×20 m的标准试验样地各3块,共计18块。试验样地均在同一坡面,坡向西北,坡度14°,平均林龄70 a,GPS记录海拔坐标。间伐林分基本状况见表1。
1.2 主要树种的确定
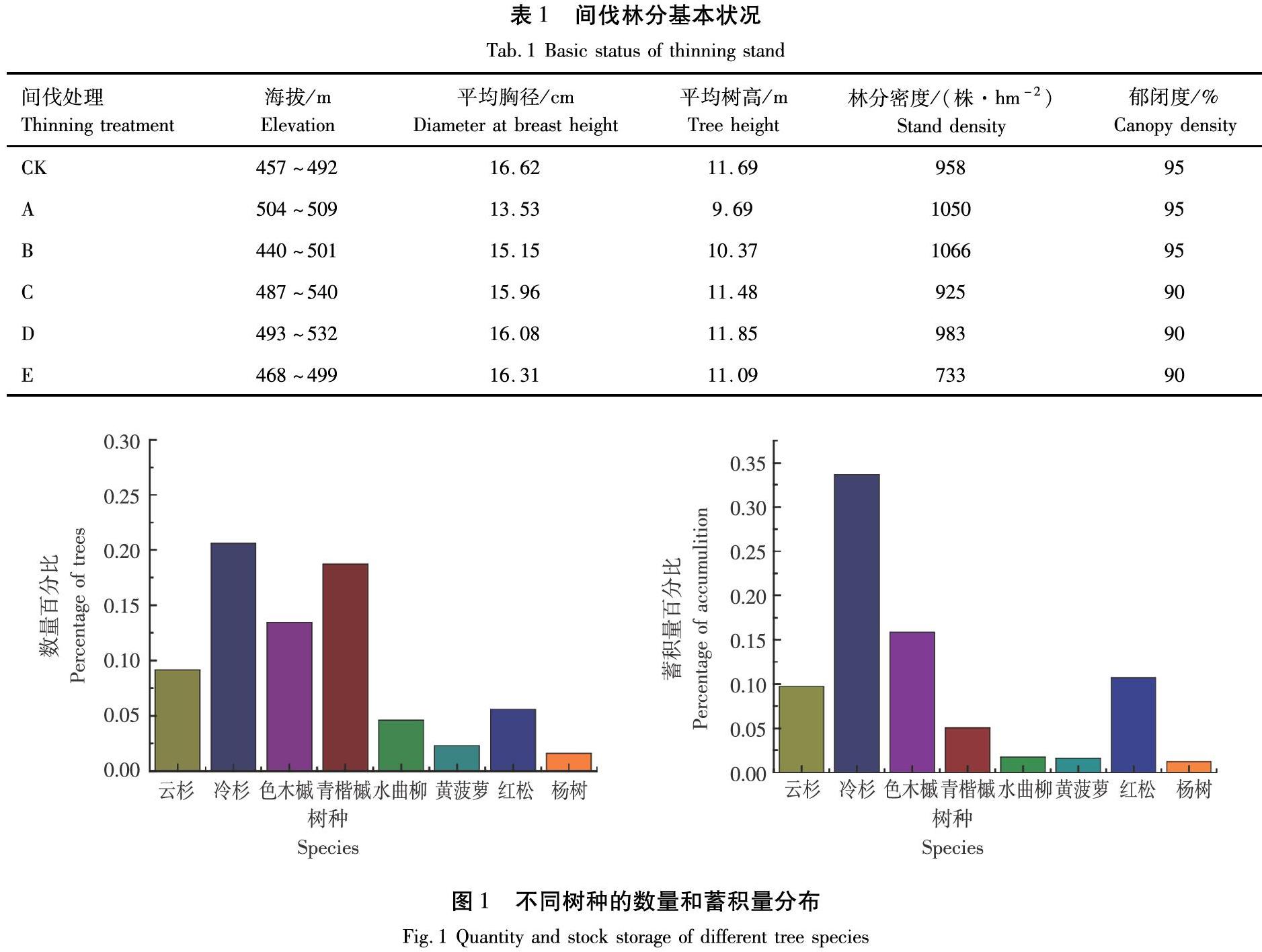
对样地内林木进行每木检尺。检尺内容包括树种、胸径、树高、冠幅、郁闭度和林分生长状况等。本次试验样地树种组成较复杂,每木检尺数据结合通用二元材积模型[13]可得各树种蓄积量。选取数量和蓄积量较大的云杉、冷杉、色木槭、青楷槭和红松5个树种作为该林分的主要树种进行测定(图1)。
1.3 样品采集与测定
样品采集:2019年8月初, 在样地选择生长状况相似的林木,使用内径5 mm生长锥采集离地面1.3 m处的树干树芯,使用高枝剪在树冠中随机剪取长势良好的树枝、树叶。每份取100 g左右待测样品, 即刻放入低温便携箱, 带回实验室处理(105 ℃杀青30 min, 65 ℃烘干48 h至恒重, 磨粉储存在广口瓶待测)。
非结构性碳水化合物中, 可溶性糖和淀粉比例达90%以上[14], 故本研究将NSC质量定义为可溶性糖和淀粉质量之和。测定方法采用苯酚浓硫酸法[15]。
可溶性糖的提取:取0.1 g样品, 加入80%乙醇10 mL, 于沸水浴中提取10 min, 然后4 000 r/min离心10 min, 收集上清液于50 mL容量瓶中, 反复3次, 合并上清液, 定容, 即为可溶性糖提取液。
淀粉含量的提取:采用高氯酸法,用上述离心后的沉淀物测定样品中的淀粉质量。沉淀物加入体积分数30%的高氯酸10 mL, 静置过夜, 然后于80 ℃水浴中继续准确提取10 min, 冷却, 4 000 r/min离心10 min, 取上清液定容到50 mL的容量瓶中。
1.4 數据分析
NSC各组分质量分数等于NSC各组分质量除以总干重。利用变异系数(CV)=(标准偏差/平均值)×100%,比较NSC各组分质量分数在树种、器官和间伐强度3个水平的变异程度(变异系数(CV)小于25%为弱变异,25%~75%为中等强度变异,大于75%为强变异)。采用Excel 2010与Orign 9进行数据的整理、分析以及图表的绘制,用SPSS 17.0进行方差分析和Duncan多重比较进行差异显著性检验(α= 0. 05)。
2 结果与分析
2.1 不同树种、器官和间伐强度下的NSC各组分质量分数差异
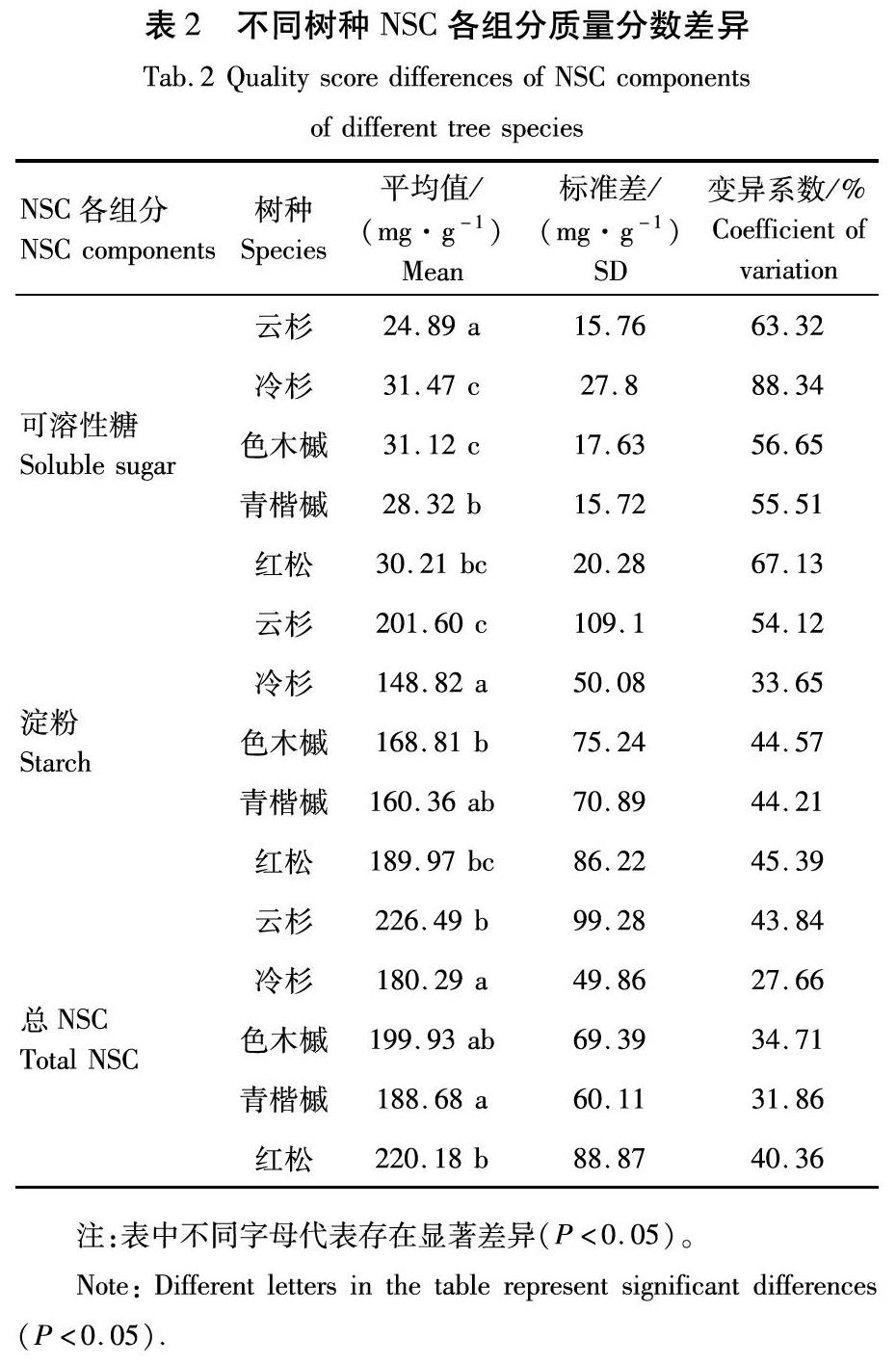
不同树种的NSC质量分数上的差异体现出树木对生存环境的适应情况的差别,表2显示不同树种间NSC各组分质量分数差异显著(P<0.05)。可溶性糖质量分数较高的树种为冷杉与色木槭,云杉可溶性糖质量分数最低。云杉的淀粉质量分数显著高于其他树种,冷杉的淀粉质量分数最低。总NSC质量分数较高的树种为云杉和红松,冷杉和青楷槭总NSC质量分数较低。
不同树木器官中NSC质量分数的差异反映出器官功能和生存策略的区别,表3显示不同器官中NSC质量分数差异显著(P<0.05)。树叶可溶性糖质量分数显著高于树枝、树干,是树枝、树干的2.5~5.0倍。树干淀粉质量分数最高,树叶淀粉质量分数次之,树枝淀粉质量分数最低。总NSC质量分数梯度与淀粉一致,由高到低顺序为:树干、树叶、树枝。
不同的间伐强度对林分NSC质量分数亦有影响,表4显示不同间伐强度下林分NSC质量分数差异显著(P<0.05)。可溶性糖质量分数为23.32 ~32.19 mg/g,0%间伐强度时最高,15%间伐强度样地的可溶性糖质量分数显著低于其他间伐强度样地。20%间伐强度样地淀粉质量分数最高,但与25%间伐强度样地差异不显著;0%间伐强度时最低。20%、25%间伐强度样地中的总NSC质量分数较高, 0%与35%间伐强度样地比较低。
2.2 不同树种、器官以及间伐强度下的NSC各组分变异特征
变异系数表示树木NSC各组分质量分数在不同樹种、器官以及间伐强度下的变异特征, 变异系数越大, 表型性状的离散程度越大。由表2—表4可知,各NSC组分质量分数的变异系数范围为4.75%~88.34%,不同NSC组分质量分数的变异程度不同,其中可溶性糖质量分数的变异程度最大,平均变异系数为60.78%,淀粉质量分数次之(40.18%),总NSC质量分数变化程度相对较小,为32.01%;不同树种、器官以及间伐强度下NSC各组分质量分数的变异程度亦不同,其中树种间和不同间伐强度间的变异程度较大,平均变异系数分别为48.75%和54.05%;器官间的变异程度最小,平均变异系数为17.48%。
表2显示所有NSC组分在不同树种中的变异均较高。可溶性糖在5种树种中的变异均超过55%,达到中等程度变异水平,且冷杉达到强变异水平,变异系数高达88.34%;淀粉质量分数在5种树种的变异为33.65%~54.12%,属于中等程度变异;各树种总NSC质量分数变异均属于中等程度变异,变异系数在27.66%~43.84%波动。不同器官NSC组分变异相对较低,NSC组分变异系数在4.75%~31.71%波动(表3)。树枝可溶性糖变异系数为24.08,属于弱变异,树干、树叶可溶性糖变异系数为31.37%和26.85%,属于中等程度变异;不同器官淀粉与总NSC质量分数变异系数均为弱变异。不同间伐强度下,各NSC组分变异系数较高(表4)。可溶性糖质量分数变异系数均高于60%,为中等程度变异,且在30%、35%间伐强度下最高,达到77.74%和80.99%,为强变异水平;不同间伐强度下树木淀粉质量分数变异系数均大于40%,属于中等程度变异;总NSC质量分数变异在35.14%~42.08%波动,亦达到中等程度变异水平。
2.3 NSC各组分质量分数的变异分解
上述结果表明,树种、器官、间伐强度及其相互作用对NSC各组分质量分数均有不同程度的影响,故对NSC各组分质量分数的影响因素进行变异来源分析(表5)。变异来源分析可以确定各因素对NSC各组分质量分数的影响程度。可溶性糖质量分数的差异主要来源于树木器官的影响,其均值偏差平方和(SS)高达18 786.6,显著高于其他影响因素;其次的变异来源为树种和器官的交互作用、器官和间伐强度的交互作用以及间伐强度的影响;树种对可溶性糖质量分数差异的影响最小;树种和间伐强度的交互作用对可溶性糖质量分数影响不显著(P>0.05)。淀粉和总NSC质量分数的差异主要来源于树木器官,其均值偏差平方和(SS)分别为298 940、247 864,其次的变异来源为树种和器官的交互作用、树种的影响,间伐强度对淀粉和总NSC质量分数差异的影响最小,树种和间伐强度的交互作用以及器官和间伐强度的交互作用对淀粉和总NSC质量分数影响不显著(P>0.05)。综上,器官是影响NSC各组分质量分数的主要因素;树种与间伐强度对NSC各组分质量分数影响相对较小,但依然影响显著。
3 讨论
3.1 不同树种间NSC各组分质量分数的差异
NSC各组分质量分数受树种的影响极为显著,不同树种间可溶性糖(F=9.86, P=0.001)、淀粉(F=64.38, P=0)以及总NSC质量分数(F =37.48, P =0)均存在显著性差异(表5)。刘万德等[16]的研究显示,在相同的生存环境下,器官对NSC各组分的影响可能会增大, 树种之间的差异可能会减小, 但树种对NSC各组分仍影响显著。上官淮亮等[17]对干旱林线区不同树种非结构性碳水化合物的研究显示,不同树种NSC质量分数差异显著。成方妍等[18]和欧阳明等[19]的研究也显示,不同树种相同器官的NSC质量分数存在显著性差异,树种是变异的主要因素之一。树种间NSC各组分的质量分数的差异显著,这是由于不同树种对生态环境的适应和自身生存策略存在差异所导致的,与于丽敏等[20]的研究结果一致。
3.2 不同器官间 NSC各组分质量分数的差异
不同器官NSC各组分质量分数之间存在显著差异,这是由器官的功能所决定的。树木叶片中的可溶性糖质量分数最高,树干中的可溶性糖质量分数最低;树干是淀粉与总NSC的主要的存储器官,淀粉与总NSC质量分数在树干中最高,树枝中最低,充分反映了不同树木器官功能上的差异,与王逸然等[21]的研究结果一致。叶片是树木主要的“碳源”,其NSC质量分数主要取决于可溶性糖质量分数。树干是树木主要的“碳汇”,是主要的碳储存器官,NSC以淀粉的形式大量储存, 所以表现出树干淀粉质量分数最高[22]。
NSC各组分在不同器官中变异较低,变异系数均低于35%,但标准差较大,说明NSC各组分质量分数波动范围广,受器官影响大(表3)。 表5中的变异来源分析的结果亦表明树木器官的差异是影响淀粉质量分数的主要因素。
3.3 不同间伐强度下NSC各组分质量分数的差异
不同间伐强度下,可溶性糖(F=4.03,P= 0.011)、淀粉(F=3.59,P=0.018)、总NSC质量分数(F=2.84,P=0.043)均存在显著性差异(表5)。除受树种与器官的影响,NSC质量分数还受光照、水分和土壤有机质(主要是C、N)等因素的影响[12]。间伐活动伐除了部分林木,改变了林地透光率,残留的凋落物改变了树木养分的供给,以及间伐后土壤的理化性质的变化,从而对NSC质量分数造成影响[23]。
本研究结果发现可溶性糖质量分数在0~15%间伐强度时显著下降,在>15%~25%间伐强度时逐渐上升,大于25%间伐强度时逐渐下降;淀粉和总NSC质量分数随着间伐强度的增大而呈现出先上升后下降的趋势。间伐活动会对林地的生态环境造成破坏,影响树木的生理活动,使可溶性糖质量分数下降。随着间伐强度的增大,林地的透光率得到提高,增强了树木的光合作用从而有利于NSC的恢复,这与王建平等[24]、于亦彤[25]的研究一致。抚育间伐活动残留的枯枝落叶显著增加了土壤有机质的C、N质量分数,枯枝落叶的腐烂亦增加了林内CO2的体积分数,李斌[26]、杜启燃等[27]的研究结果表示C、N供给的增加使得林木可以更好地積累NSC。而当间伐强度继续增大,植物的蒸腾作用随光照的强度的增强而增大,当光照强度继续增强时,植物为降低蒸腾速率而关闭气孔,降低了净光合速率,从而对林木NSC的积累起到抑制作用,这与赵丽英等[28]的研究一致。郝亚涵等[29]与李浩铭等[30]的研究亦表明光照强度过大会使植物光合作用下降,钱杨等[31]亦在研究中指出在高CO2体积分数下随着处理时间的延长,呼吸作用持续增大,光合能力逐渐下降,从而消耗已积累的碳水化合物。刘忠玲等[32]的研究表明间伐强度过大会显著降低土壤最大持水量、最小持水量、毛管持水量、总孔隙度和速效P质量分数,过度间伐会显著降低土壤水分与孔隙状况以及土壤化学性质各指标,造成NSC质量分数下降。光照强度过大、CO2体积分数过高以及土壤水分及土壤养分下降等多个因素导致高间伐强度样地的NSC质量分数较低。
4 结论
小兴安岭针阔混交天然次生林主要树种可溶性糖平均质量分数为29.16 mg/g, 淀粉平均质量分数为173.08 mg/g,总NSC平均质量分数为202.24 mg/g。NSC各组分质量分数在不同树种之间差异极为显著(P<0.01), 对生境不同的适应方式和生存策略是造成差异的主要原因。NSC各组分质量分数在不同树种中的变异性较大,平均变异系数接近50%。器官是NSC各组分质量分数的主要影响因素,对可溶性糖、淀粉以及总NSC质量分数的影响均极为显著(P<0.01)。其中可溶性糖质量分数最高的器官是叶片, 淀粉和总NSC质量分数最高的器官为树干,不同树木器官功能上的区别造成了NSC各组分质量分数在不同器官之间的差异。间伐强度也是NSC各组分的重要因素,对NSC各组分影响显著(P<0.05)。
【参 考 文 献】
[1]潘庆民,韩兴国,白永飞,等.植物非结构性贮藏碳水化合物的生理生态学研究进展[J].植物学通报,2002,37(1):30-38.
PAN Q M, HAN X G, BAI Y F, et al. Advances in physiology and ecology studies on stored non-structure carbohydrates in plants[J]. Chinese Bulletin of Botany, 2002, 37(1): 30-38.
[2]张海燕,王传宽,王兴昌.温带12个树种新老树枝非结构性碳水化合物浓度比较[J].生态学报,2013,33(18):5675-5685.
ZHANG H Y, WANG C K, WANG X C. Comparison of concentrations of non-structural carbohydrates between new twigs and old branches for 12 temperate species[J]. Acta Ecologica Sinica, 2013, 33(18): 5675-5685.
[3]王文娜,李俊楠,王会仁,等.不同树种叶片非结构性碳水化合物季节动态比较[J].东北林业大学学报,2014,42(4):46-49.
WANG W N, LI J N, WANG H R, et al. Seasonal dynamics of leaf nonstructural carbohydrate content in four temperate tree species[J]. Journal of Northeast Forestry University, 2014, 42(4): 46-49.
[4]FAJARDO A, PIPER F I. An experimental approach to explain the southern Andes elevational treeline[J]. American Journal of Botany, 2014, 101(5): 788-795.
[5]杜建会,邵佳怡,李升发,等.树木非结构性碳水化合物含量多时空尺度变化特征及其影响因素研究进展[J].应用生态学报,2020,31(4):1378-1388.
DU J H, SHAO J Y, LI S F, et al. Non-structural carbohydrate content of trees and its influencing factors at multiple spatial-temporal scales: A review[J]. Chinese Journal of Applied Ecology, 2020, 31(4):1378-1388.
[6]DICKSON R E. Carbon and nitrogen allocation in trees[J]. Annals of Forest Science, 1989, 46(Supplement): 631-647.
[7]王曉雨,王守乐,唐杨,等.长白山阔叶红松林3个主要树种的非结构性碳储存特征[J].应用生态学报,2019,30(5):1608-1614.
WANG X Y, WANG S L, TANG Y, et al. Characteristics of non-structural carbohydrate reserves of three dominant tree species in broadleaved Korean pine forest in Changbai Mountain, China[J]. Chinese Journal of Applied Ecology, 2019, 30(5): 1608-1614.
[8]马履一,李春义,王希群,等.不同强度间伐对北京山区油松生长及其林下植物多样性的影响[J].林业科学,2007,43(5):1-9.
MA L Y, LI C Y, WANG X Q, et al. Effects of thinning on the growth and the diversity of undergrowth of Pinus tabulaeformis plantation in Beijing mountainous areas[J]. Scientia Silvae Sinicae, 2007, 43(5): 1-9.
[9]刘思泽,尹海锋,沈逸,等.间伐强度对马尾松人工林间伐初期林下植被群落物种组成和多样性的影响[J].应用生态学报,2020,31(9):2866-2874.
LIU S Z, YIN H F, SHEN Y, et al. Effects of thinning intensity on species composition and diversity of undergrowth vegetation community in Pinus massoniana plantation at initial stage of thinning[J]. Chinese Journal of Applied Ecology, 2020, 31(9): 2866-2874.
[10]张甜.抚育间伐对小兴安岭天然针阔混交林生态功能的影响[D].哈尔滨:东北林业大学,2019.
ZHANG T. Effect of thinning on the ecological function of natural mixed broadleaf-conifer secondary forest in Xiaoxingan mountains[D]. Harbin: Northeast Forestry University, 2019.
[11]王杰,顾凤歧.间伐强度对土地肥力影响的模糊综合评判[J].森林工程,2020,36(1):25-32.
WANG J, GU F Q. Fuzzy comprehensive evaluation of the influence of thinning intensity on soil fertility[J]. Forest Engineering, 2020, 36(1):25-32.
[12]郑云普,王贺新,娄鑫,等.木本植物非结构性碳水化合物变化及其影响因子研究进展[J].应用生态学报,2014,25(4):1188-1196.
ZHENG Y P, WANG H X, LOU X, et al. Changes of non-structural carbohydrates and its impact factors in trees: a review[J]. Chinese Journal of Applied Ecology, 2014, 25(4): 1188-1196.
[13]程文生,冯仲科,于景鑫.中国主要树种通用二元材积模型与推导形数模型研究[J].农业机械学报,2017,48(3):245-252.
CHENG W S, FENG Z K, YU J X. Development of generic standard volume model and derived form factor model for major tree species in China[J]. Transactions of the Chinese Society for Agricultural Machinery, 2017, 48(3): 245-252.
[14]HOCH G, RICHTER A, KORNER C. Non-structural carbon compounds in temperate forest trees[J]. Plant, Cell & Environment, 2003, 26(7): 1067-1081.
[15]HOCH G, POPP M, KORNER C. Altitudinal increase of mobile carbon pools in Pinus cembra suggests sink limitation of growth at the Swiss treeline[J]. Oikos, 2002, 98(3): 361-374.
[16]刘万德,苏建荣,李帅锋,等.云南普洱季风常绿阔叶林主要树种非结构性碳水化合物变异分析[J].林业科学,2017,53(6):1-9.
LIU W D, SU J R, LI S F, et al. Variation of non-structural carbohydrates for the dominant species in a monsoon broad-leaved evergreen forest in PuEr, Yunnan Province[J]. Scientia Silvae Sinicae, 2017, 53(6): 1-9.
[17]上官淮亮,劉鸿雁,胡国铮,等.干旱林线区不同树种非结构性碳水化合物的季节格局及其主导因子[J].北京大学学报(自然科学版),2019,55(3):553-560.
SHANGGUAN H L, LIU H Y, HU G Z, et al. Seasonal patterns and their determinants of non-structural carbohydrates in different tree species at xeric timberline[J]. Acta Scientiarum Naturalium Universitatis Pekinensis, 2019, 55(3): 553-560.
[18]成方妍,王传宽.树种和组织对树干非结构性碳水化合物储量估测的影响[J].林业科学,2016,52(2):1-9.
CHENG F Y, WANG C K. Impacts of tree species and tissue on estimation of nonstructural carbohydrates storage in trunk[J]. Scientia Silvae Sinicae, 2016, 52(2): 1-9.
[19]欧阳明,杨清培,祁红艳,等.亚热带落叶与常绿园林树种非结构性碳水化合物的季节动态比较[J].南京林业大学学报(自然科学版),2014,38(2):105-110.
OUYANG M, YANG Q P, QI H Y, et al. A comparison of seasonal dynamics of nonstructural carbohydrates for deciduous and evergreen landscape trees in subtropical region, China[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2014, 38(2): 105-110.
[20]于丽敏,王传宽,王兴昌.三种温带树种非结构性碳水化合物的分配[J].植物生态学报,2011,35(12):1245-1255.
YU L M, WANG C K, WANG X C. Allocation of nonstructural carbohydrates for three temperate tree species in Northeast China[J]. Chinese Journal of Plant Ecology, 2011, 35(12): 1245-1255.
[21]王逸然,郑成洋,曾发旭.内蒙古白音敖包沙地云杉生长季非结构性碳水化合物含量动态[J].北京大学学报(自然科学版),2016,52(5):967-976.
WANG Y R, ZHENG C Y, ZENG F X. Seasonal dynamic changes of non-structural carbohydrate in tissues of Picea mongolica in Baiyinaobao[J]. Acta Scientiarum Naturalium Universitatis Pekinensis, 2016, 52(5): 967-976.
[22]SALA A N, MENCUCCINI M. Plump trees win under drought[J]. Nature Climate Change, 2014, 4(8): 666-667.
[23]陈秀波,段文标,陈立新,等. 小兴安岭3种原始红松混交林土壤nirK型反硝化微生物群落特征[J]. 南京林业大学学报(自然科学版), 2021, 45(2): 77-86.
CHEN X B, DUAN W B, CHEN L X, et al. Community structure and diversity of soil nirK-type denitrifying microorganisms in three forest types of primitive Pinus koraiensis mixed forest in Liangshui National Nature Reserve, Lesser Khingan Mountains[J].Journal of Nanjing Forestry University (Natural Science Edition), 2021, 45(2): 77-86.DOI: 10.12302/j.issn.1000-2006.201902033.
[24]王建平,王纪章,周静,等.光照对农林植物生长影响及人工补光技术研究进展[J].南京林业大学学报(自然科学版),2020,44(1):215-222.
WANG J P, WANG J Z, ZHOU J, et al. Recent progress of artificial lighting technique and effect of light on plant growth[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2020, 44(1): 215-222.
[25]于亦彤.基于择伐的云冷杉天然次生林结构动态和优化模型的研究[D].北京:北京林业大学,2019.
YU Y T. Study on forest structure of different recovery stages and optimization models of natural mixed spruce-fir secondary forests on selective cutting[D]. Beijing: Beijing Forestry University, 2019.
[26]李斌.华北落叶松人工林土壤呼吸对间伐强度的响应[D].北京:北京林业大学,2019.
LI B. Response of soil respiration in larch plantations in North China to thinning intensity[D]. Beijing: Beijing Forestry University, 2019.
[27]杜启燃,雷静品,刘建锋,等.CO2浓度增加和施氮对栓皮栎幼苗生理生态特征的影响[J].应用生态学报,2014,25(1):24-30.
DU Q R, LEI J P, LIU J F, et al. Eco-physiological response of Quercus variabilis seedlings to increased atmospheric CO2 and N supply[J]. Chinese Journal of Applied Ecology, 2014, 25(1): 24-30.
[28]赵丽英,邓西平,山仑.活性氧清除系统对干旱胁迫的响应机制[J].西北植物学报,2005,25(2):413-418.
ZHAO L Y, DENG X P, SHAN L. The response mechanism of active oxygen species removing system to drought stress[J]. Acta Botanica Boreali-Occidentalia Sinica, 2005, 25(2): 413-418.
[29]郝亞涵,方如萍,王冉,等.光照强度和施氮量对白芨幼苗生长和氮含量的影响[J].西南林业大学学报(自然科学),2021,41(6):1-8.
HAO Y H, FANG R P, WANG R, et al. Effects of light intensity and nitrogen application on growth and nitrogen content of Bletilla striata seedlings[J]. Journal of Southwest Forestry University (Natural Science), 2021, 41(6): 1-8.
[30]李浩铭, 余著成, 陈卓, 等. 光照强度对伯乐树幼苗生长及相关生理指标的影响[J]. 西南林业大学学报(自然科学), 2021, 41(3): 23-30.
LI H M, YU Z C, CHEN Z, et al. Effects of light intensity on the growth and related physiological indexes of Bretschneidera sinensis[J]. Journal of Southwest Forestry University (Natural Science), 2021, 41(3): 23-30.
[31]钱杨,孙洪刚,董汝湘,等.针叶树碳水化合物分配研究进展[J].林业科学,2018,54(1):141-153.
QIAN Y, SUN H G, DONG R X, et al. Research progress of carbohydrates allocation in conifers[J]. Scientia Silvae Sinicae, 2018, 54(1): 141-153.
[32]刘忠玲,姚颖,刘建明,等.抚育间伐对白桦天然次生林枯落物与土壤持水特性的影响[J].森林工程,2020,36(4):29-37.
LIU Z L, YAO Y, LIU J M, et al. Effects of thinning on water-holding characteristics of litter and soil in Betula platyphylla natural secondary forest stand[J]. Forest Engineering, 2020, 36(4): 29-37.
猜你喜欢
中国食品(2022年3期)2022-02-25
阅读(科学探秘)(2021年11期)2021-03-10
祝您健康·文摘版(2021年3期)2021-03-09
现代农业科技(2020年12期)2020-07-06
森林工程(2018年5期)2018-05-14
饮食与健康·下旬刊(2018年3期)2018-04-11
学校教育研究(2017年28期)2017-10-21
食品界(2017年4期)2017-05-17
科技视界(2016年9期)2016-04-26
中学化学(2014年11期)2015-01-20
