
南太行八里沟景区草本植物多样性对海拔梯度的响应
2021-07-19 09:36王威唐蛟靳沙沙王伟李胜杰吴大付
河南科技学院学报(自然科学版) 2021年4期
王威,唐蛟,靳沙沙,王伟,李胜杰,吴大付
(河南科技学院资源与环境学院,河南新乡453003)
生物多样性是维持地球生态系统结构和功能稳定的基础,主要包括遗传多样性、物种多样性、生态系统多样性、以及景观多样性[1-3].进行生物多样性恢复是进行受损或者退化生态系统恢复过程的首要目标.物种多样性是生物多样性的重要组成部分,体现了群落的结构类型、组织水平、发育阶段、稳定程度和生境差异,也反映出群落结构和功能方面的动态异质性,为生态系统功能的运行和维持提供必要基础和有利支撑[3-4].由于物种多样性既是生态系统多样性的基础,又是遗传多样性的载体,因此保护物种多样性是最关键和最基本的保护.植物是生产者,是食物链的开端,保护植物物种多样性在保护生物多向性中起关键作用.
物种多样性在地球表面的丰富程度因地而异,其分布格局受环境因素(经纬度、海拔、水分、土壤、人类活动等)的影响[5-7].植被的物种组成和物种多样性随海拔梯度的变化格局反映了群落环境的变化以及植物群落与海拔梯度的关系[6].山地植物群落多样性随海拔梯度的变化规律一直是植物群落生态学研究的重点问题之一[5].海拔梯度的变化反映在温度、湿度、光照等生态因子的变化上,局部区域小气候的不同,也会导致山地群落多样性在海拔高度格局分布的差异性.多样性随环境因子发生变化,其中海拔作为一个重要的生态因子,对于物种多样性产生重要影响.目前在全世界范围内已经开展诸多针对海拔变化与植物多样性关系的相关研究工作,例如在云南省石葵河生态环境脆弱区进行的植物调查发现植物物种多样性整体上随海拔的升高而降低[8];然而在青藏高原高山草甸地区的研究却证明物种多样性随海拔的升高表现为先增加后降低的趋势[9];马云锡等在西藏地区果园昆虫种类及多样性调查发现,昆虫群落均匀度、群落物种多样性、群落相似性随海拔的升高而降低,而优势集中度则随海拔升高而增大[10].同时也有一些地区的植物物种多样性表现为随海拔的升高先降低后升高,在中海拔附近区域较低的特点[11].海拔梯度变化会引起森林物种垂直分布的不同,进而影响草本植被的物种多样性.
草本植物在维持森林生态系统的物质循环方面起着非常重要的作用[4],生态系统的能量流动、物质循环以及生产力的高低都与草本植物的种类、数量、分布等息息相关[12].草本植物具有很多用处,所有重要的粮食作物都是草本植物,如小麦、粟米、玉米、大麦、高粱等,是人类食物的重要来源;草本植物还是自然界很多动物的食物,以及猪牛马羊等各类家畜也都吃草本植物,所以在食物链中草本植物是重要的组成部分;还有一些草本植物在日常生活中随处可见,它们可以用来编制和制作用具,东南亚人还用竹子造房屋;大自然中的许多草本植物不仅是动物的食物,还能产生大量氧气,防风固沙,防止水土流失;也有很多草本植物是中药材的重要来源,比如地黄、香附子、艾草等.因此,对林下草本植物群落多样性的研究,有助于更好地认识与了解群落的组成、结构、功能、演替动态和稳定性[12].太行山处于中国第二阶梯东沿,东临华北平原,西接黄土高原,是中国东部地区的重要山脉之一,也是一条东部地区重要的地理分界线.太行山经历多期次的地质变动过程,多样的地形和巨大的海拔高差形成了丰富多样的区域环境条件,形成了丰富的动植物资源,具有多样性的生态系统和较高的生物多样性,是《中国生物多样性保护战略与行动计划(2011-2030年)》确定的35个中国生物多样性优先保护区域之一[13-14].然而,由于人类活动的逐年加剧以及不合理的开发利用自然资源,已经对区域物种多样性造成不同程度上的影响.因此,研究调查太行山物种多样性的分布格局,分析其内在形成机制和影响因素,对于太行山物种多样性保护和管理具有重要的战略意义.新乡南太行山区是太行山脉植物种类丰富,植被覆盖最为密集的区域.本研究以南太行植物生物多样性保护典型区域八里沟景区为研究区域,通过对不同海拔梯度下的草本植物多样性进行调查,针对物种丰富度、多样性指数、均匀度指数开展比较研究,进而了解林下草本植被特征随海拔梯度的变化规律,以期为我国北方太行山植物资源的利用和保护提供参考和依据.
1 研究区概况
南太行八里沟景区(东经113°53′84′,北纬35°59′23′)地处河南省新乡市西北约50 km处,位于太行山南麓的深山之中.该地区是太行山与华北平原的结合区域,是北亚热带向暖温带过渡区域,属于暖温带大陆性季风型气候[11].受山脉走向和海拔高度的影响,季风作用比较明显,春季多风少雨,夏季多雨热,秋季气候凉爽,冬季少雪.年平均气温12℃,年均无霜期214 d,年均日照时间2 020.1 h,年平均降水量589.1 mm.八里沟景区森林覆盖率高,分布有典型的温带落叶阔叶林,生物资源丰富,现存植被类型主要包括中生型植物群落、旱生植被和后期人工植被[11].
2 研究方法
2.1 调查方法
2018年5 月对南太行山八里沟景区典型温带阔叶林林下草本植物进行样方调查,每间隔100 m海拔梯度设置调查样方,海拔梯度分别设置为350、450、550、650、750 m,从不同的海拔梯度内选取5个具有代表性的样方,每个样方的面积为1 m*1 m,采用目视法对样方内所有草本植物的种类、多度、频度、盖度等逐一进行测量并记录.
2.2 计算方法
重要值是研究某个种在群落中的地位和作用的综合数量指标,是相对密度、相对频度、相对优势度的总和,反映了植物群落的结构和功能特征.为了方便在草地植被调查中使用,经典的重要值计算公式需要重新构建.多数情况下,适合草本植被的重要值计算公式为

式(1)中:IV为重要值,RA为相对多度、RH为相对高度、RC为相对盖度,是每个样方中总所调查的植物种类的与总植物个体数目、盖度和高度的比值.
相对盖度=(各物种的盖度/全部物种的盖度之和)×100,
相对高度=(各物种的平均高度/样地内全部物种的总平均高度)×100,
相对多度=(各物种的多度/全部物种的多度之和)×100.
辛普森多样性指数和香农威纳多样性指数包含了群落异质性的测量.辛普森多样性指数表示从一个群落中连续抽样两次获得的个体数属于同一个物种的概率;香农威纳多样性指数包含两个因素:物种数目(即物种丰富度)和不同物种个体分布的平均性或均匀性,被用来描述物种的个体出现的无序程度和不确定性,不确定性越高,多样性也就越高;均匀度指数用来反映群落中物种的个体数量分配的均匀程度[12,19-20].主要计算公式为

香农威纳多样性指数(H):H=-∑Pilnpi,

式(2)~(4)中:S为样方中的物种数目,Ni为样方中某一个物种的个体数,N为群落中所有物种的个体数,Pi=Ni/N.
2.3 数据处理方法
对不同海拔梯度条件下植物多样性指数采用单因素方差分析(ANOVA),并利用Turkey检验法进行多重比较,显著水平设为5%.所有数据统计和分析过程在Excel 2010软件和Origin 2017软件中进行.
3 结果与分析
3.1 草本植物物种组成
研究区域共发现64种草本植物,隶属于30科60属,其中菊科(Asteraceae)的物种数目最多,为14种,占样地所有物种数的21.88%;其次是禾本科(Poaceae),物种数为6种,占样地物种数的9.38%;毛莨科(Ranunculaceae)有4个物种,占样地物种数的6.25%;十字花科(Brassicaceae)、唇形科(Lamiaceae)、罂粟科(Papaveraceae)有3个物种,各占样地物种数的4.69%;桑科(Moraceae)、堇菜科(Violaceae)、紫草科(Boraginaceae)、天南星科(Araceae)、鸭跖草科(Commelinaceae)、薯蓣科(Dioscoreaceae)、蔷薇科(Rosaceae)均有2个物种,各占样地所有物种数的3.13%;莎草科(Cyperaceae)、壳斗科(Fagaceae)、大戟科(Euphorbiaceae)、虎耳草科(Saxifragaceae)、苦木科(Simaroubaceae)、萝藦科(Asclepiadaceae)、透骨草科(Phrymaceae)、酢浆草科(Oxalidaceae)、旋花科(Convolvulaceae)、玄参科(Scrophulariaceae)、苋科(Amaranthaceae)、中国蕨科(Sinopteridaceae)、荨麻科(Urticaceae)、柳叶菜科(Onagraceae)、豆科(Leguminosae)、百合科(Liliaceae)、葡萄科(Vitaceae)均只有1个物种.在60个属中,只有4个属有2个物种,剩下的56个属只有1个物种.菊科(Asteraceae)和禾本科(Poaceae)占样地所有物种数的比例较大,其他植物属种所占的比例较小,但是从总体来看,不同海拔梯度样地中物种数目具有差异,研究区的草本植物种类丰富,来源广泛.
所研究的草本群落中,仅生长在350 m海拔梯度的物种是苦荬菜(Ixeris polycephala)、鬼针草(Bidens pilosa)、一年蓬(Erigeron annuus)、钻叶紫苑(Aster subulatus)、银胶菊(Parthenium hysterophorus)、小麦(Triticum aestivum)、竹叶子(Streptolirion volubile)、犁头尖(Typhonium divaricatum)、香附子(Cyperus rotundus)、栎树(Quercus),仅生长在450 m海拔梯度的物种是秋英(Cosmos bipinnata)、菊花(Dendranthema morifolium)、小花糖芥(Erysimum cheiranthoides)、紫堇(Corydalis edulis)、虎掌(Pinellia pedatisecta)、虎耳草(Saxifraga stolonifera)、臭椿(Ailanthus altissima),仅生长在550 m海拔梯度的物种是蒲公英(Taraxacum mongolicum)、荠菜(Capsella bursa-pastoris)、大叶铁线莲(Clematis heracleifolia)、太行铁线莲(Clematis kirilowii)、香茶菜(Rabdosia amethystoides)、血见愁(Teucrium viscidum)、马蹄金(Dichondra micrantha)、地 黄(Rehmannia glutinosa)、牛 膝(Achyranthes bidentata)、银 粉 背 蕨(Aleuritopteris argentea),仅生长在650 m海拔梯度的物种是香丝草(Conyza bonariensis)、棒头草(Polypogon fugax Nees ex Steud)、耧斗菜(Aquilegia viridiflora)、荔枝草(Salvia plebeia)、白屈菜(Chelidonium majus)、蝎子草(Girardinia suborbiculata)、露珠草(Circaeacordata),只在750 m海拔梯度的物种是唐松草(Thalictrum aquilegiifolium)、覆盆子(Rubus idaeus)、穿龙薯蓣(Discorea nipponica)、吉祥草(Reineckia carnea)、蛇葡萄(Ampelopsis sinica),在小于500 m的低海拔梯度均有分布的物种是艾草(Artemisia argyi)、茅草(Imperata cylindrica)、葎草(Humulus scandens)、雀儿舌头(Leptopus chinensis),在大于600 m的高海拔梯度均有分布的物种是艾草(Artemisia argyi)、假还阳参(Crepidiastrum lanceolatum)、求米草(Oplismenus undulatifolius)、蛇莓(Duchesnea indica)、葎草(Humulus scandens)、堇菜(Viola verecunda)、紫花地丁(Viola philippica)、透骨草(Phryma leptostachya),在450~750 m海拔梯度上均有生长的物种只有艾草(Artemisia argyi),且物种的种类随海拔的升高呈现先增加后降低的趋势,这与陈斯等的研究结果物种的种类随海拔梯度的上升总体呈下降趋势不同[15].可能原因是研究所选海拔梯度的范围不同,本研究的起始海拔梯度较低,易受人为干扰.

表1 不同海拔高度植物种类分布情况Tab.1 Distribution of plants at different altitudes m

(续表1)

(续表1)
3.2 不同海拔梯度优势种重要值分析
海拔梯度350 m植物群落优势种重要值见表2.
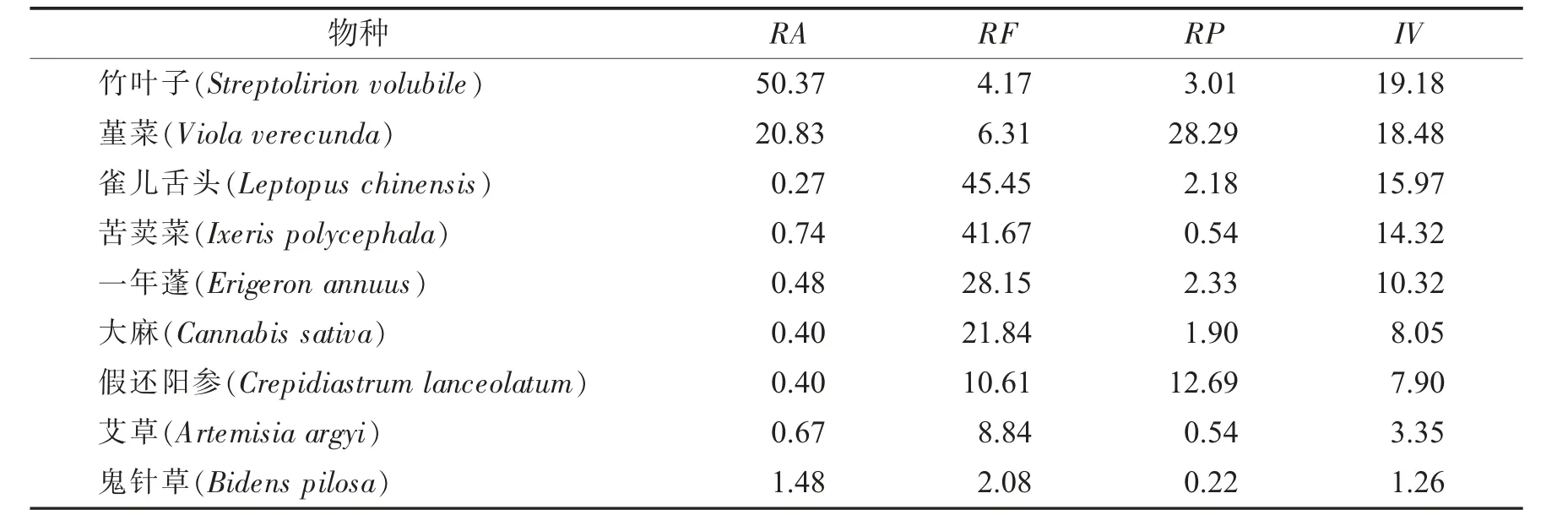
表2 海拔梯度350 m植物群落优势种重要值Tab.2 Important values of dominant species in plant communities with an altitude gradient of 350 m
由表2可知,海拔梯度350 m时,草本群落为主要优势种,重要值最大值为19.18,比最小值1.26大17.92.
海拔梯度450m植物群落优势种重要值见表3.
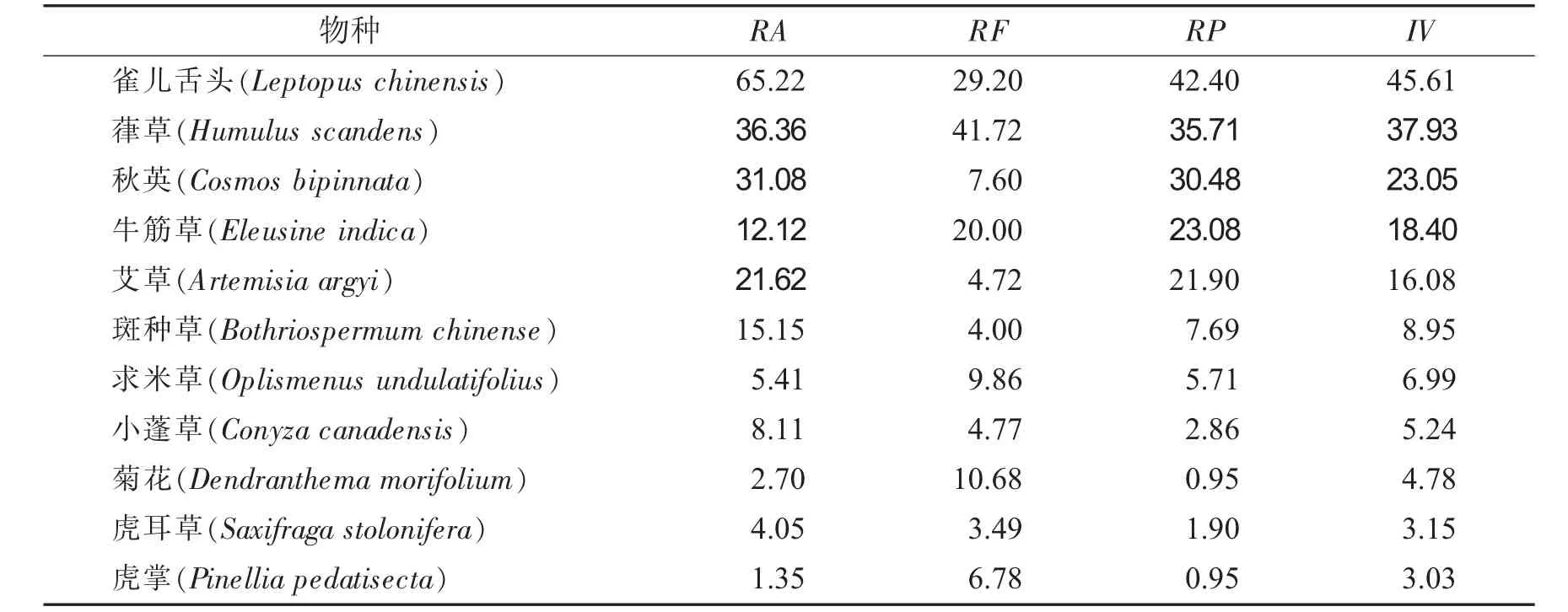
表3 海拔梯度450 m植物群落优势种重要值Tab.3 Important values of dominant species in plant communities with an altitude gradient of 450 m
由表3可知,海拔梯度450 m时,草本群落主要优势种,重要值最大值为45.61,比最小值3.03大42.58.
海拔梯度550 m植物群落优势种重要值见表4.
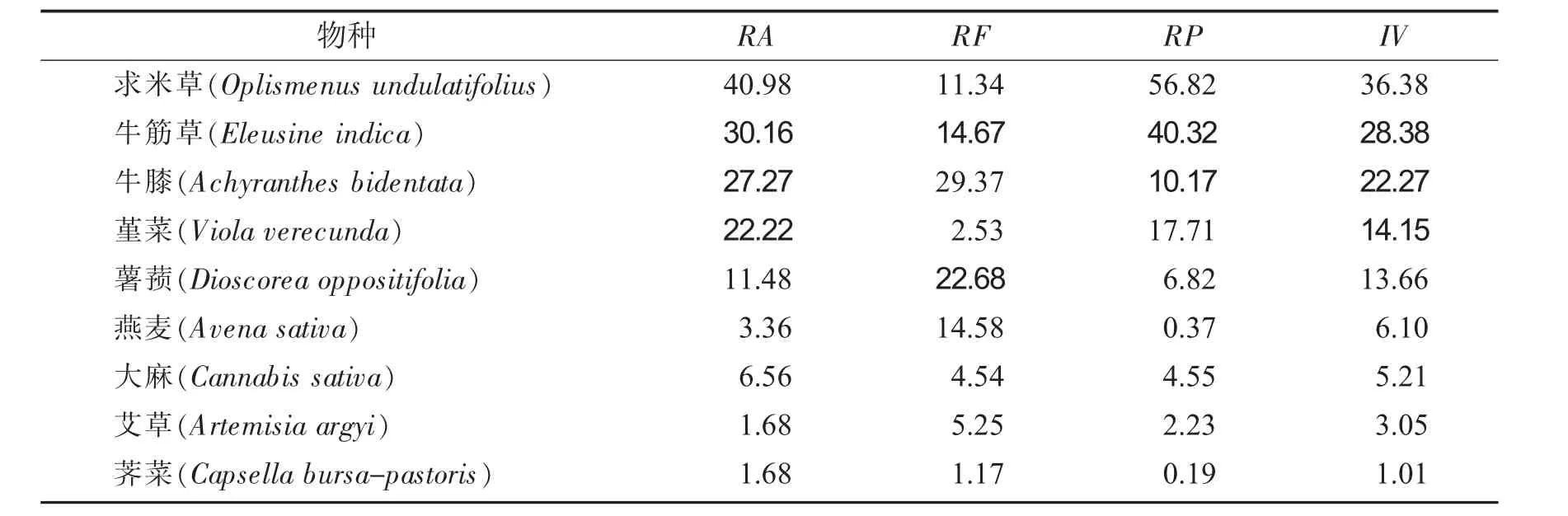
表4 海拔梯度550 m植物群落优势种重要值Tab.4 Important values of dominant species in plant communities with an altitude gradient of 550 m
由表4可知,海拔梯度550 m时,草本群落主要优势种,重要值最大值为36.38,比最小值1.01大35.37.
海拔梯度650 m植物群落优势种重要值见表5.
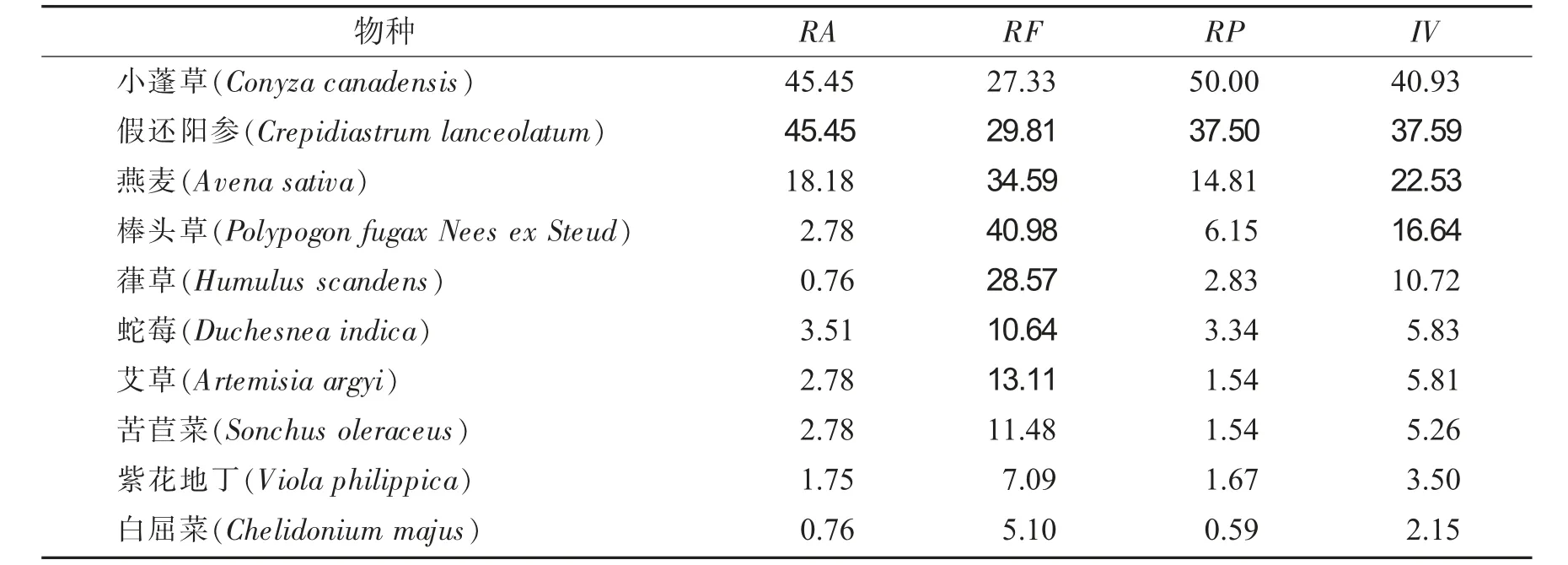
表5 海拔梯度650 m植物群落优势种重要值Tab.5 Important values of dominant species in plant communities with an altitude gradient of 650 m
由表5可知,海拔梯度650 m时,草本群落主要优势种,重要值最大值为40.93,比最小值2.15大38.78.
海拔梯度750 m植物群落优势种重要值见表6.

表6 海拔梯度750 m植物群落优势种重要值Tab.6 Important values of dominant species in plant communities with an altitude gradient of 750 m
由表6可知,海拔梯度750 m时,草本群落主要优势种,重要值最大值为44.30,比最小值3.25大41.05.
由表2~6可知,在不同海拔梯度不仅植物种类不同,各物种的重要值也各不相同.其中艾草(Artemisia argyi)在各海拔梯度均存在,但是在不同海拔梯度的重要值差别很大,在750 m海拔梯度时重要值最大,为26.42.在不同海拔梯度,重要值最大的物种也不同,海拔梯度由低到高的最大重要值物种分别是竹叶子(Streptolirion volubile)、雀儿舌头(Leptopus chinensis)、求米草(Oplismenus undulatifolius)、小蓬草(Conyza canadensis)、透骨草(Phryma leptostachya).
3.3 不同海拔梯度上物种丰富度变化
丰富度代表群落中物种的数量,该研究区内草本植物的物种丰富度随海拔的升高表现为先升高后降低的趋势(图1-A).进行方差分析发现海拔高度对物种丰富度无显著影响(F=0.49<F0.05,P>0.05),各海拔梯度之间物种丰富度不存在显著差异(表7).但是在海拔350 m条件下,草本植物的物种数目最小,为18,物种丰富度为5.8个,主要分布有艾草、鬼针草、竹叶子等;随着海拔的升高,物种丰富度增加,在海拔450 m时物种丰富度达到最大7.4个,物种数目为22个,主要物种有艾草、葎草、斑种草等;物种数目在海拔550 m条件下最多,达到24,主要物种包括堇菜、酢浆草、求米草等;随着海拔的持续升高,物种丰富度逐渐降低,在海拔750 m时物种数下降为19个.
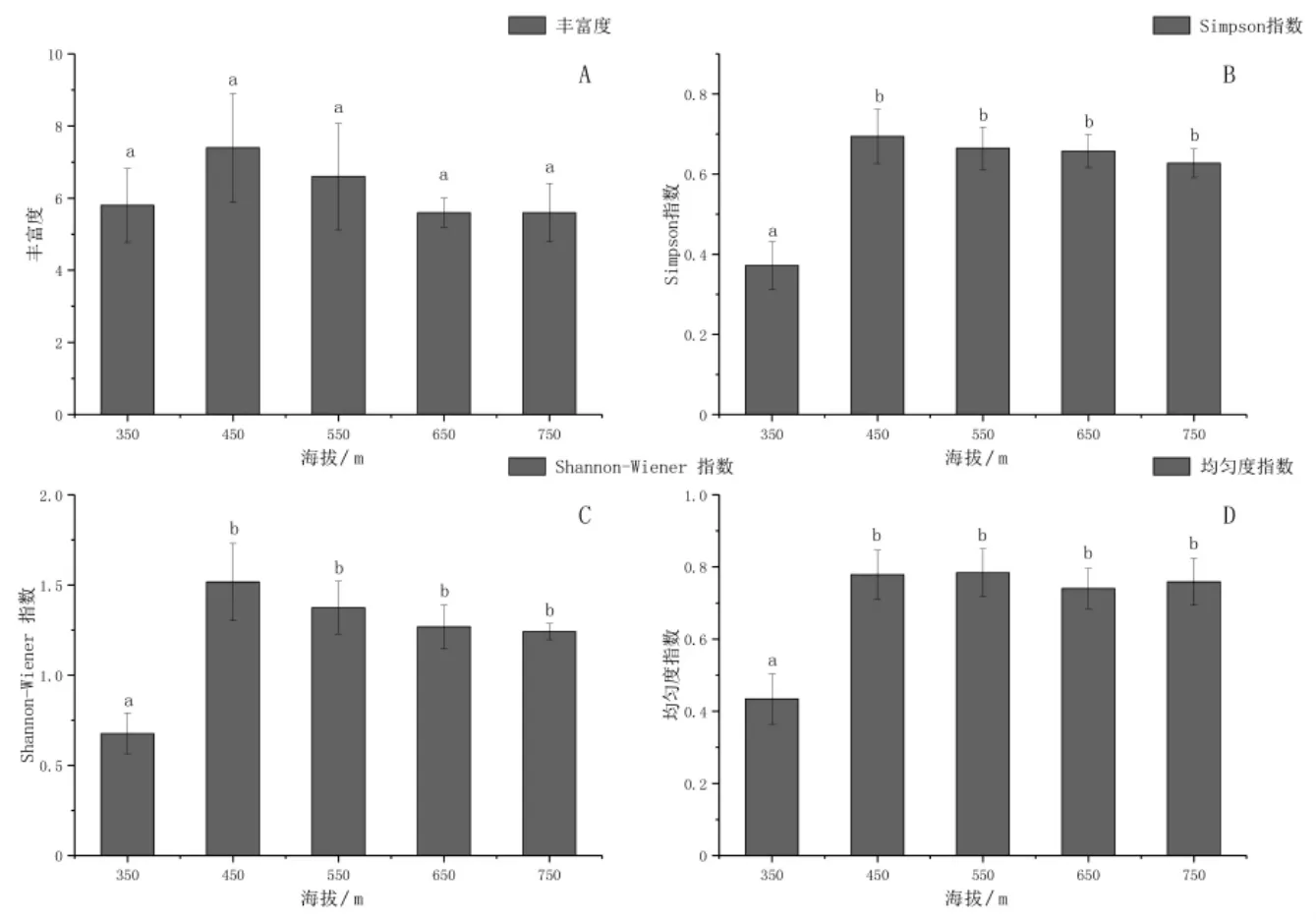
图1 群落物种多样性随海拔梯度变化情况Fig.1 Plant species diversity in response to altitude gradients
3.4 不同海拔梯度上多样性指数变化
物种多样性表示群落中生物种类的丰富程度,主要包括辛普森(Simpson)多样性指数、香农威纳(Shannon-Wiener)多样性指数和均匀度指数.由方差分析结果可知,不同海拔高度对辛普森多样性指数产生显著影响(F=6.2>F0.05,P<0.05),辛普森多样性指数随海拔的升高表现为先升高后降低的趋势(表7).进行多重比较发现海拔高于350 m条件下,各辛普森多样性指数之间无显著差异(P>0.05),维持在0.62~0.69之间.海拔为350 m时,辛普森多样性指数最低,为0.37,并与其余各处理之间存在显著性差异(P<0.05)(图1-B).
不同海拔高度对香农威纳多样性指数产生显著影响(F=5.31>F0.05,P<0.05),香农威纳多样性指数随海拔的升高表现为先升高后降低的趋势(表7).进行多重比较发现海拔高于350 m条件下,各香农威纳多样性指数之间无显著差异(P>0.05),维持在1.24~1.52之间.海拔为350 m时,香农威纳多样性指数最低,为0.68,并与其余各处理之间存在显著性差异(P<0.05)(图1-C).
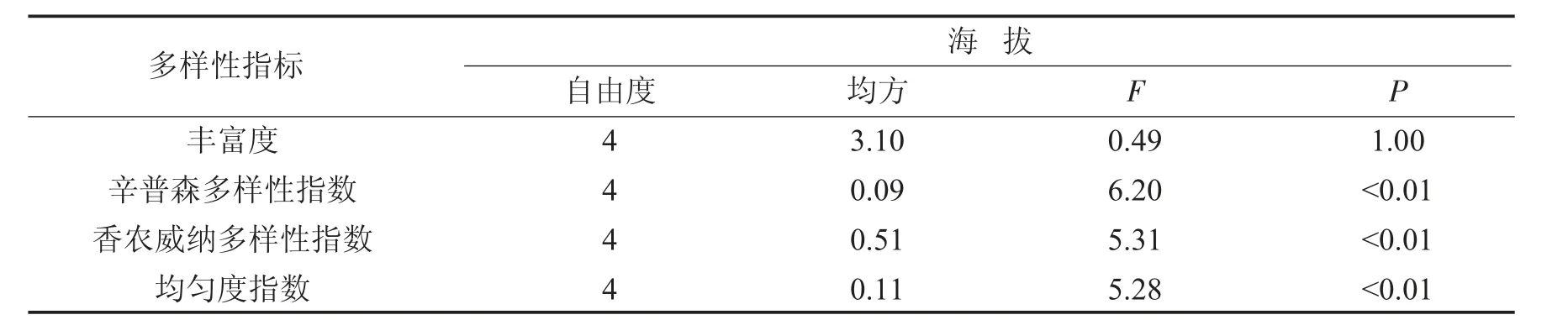
表7 群落物种多样性方差分析Tab.7 Variance analysis of community species diversity
不同海拔高度对均匀度指数产生显著影响(F=5.28>F0.05,P<0.05),均匀度指数随海拔的升高表现为先升高后降低的趋势(表7).进行多重比较发现海拔高于350 m条件下,各均匀度指数之间无显著差异(P>0.05),维持在0.74~0.78之间.海拔为350 m时,均匀度指数最低,为0.35,并与各处理之间存在显著性差异(P<0.05)(图1-D).
4 讨论
海拔一直被认为是影响区域生境差异的主导因子,海拔梯度不同,导致水热条件的空间分布不同,进而形成不同区域植物群落结构[4].本研究发现南太行八里沟景区的草本植物以菊科和禾本科为主,菊科物种数目最多,为15种,占样地所有物种数目的21.43%,其次是禾本科,为9种,占样地物种数目的12.86%,但是其余的绝大部分科只有一个属一个物种,这可能与本地区不同海拔梯度下生境异质性有关,生境异质性使得不同的植物占据不同的生态位,有助于各植物物种共存.同时这也与本地区长时间的封山育林措施有密切关系,通过封山育林降低了人为干扰程度,促进了植物多样性的恢复和保护.
丰富度表示群落中物种数目的多少,植物物种丰富度在一定程度上能够反映出生境条件的差异[6].研究发现,南太行八里沟景区林下草本植物物种丰富度随海拔的升高表现出先升高后降低的特点,但不同海拔梯度物种丰富度的变化范围是5.6~7.4,变化范围不大,方差分析结果也显示海拔对草本植物物种丰富度无显著影响,但各海拔梯度的物种类型大不相同.屈月雷等[8]对云南省东南部文山壮族苗族自治州丘北县境内的石葵河不同海拔梯度的植物多样性分析表明草本植物物种丰富度随海拔的升高表现为先下降后上升的趋势.这是因为在低海拔区域均为郁闭林分,由于光照条件差异较小,所以草本植物主要受海拔升高所导致的温度变化的影响[11].王德君等[16]对甘肃莲花山研究得出在低海拔地区,草本植物群落物种丰富度较低.本次研究得出海拔对物种丰富度没有显著影响,可能是因为本研究的海拔较低,梯度范围设置的较小,光照、温度、湿度等变化不明显,导致对草本植物物种丰富度的影响不显著.
多样性指数是用来衡量生物多样性丰富程度的指标.方差分析结果显示,海拔对辛普森多样性指数、香农威纳多样性指数、均匀度指数均有显著影响,且均随着海拔的升高表现为先升高后下降的特点,香农威纳多样性指数和辛普森多样性指数在海拔450 m左右达到了最大值,均匀度指数在海拔550 m左右达到了最大值.有不同地区的多个研究表明物种多样性随海拔梯度升高呈单峰格局,即随着海拔的升高物种多样性呈现先增加后减少的变化趋势.例如,Acharya、Manish等研究发现受热量、水分、空间因素、生产力、历史因素等影响,喜马拉雅山地区兰科植物及东部地区植物多样性随海拔升高呈单峰格局[17-18];刘哲等[9]对青藏高原高山草甸物种多样性的研究显示物种多样性指数与海拔的关系呈单峰曲线,这与本研究结果一致.有些研究者将干扰视为多样性指数显示单峰曲线的重要原因,认为低海拔梯度受到人类活动的干扰比较严重,导致其物种多样性较低[6];但是有些研究者认为植物多样性格局在一定程度上受能量随海拔梯度变化的影响,其中主要受水热条件限制[11,22-23];同时草本植物的分布及其多样性也受森林群落内部环境差异的影响,如林分结构[19-20]、林分密度[24]、立地条件等[25].
5 结论
在南太行八里沟350~800 m海拔区域植物多样性丰富,共调查到64种草本植物,隶属于30个科60个属;同时物种丰富度、辛普森多样性指数、香农威纳多样性指数、均匀度指数均随海拔的升高表现为先升高后降低的特点.海拔梯度对辛普森多样性指数、香农威纳多样性指数、均匀度指数变化均有显著影响.香农威纳多样性指数、辛普森多样性指数在海拔450 m左右达到了最大值,均匀度指数在海拔550 m左右达到了最大值.
猜你喜欢
做人与处世(2021年16期)2021-10-13
雪豆月读·低年级(2021年7期)2021-08-27
科学之谜(2018年10期)2018-01-02
湖北畜牧兽医(2017年4期)2017-05-18
绿色科技(2017年7期)2017-05-12
新东方英语·中学版(2017年4期)2017-05-04
纺织导报(2014年8期)2014-10-31
故事会(2014年5期)2014-05-14
意林·少年版(2008年9期)2008-09-03
