
红富士芽变类型及芽变品种叶片特征与光合特性
2021-07-05 01:29:46吴泽珍王安丽周文静李文胜
新疆农业科学 2021年6期
吴泽珍,温 玥,胡 真,王安丽,周文静,李文胜
(新疆农业大学林学与园艺学院/新疆农业大学特色果树研究中心,乌鲁木齐 830052)
0 引 言
【研究意义】光合能力是果树产量和品质形成的基础,树体90%以上的干物质都来源于叶片的光合产物[1]。叶片是光合作用的主要场所,叶片长度、叶片宽度、叶片厚度等指标与其光合能力密切相关[2]。叶绿素是叶片光合作用的物质基础,其含量高低也会影响叶片的光合效率[3]。研究叶片特征、光合特性及相关性对优良芽变选种有重要作用。【前人研究进展】苹果是一种高度杂合的多年生木本植物,芽变频率较高[4]。国内外已利用苹果芽变的普遍性和多样性,选育了大量的优良品种[5]。苹果芽变选种已成为苹果优良品种的丰富来源。红富士苹果经过多代持续芽变选育,其生长特性、结果习性、果实着色、果实品质等性状得到有效改良[6]。叶片主要是通过气孔进行蒸腾作用,气孔导度通常是影响净光合速率的初始因素[8]。果树光合作用的主要原料之一是CO2,胞间CO2浓度对净光合速率有直接影响,从侧面反映果树光合能力的强弱[9]。水分利用率是植物光合与蒸腾特性的综合反映,其值大小可以反映植物对逆境适应能力的强弱[10]。利用苹果芽变的重演性,分析芽变的生物学特性,研究优良红富士芽变选种,对优质、晚熟、耐贮的红富士苹果产业的发展有重要意义。【本研究切入点】红富士品种在我国苹果产业中占比高达69.6%,是栽培面积最大的单一品种,但生产上也还存在着色不良、品质不佳等问题[7]。研究红富士芽变叶片的生物学特征。【拟解决的关键问题】优选出的3个易着色短枝红富士芽变类型和2个红富士芽变品种为材料,对比其叶片特征和光合特性,分析相关性,为优良红富士芽变选种提供参考依据。
1 材料与方法
1.1 材 料
试验地位于新疆阿克苏地区红旗坡农场二分场(N 41°17′15″,E 80°17′33″),属暖温带大陆性干旱气候,气候干燥、昼夜温差大、降水量稀少、蒸发量大,年日照时数2 505~3 136 h,平均日照时2 809 h,数全年日照百分率在60%~70%,太阳总辐射量为130~140 KW/ cm2,无霜期为190~251 d,年平均降水80.4 mm,年平均气温10.8℃,光能资源丰富,年平均≥10℃的稳定积温达3 953℃。土壤类型主要为沙壤土。
2015年优选出的3个红富士短枝芽变类型及芽变品种,芽变类型编号为N1、N2和N3,2个红富士芽变品种分别为长富2号和新红1号。供试材料均为20年生,株行距为3 m×5 m。
1.2 方 法
1.2.1 试验设计
每个供试材料选择3株生长状况良好、长势相同且无病虫害的植株。园地管理、环境条件基本一致,果园管理水平良好。
1.2.2 测定指标1.2.2.1 叶片指标
每株树随机选取新梢上100片健康完整的叶片测定叶片形态指标。
叶片长度、宽度:游标卡尺测量。
单叶重:电子秤称重,精确到0.01 g。
叶片厚度:从100个叶片中随机选取30片叠起,游标卡尺测量厚度,重复10次,测算平均叶片厚度。
叶面积:用网格纸(1 cm×1 cm)沿叶片边缘画出叶片图形,统计图形所占的方格数,计算叶面积。
叶绿素:手持SPAD-502叶绿素仪测定叶绿素含量,每片叶重复3次,取平均值。
1.2.2.2 光合特性
2019年7月中旬选择晴朗无风天气,利用Li-6800便携式光合测定系统测定光合指标:净光合速率、气孔导度、胞间CO2浓度、蒸腾速率。每株试验树随机选取树冠外围春梢上长势一致、健康完整的5片叶测定,重复3次,取平均值。测定时间为10:00~12:00,光源为太阳光,温度、相对湿度和CO2浓度为自然状态。
参数日变化:从08:00至20:00,每隔2 h(即08:00、10:00、12:00、14:00、16:00、18:00、20:00)测定,每片叶测3次,取平均值。
测定指标有净光合速率、气孔导度胞间CO2浓度、蒸腾速率。水分利用率=净光合速率/蒸腾速率。
1.3 数据处理
采用Excel 2010整理数据,用SPSS 20.0对数据进行方差分析及相关性分析,叶片、光合特征采用平均值±标准误表示,并利用Microsoft Office Excel 2013制作图表。
2 结果与分析
2.1 5份苹果叶片特征比较
研究表明,苹果叶片长度、叶片宽度、叶片厚度和叶面积的差异不显著,而单叶重、叶绿素含量差异显著。N1与N2单叶重差异显著,与其它3份材料(N3、长富2号和新红1号)差异不显著,其中N1的叶片最重,为1.09 g,N2最轻,为0.78 g;N1与N2叶片的叶绿素含量差异显著,与其它3份材料差异也显著,叶绿素含量由高到低次序为N1>长富2号>新红1号>N3>N2。表1
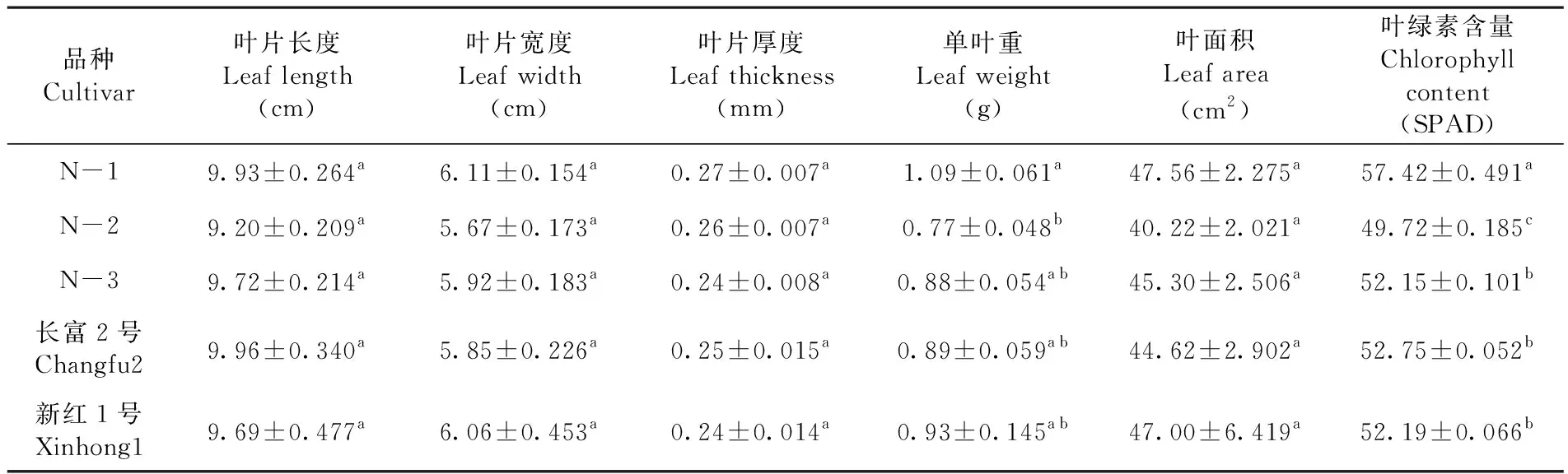
表1 5份苹果叶片特征比较Table 1 The comparison of leaf characteristics of five apple materials
2.2 5份苹果光合特征参数比较
研究表明,光合特征参数差异显著。N1的净光合速率明显高于其它材料,较长富2号和新红1号分别高32.59%和25.31%,其次为N3和N2,长富2号最低;供试材料气孔导度也表现出明显差异,N1叶片的气孔导度最大,为0.21 mol/(m2·s),长富2号的最小,为0.16 mol/(m2·s);胞间CO2浓度N1与N3无显著性差异,但与N2和长富2号差异显著,其中长富2号的胞间CO2浓度最高,为241.77 μmol/(m2·s),N1最低,为224.53 μmol/(m2·s);供试材料的蒸腾速率差异显著,新红1号叶片的蒸腾速率最高,为5.47 mmol/(m2·s),长富2号最低,为3.77 mmol/(m2·s);长富2号与其它材料的水分利用率也表现出显著差异,N2的水分利用率最高,为2.85 μmol/mmol,长富2号最低,为2.07 μmol/mmol。表2
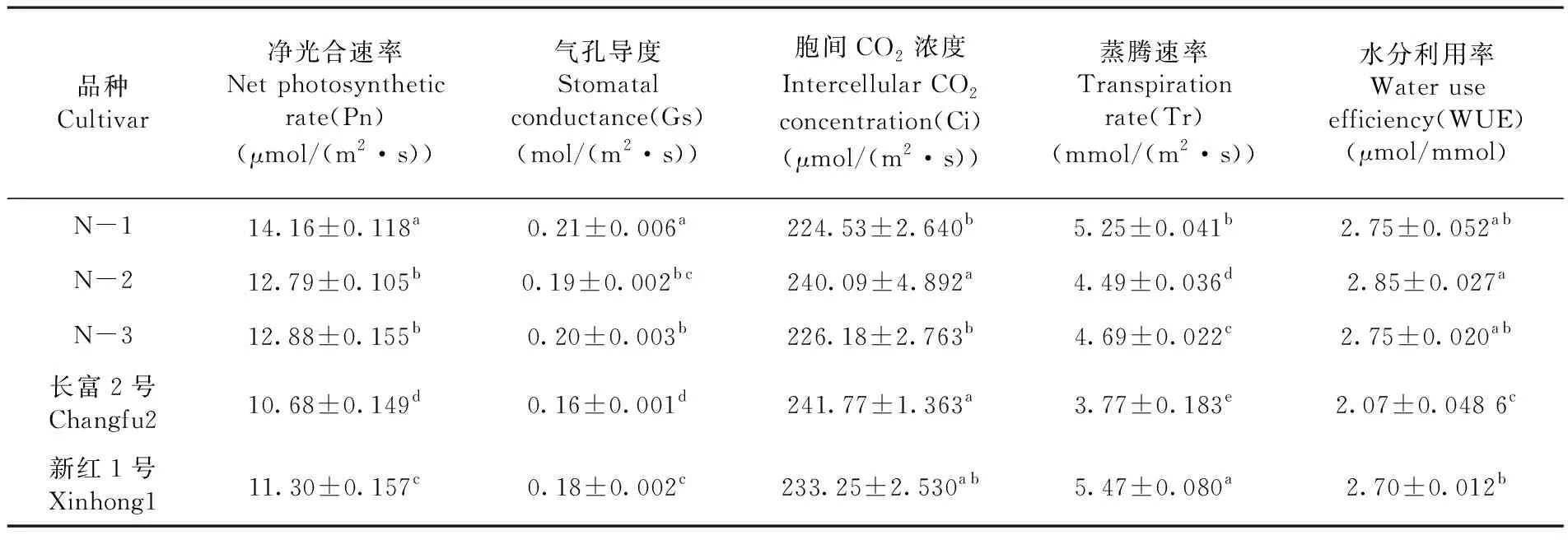
表2 5份苹果光合特征参数Table 2 Photosynthetic characteristic parameters of five apple materials
2.3 5份苹果叶片光合日动态变化
2.3.1 净光合速率日动态变化
研究表明,净光合速率日变化呈“双峰型”曲线,表现出明显的光合“午休”现象,但高峰时间、午休时间和“午休”时的光合速率不同。长富2号和新红1号净光合速率高峰分别出现在12:00和18:00;3个红富士芽变峰值则在12:00和16:00出现。08:00~12:00试材随着PAR和T的升高而逐渐升高,12:00出现第1次高峰,此时N1的净光合速率最高,为14.16 μmol/(m2·s),长富2号最低,为10.68 μmol/(m2·s);之后随着PAR和T的升高开始下降,各材料的光合速率也开始下降,14:00时3个芽变材料出现低谷,表现出光合“午休”现象,而长富2号和新红1号的光合速率则持续下降,到16:00时才出“午休”现象,晚于芽变材料。因温度下降,气孔开放,光合速率上升,光合速率出现次高峰,随后各材料净光合速率迅速降低。图1
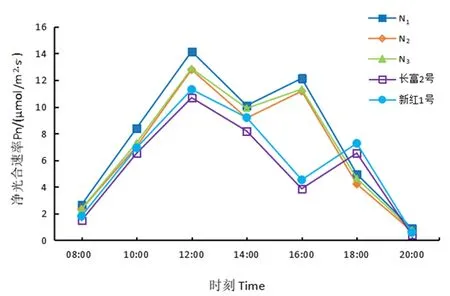
图1 5份苹果Pn日变化Fig.1 Diurnal changes of net photosynthetic rate of five apple materials
2.3.2 气孔导度日动态变化
研究表明,芽变的气孔导度日变化曲线呈“双峰型”,长富2号和新红1号呈“单峰型”。长富2号和新红1号自08:00起气孔导度上升,12:00时达到高峰,其峰值分别为0.16和0.18 mol/(m2·s),因光合有效辐射、气温相对较高,气孔关闭,气孔阻力增大,气孔导度呈下降趋势。3个红富士芽变的气孔导度自08:00起逐渐上升,第1次峰值出现在12:00,此后又开始下降,14:00降至最低,随后又逐渐上升,16:00出现次高峰,然后快速降低,18:00开始缓慢下降。N1的2次峰值最高,分别为0.21和0.18 mol/(m2·s),长富2号的2次峰值最低。图2
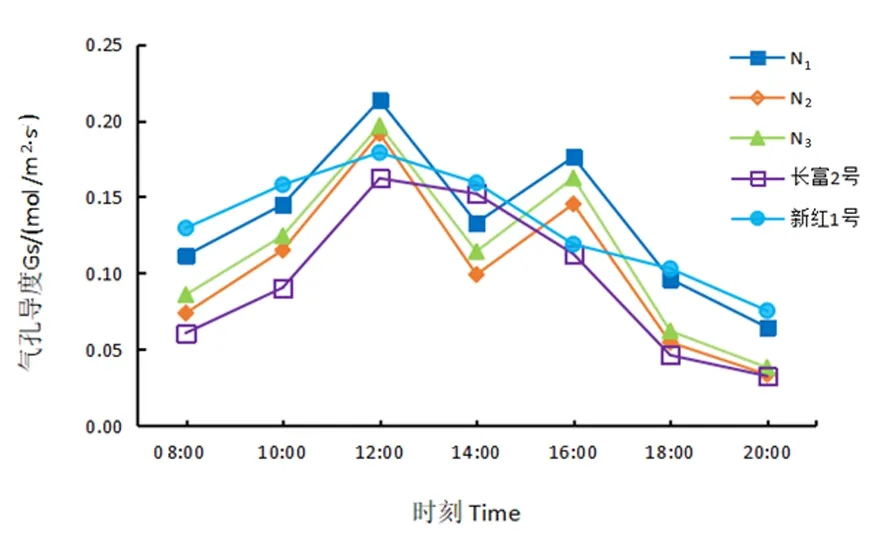
图2 5份苹果Gs日变化Fig.2 Diurnal changes of stalemate stalemat conductance rate of five apple materials
2.3.3 胞间CO2浓度日动态变化
研究表明,胞间CO2浓度日变化规律基本相似,表现出与净光合速率、气孔导度日变化相反趋势,呈“W型”。08:00~12:00胞间CO2浓度随着光合有效辐射和净光合速率的增加呈下降趋势,08:00~10:00下降较缓,12:00时出现第1次低谷,随后逐渐上升,14:00出现高峰,此后逐渐下降,在18:00出现第2次低谷,随后又上升。图3
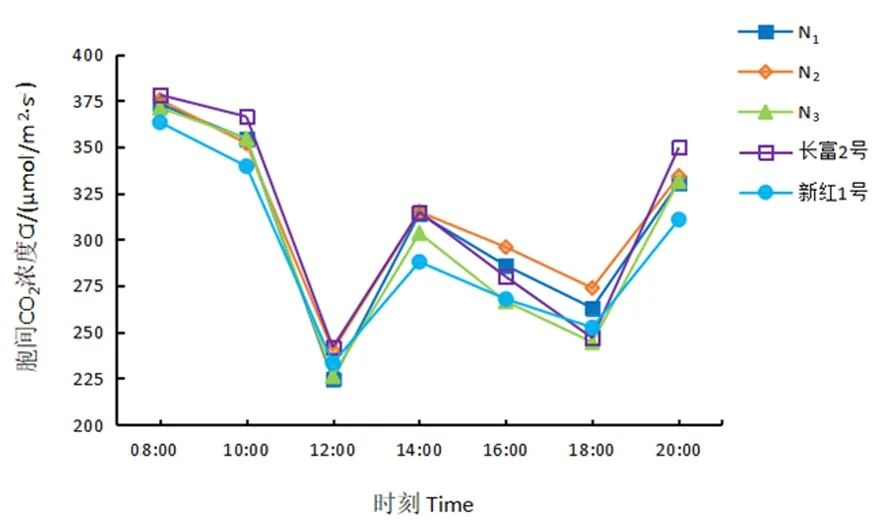
图3 5份苹果CO2日变化Fig.3 Diurnal changes of inter cellular CO2 concentration rate of five apple materials
2.3.4 蒸腾速率日动态变化
研究表明,蒸腾速率的日变化曲线均呈“单峰型”,14:00时长富2号和新红1号蒸腾速率最高,而3个红富士芽变的蒸腾速率高峰出现在16:00,其中N1蒸腾速率最高,达8.64 mmol/(m2·s),长富2号最低,为3.77 mmol/(m2·s)。自08:00起,各供材料随着气温和光合有效辐射的增加,蒸腾速率逐渐上升,芽变材料和品种材料分别在14:00和16:00达到高峰,此后逐渐下降。图4
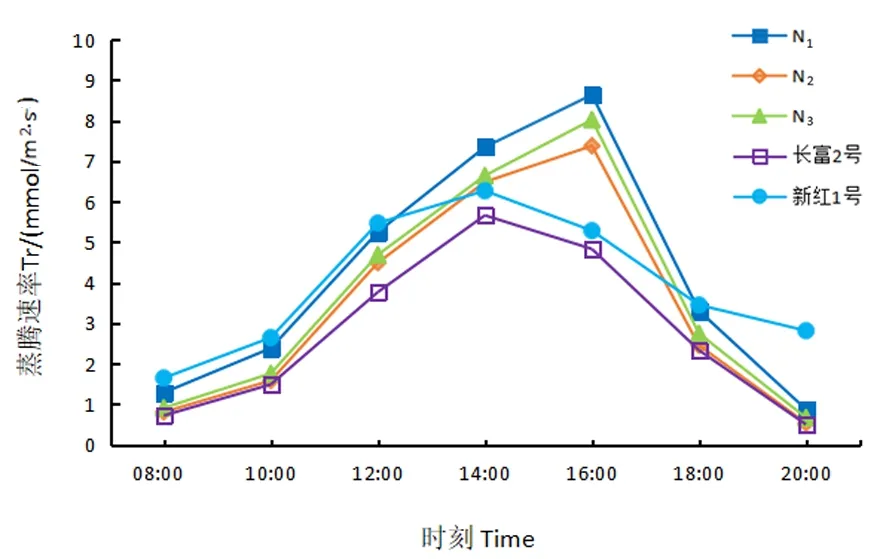
图4 5份苹果Tr日变化Fig.4 Diurnal changes of stalemate transpiration rate of five apple materials
2.3.5 水分利用率日动态变化
研究表明,水分利用率日变化规律基本相近,呈“双峰”型。自08:00时水分利用率逐渐上升,10:00达到高峰,之后水分利用率明显下降,红富士芽变14:00出现低谷,而长富2号和新红1号则在16:00最低,水分利用率缓慢上升,第2次高峰出现在18:00,随后迅速下降。图5
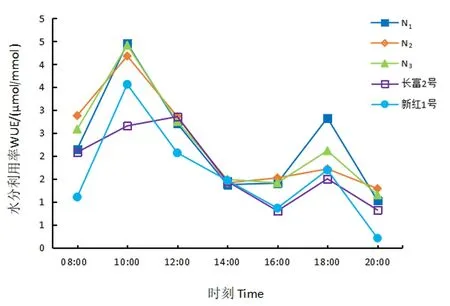
图5 5份苹果WUE日变化Fig.5 Diurnal changes of stalemate water use efficiency rate of five apple materials
2.4 光合特性与叶片特征参数相关性
研究表明,净光合速率与气孔导度、叶绿素含量、水分利用率和蒸腾速率呈极显著正相关(P<0.01),与胞间CO2浓度呈极显著负相关(P<0.01),其中气孔导度相关系数最高,为0.747,其次为叶绿素含量、水分利用率、蒸腾速率。气孔导度与叶绿素含量、蒸腾速率和水分利用率呈极显著正相关(P<0.01);胞间CO2浓度与叶绿素含量呈极显著负相关(P<0.01),与蒸腾速率呈显著负相关(P<0.05),蒸腾速率与叶绿素含量呈极显著正相关(P< 0.01),与水分利用率呈极显著负相关(P<0.01)。叶面积与叶片宽度、单叶重及叶片长度呈极显著正相关(P<0.01);单叶重与叶片长度、叶片宽度呈极显著正相关(P<0.01)。表3

表3 光合特性及与其叶片特征参数间相关性Table 3 Correlation analysis of photosynthetic characteristics and leaf characteristic parameters of five apple materials
3 讨 论
3.1 光合作用是果树生长发育的基础,而叶片是植物接受光的器官,叶片形态、叶绿素含量等均会影响植物光合能力[11-12 ]。不同生长型自身的遗传对叶片特征的影响很大,普通型、短枝型和柱型苹果的叶片特征之间存在很大差异,短枝型的叶片质量较为突出,比叶重较大,短枝型光合性能较高[13-14 ]。研究发现,供试材料叶片长度、叶片宽度、叶片厚度和叶面积之间没有明显差异,而单叶重、叶绿素含量存在明显差异(P<0.05),N1单叶重、叶绿素含量分别为1.09 g、57.42,其有利于同化产物的积累,具有较高的光合效能。光合作用不仅受叶片自身因素影响,还受气孔导度、蒸腾速率、胞间CO2浓度等因素影响。各供试材料光合特征参数存在明显差异,N1净光合速率、气孔导度分别为14.16、0.21 μmol/(m2·s),明显高于其他材料,而胞间CO2浓度为224.53 μmol/(m2·s),显著低于长富2号,其在“午休”期间气孔闭合的条件下对胞间CO2的利用能力较强,表现出较强的光合能力。
3.2 光合日变化既反映环境对光合作用的影响,是果树对生态环境适应能力的主要表现,大多数果树光合日变化呈“单峰型”或“双峰型”曲线[15-16 ]。在试验中,各供试材料净光合速率日变化呈“双峰型”曲线,表现光合“午休”现象。这与多数人研究的结果一致[17-19 ]。也有研究表明,“午休”现象越明显、降幅越大,对强光或空气湿度的适应能力越差[20]。从供试材料光合速率日动态变化来看,N1的2次峰值分别为14.15、12.13 μmol/(m2·s),均较高于其他材料,其适应强光能力较强,具有较高的光合能力,而长富2号的Pn值一直较低,且“午休”现象比较明显,日平均值也较低,表明其光合能力较弱。光合“午休”原因有气孔限制、非气孔限制,如果气孔导度、胞间CO2浓度都随净光合速率降低而降低,是由气孔因素引起;如果气孔导度降低,胞间CO2浓度升高,则为非气孔因素[21-22 ]。在试验中,气孔导度随净光合速率升高而升高,而胞间CO2浓度没有升高,反而降低,芽变光合“午休”主要是非气孔因素造成的,属于非气孔限制。这与曹生奎等[23 ]研究结果相似。蒸腾速率大小可以反映植物调节水分能力以及适应逆境能力的强弱[ 24]。该试验结果表明,N1的蒸腾速率、净光合速率日均值分别为4.16、7.61 μmol/(m2·s),均高于其他供试材料,表明其对分水利用效率较好,耐旱的能力较强,其次为N3,而长富2号耐旱能力最弱。
3.3 有研究表明,光合指标间及其与叶片特征参数之间有密切联系[25-26 ]。研究发现,供试材料的净光合速率与气孔导度、蒸腾速率、水分利用率、叶绿素含量呈极显著正相关,与单叶重呈显著正相关,与胞间CO2浓度呈极显著负相关,与气孔导度相关系数最高,其次为叶绿素含量、水分利用率、蒸腾速率,说明净光合速率受气孔导度影响最大,叶绿素含量次之。这与魏书銮等[27 ]结果研究相似。
4 结 论
4.1 3个红富士芽变类型和2个芽变品种的叶片长度、叶片宽度、叶片厚度和叶面积的之间无明显差异;N1与N2的单叶重之间差异显著,其中N1的叶片最重,为1.09 g;N1与N2、N3、长富2号和新红1号的叶绿素含量之间差异显著,叶绿素含量最高的是N1,为57.42 SPAD。
4.2 3个红富士芽变类型和2个芽变品种的光合特征参数之间有明显差异。N1的净光合速率为14.16 μmol/(m2·s),明显高于N2、N3、新红1号和长富2号;N1叶片的气孔导度最大,为0.21 mol/(m2·s),长富2号的最小,为0.16 mol/(m2·s);长富2号的胞间CO2浓度最高,为241.77 μmol/(m2·s),N1最低,为224.53 μmol/(m2·s)。;新红1号和N1的叶片蒸腾速率较高,分别为5.47 mmol/(m2·s)、5.25 mmol/(m2·s),长富2号最低,为3.77 mmol/(m2·s),;N2的水分利用率较高,为2.85 μmol/mmol,其次为N1和N3,其值均为2.75 μmol/mmol,长富2号最低,为2.07 μmol/mmol。
4.3 3个红富士芽变类型和2个芽变品种叶片的净光合速率、水分利用率日变化均呈双峰型曲线;N1、N2和N3的气孔导度日变化曲线呈双峰型,长富2号和新红1号呈单峰型;3个红富士芽变类型和2个芽变品种的胞间CO2浓度日变化呈W型;蒸腾速率的日变化曲线均呈单峰型。
4.4 3个红富士芽变类型和2个芽变品种的净光合速率与气孔导度、叶绿素含量、水分利用率、蒸腾速率呈极显著正相关(P<0.01),与胞间CO2浓度呈极显著负相关(P<0.01),其中气孔导度相关系数最高,为0.747,其次为叶绿素含量、水分利用率、蒸腾速率,其值分别为0.653、0.619和0.511。
芽变N1的净光合速率、气孔导度、单叶重和叶绿素含量明显高于N2、N3、新红1号和长富2号,表现较强的光合能力。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
湖北农业科学(2021年13期)2021-08-04 03:56:18
新疆农业科技(2021年2期)2021-07-03 06:39:34
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
中学生物学(2020年12期)2020-03-30 07:27:46
中国果业信息(2018年11期)2018-01-17 10:33:55
幸福·悦读(2016年8期)2016-10-21 12:18:20
生物技术通报(2014年10期)2014-03-21 06:56:38
