
非洲猪瘟多基因家族MGF-505-3R 基因重组原核表达质粒的构建及表达
2021-06-18 01:45武悦许会会徐一鸣王楠王泽宇杨莲花鲁中爽蔡维北吕忠蕾李霜
中国生物制品学杂志 2021年6期
武悦 ,许会会 ,徐一鸣 ,王楠 ,王泽宇 ,杨莲花 ,鲁中爽,蔡维北,吕忠蕾,李霜
1.吉林元和元生物工程有限公司,吉林长春130102;2.吉林省疫病预防控制中心,吉林长春130102
2018 年8 月1 日,辽宁沈阳一养猪户饲养的猪陆续死亡,疑似非洲猪瘟病毒(African swine fever virus,ASFV)感染。经检测,确诊为ASFV 核酸阳性,序列分析发现,其基因序列与俄罗斯毒株100%匹配。这是我国发现的首起非洲猪瘟。
ASFV 是一种大型双链DNA 病毒,是Asfarviridae 家族的唯一成员。ASFV 可感染家猪、野猪,它们可能会成为一个载体,主要靶点是猪单核巨噬细胞。ASFV 能引起高烧,出血性损伤,家猪厌食,甚至死亡。目前尚无没有可用的ASFV 疫苗,也无有效的措施控制病毒传播,给全球养猪业造成了极大的负担[1-3]。
MGF 严格来说不是指一个单独的基因,而是一个多基因家族,MGF 505 是其中更小的家族基因,MGF-505-3R 中的3R 为非洲猪瘟基因缺失疫苗缺失毒株的基因结构向右侧方向的第3 段基因,只是位次关系,并无特殊含义。ASFV 编码的多个多基因家族蛋白中MGF 505 决定了ASFV 的细胞嗜性,与病毒在巨噬细胞内的复制密切相关,是ASFV 毒力的决定因素[2,4-6]。MGF 505 还参与抑制宿主干扰素的产生和调控促炎性细胞因子表达,并通过延长感染细胞的存活时间来提高病毒在宿主细胞内的增殖效率和数量[7-8]。此前,军事兽医研究所以我国首起非洲猪瘟病毒中分离到的流行毒株(SY-18)为亲本构建了MGF 基因缺失的ASFV 基因缺失疫苗候选株,结果显示,接种猪能够100%抵抗亲本毒株SY-18 的攻击,因此,以MGF 为靶标的基因缺失毒株是较有前景的ASFV 疫苗候选毒株[9]。
本研究旨在构建ASFV SY-18 株多基因家族MGF-505-3R 基因的重组原核表达质粒,并表达重组蛋白,以期提供有效的抗原蛋白用于ASFV 的临床诊断及防控。
1 材料与方法
1.1 菌株、载体及基因组 大肠埃希菌DH5α 和BL21(DE3)感受态细胞购自宝日医生物技术有限公司;原核表达载体pET-32a(+)购自诺唯赞生物公司;ASFV 基因组由军事医学科学院军事兽医研究所扈荣良研究员惠赠。
1.2 主要试剂 基因组提取试剂盒、质粒DNA 小量纯化试剂盒和DNA 凝胶回收试剂盒均购自康宁生命科学有限公司;Premix Prime-STAR HS 高保真DNA聚合酶、限制性内切酶XhoⅠ和EcoRⅠ及DNA marker均购自宝日医生物技术有限公司;IPTG 购自诺唯赞生物公司;单片段无缝克隆试剂盒购自莫纳(武汉)生物科技有限公司。
1.3 引物设计及合成 根据NCBI 中ASFV SY-18 株基因的核苷酸序列(MN393476.1)设计1 对特异性引物,上游引物 MGF-505-3R-F 序列:5′-ggtggtggtgctcgagatgtcctcttctcttcaggaac-3(′下划线部分为EcoRⅠ酶切位点),下游引物 MGF-505-3R-R 序列:5′-tatcggatccgaattcctagctttcccaggtttcgaag-3(′下划线部分为XhoⅠ酶切位点),并加入保护性碱基,扩增片段大小为843 bp。引物由生工生物工程(上海)股份有限公司长春分部合成。
1.4 MGF-505-3R 基因的扩增 以ASFV 基因组为模板,PCR 扩增MGF-505-3R 基因。反应体系:Premix Prime-STAR HS 10 μL,ddH2O 7 μL,上、下游引物各 1 μL,模板 1 μL,共 20 μL。反应条件:95 ℃预变性3 min;95 ℃变性15 s,退火温度设置为梯度(45 ~ 55 ℃之间)15 s,72 ℃延伸 15 s,共 35 个循环;72 ℃再延伸7 min。经凝胶电泳验证后,发现在51 ℃时条带最亮,因此用上述20 μL 体系做了5 管PCR 扩增,共100 μL。PCR 产物经1%琼脂糖凝胶电泳鉴定后,用胶回收试剂盒纯化MGF-505-3R 基因片段。
1.5 重组原核表达质粒的构建 用EcoRⅠ和XhoⅠ双酶切pET-32a(+)载体,胶回收试剂盒纯化回收。用无缝克隆试剂盒将纯化回收的目的基因MGF-505-3R 及双酶切载体pET-32a(+)连接,连接产物转化至大肠埃希菌DH5α 感受态细胞中,涂布于含1 mg / mL 氨苄西林的固体 LB 平板上,37 ℃静置过夜。挑取7 个单菌落,进行菌液PCR(反应体系:高保真 DNA 聚合酶 Premix Prime-STAR HS 10 μL,上下游引物各 1 μL,模板 1 μL,ddH2O 补足体积至 20 μL。反应条件:95 ℃预变性 3 min;95 ℃变性 15 s,51 ℃退火15 s,72 ℃延伸15 s,共35 个循环;72 ℃再延伸7 min)及质粒双酶切鉴定,并送生工生物工程(上海)股份有限公司测序。
1.6 生物信息学分析 通过ExPASy 在线预测分析蛋白质的理化性质,TMPred 法和HMM 法预测分析蛋白质的跨膜区,PSIPRERD 法在线预测分析蛋白质的二级结构,通过同源建模SWISS-MODEL 在线预测蛋白质的三级结构。
1.7 重组MGF-505-3R 蛋白的诱导表达及纯化 将鉴定正确的重组原核表达质粒pET-32A(+)-MGF-505-3R 转化入大肠埃希菌BL21(DE3)中,挑取单个菌落,接种至含1 mg/mL 卡那霉素的LB 液体培养基中,37 ℃振荡培养过夜,当菌液A600为0.8 ~1时,加入 1 mol / L IPTG,使其终浓度为 1 mmol / L,37 ℃继续培养6 h;分别各取1 mL 诱导前后菌液,6 000 × g离心10 min,收集菌体沉淀,用30 μL PBS 重悬,取样进行10%SDS-PAGE 分析,经考马斯亮蓝染色3 h,脱色2 次后观察结果。
纯化过程利用凝胶扫描法计算融合蛋白的表达量,菌体超声破碎后获得的上清液经Ni-NTA 亲和层析和CMSepharose FF 阳离子交换层析纯化,10%SDS-PAGE 分析蛋白纯度,BCA 法测定蛋白产量。
2 结 果
2.1 MGF-505-3R 基因扩增产物的鉴定 PCR 产物经1%琼脂糖凝胶电泳分析,可见843 bp 的特异性条带,大小与预期一致,见图1。
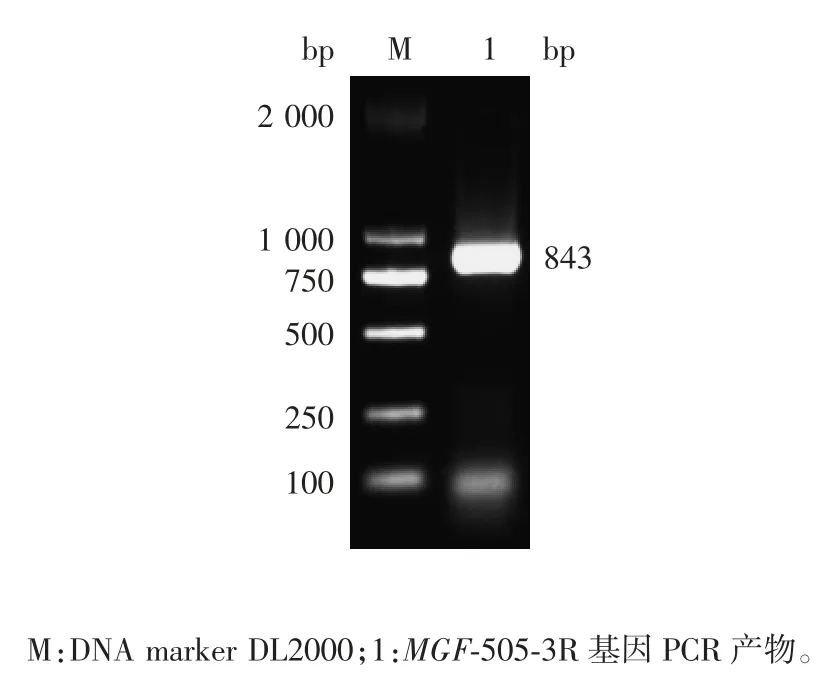
图1 ASFV MGF-505-3R 基因PCR 产物电泳图Fig.1 Electrophoretic profile of PCR product of ASFV MGF-505-3R gene
2.2 重组原核表达质粒的鉴定 重组原核表达质粒pET-32A(+)-MGF-505-3R 经EcoRⅠ和XhoⅠ双酶切及菌液PCR 鉴定,均可见843 bp 的目的基因片段,见图2 和图3。测序结果表明,重组质粒构建正确。
2.3 生物信息学分析
2.3.1 蛋白质的理化性质 ExPASy 在线预测分析显示,MGF-505-3R 基因编码氨基酸数量为843 个,经分析,不稳定指数为45.78,将MGF-505-3R 蛋白分类为不稳定的蛋白。亲水性平均值(GRAVY)为0.801,为疏水蛋白。
2.3.2 蛋白质跨膜区预测 TMPred 法和HMM 法分析蛋白质的跨膜区,预测图形结果见图4 和图5,分析的3 个蛋白质的TM-螺旋长度均在17 ~33 之间。TMH 数为24,为跨膜蛋白。
2.3.3 蛋白质二级结构预测 PSIPRERD 法在线预测分析蛋白二级结构显示,该蛋白由无规卷曲(random coil)、α 螺旋(alpha helix)和 β 折叠(β-sheet)组成为主要结构,见图6。
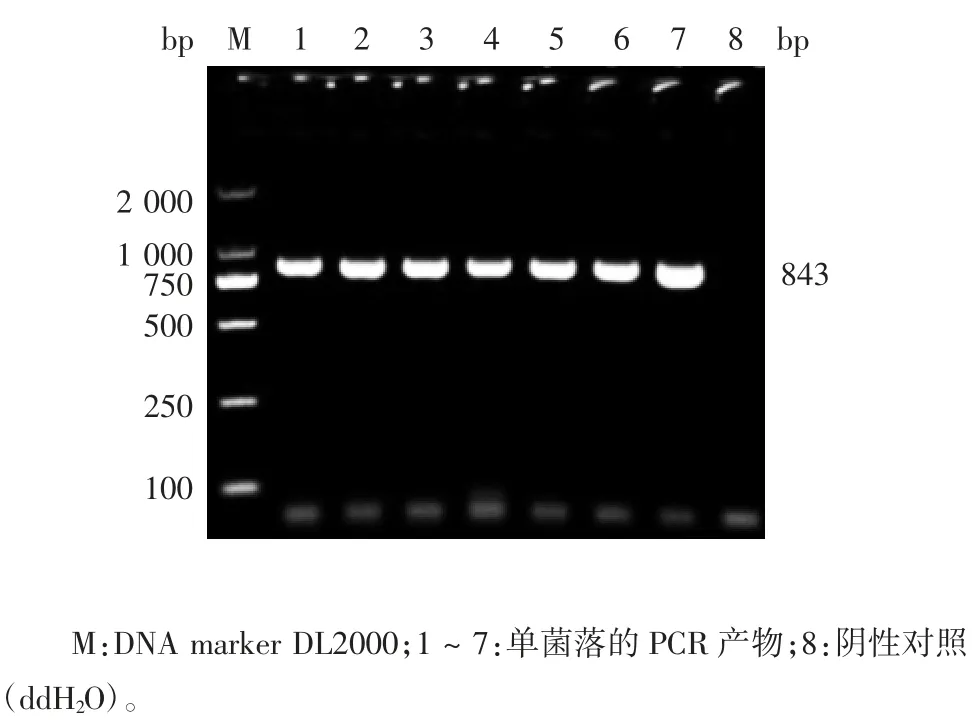
图2 重组原核表达质粒的菌液PCR 鉴定Fig.2 Identification of recombinant prokaryotic expression vector by PCR
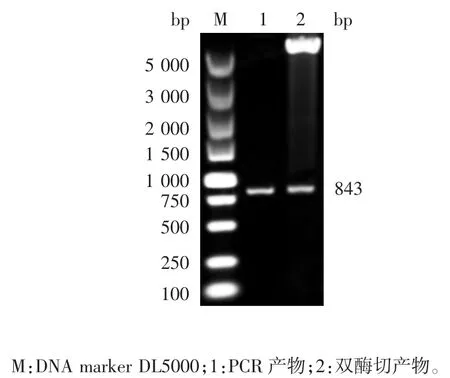
图3 重组原核表达质粒的双酶切鉴定(EcoRⅠ/ XhoⅠ)Fig.3 Restriction analysis of recombinant plasmid pET-32a(+)(EcoRⅠ/ XhoⅠ)
图4 TMPred 法蛋白质跨膜域分析Fig.4 Analysis of transmembrane domain of protein by TMPred
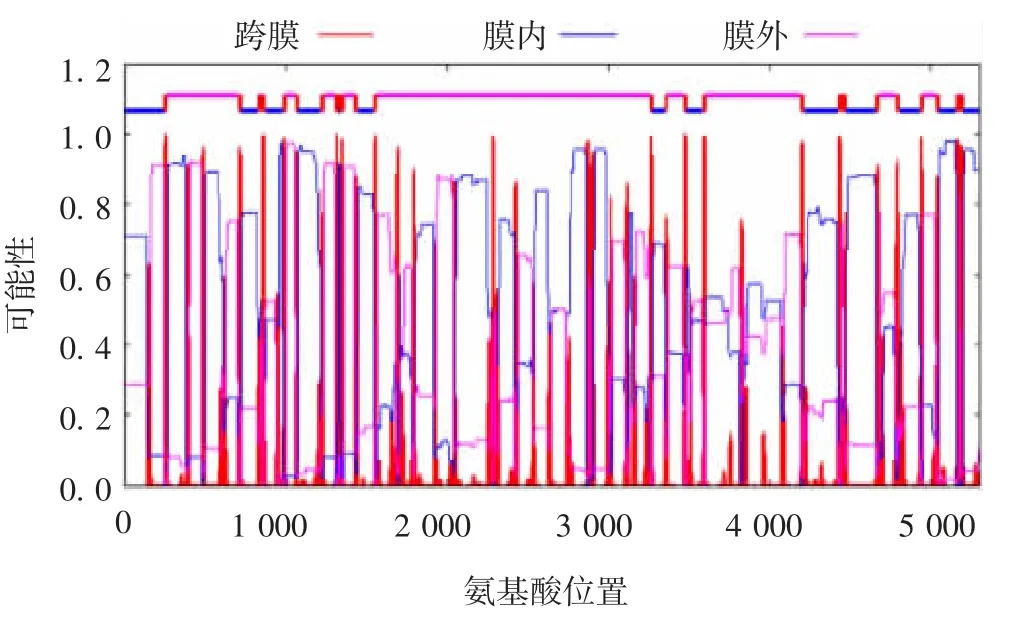
图5 HMM 法蛋白质跨膜域分析Fig.5 Analysis of transmembrane domain of protein by HMM
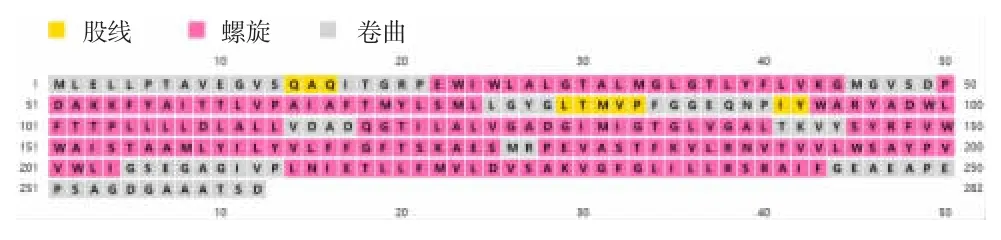
图6 蛋白质二级结构分析Fig.6 Secondary structure of protein
2.3.4 蛋白质三级结构预测 蛋白质同源建模结构见图7。无规则卷曲、α 螺旋 和β 折叠组成为主要成分,预测结果与二级结构预测相符。
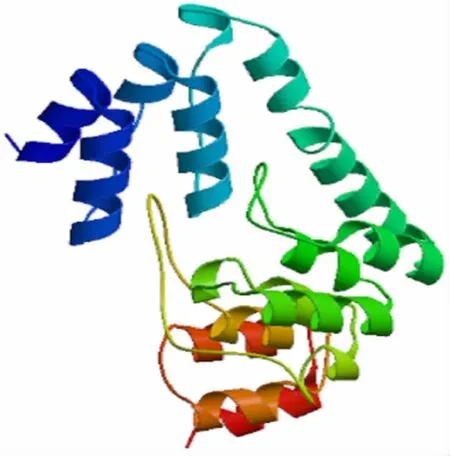
图7 编码蛋白质的三级结构Fig.7 Tertiary structure of protein
2.4 表达及纯化产物的鉴定 10% SDS-PAGE 分析显示,表达的HIS 标签重组蛋白相对分子质量约为33 000,大小与预期相符,见图8。经凝胶扫描法计算融合蛋白表达量为37.8 mg / L。BCA 法测定重组蛋白产量可达19.8 mg / L。软件分析纯化后蛋白纯度为97%。
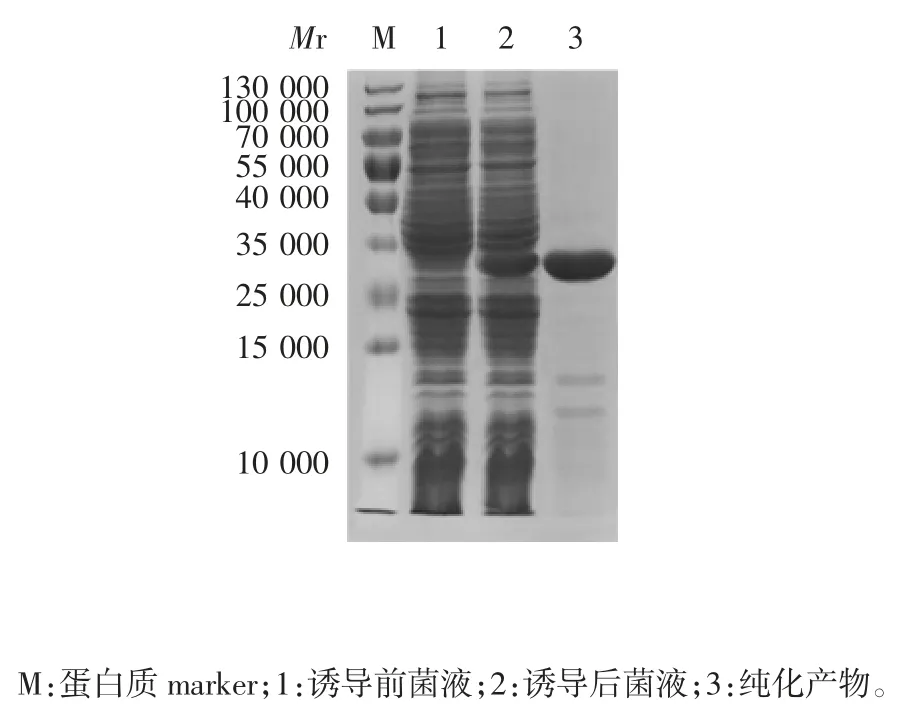
图8 表达及纯化产物的SDS-PAGE 分析Fig.8 SDS-PAGE profile of expressed and purified protein
3 讨 论
非洲猪瘟暴发速度极快,致死率接近百分之百。目前尚无有效的预防疫苗及治疗措施,给养殖业带来了巨大损失。因此,在无法建立该病血清学检测技术的情况下,利用重组ASFV 抗原建立快速、安全的诊断和检疫方法十分必要。
ASFV 的基因家族十分庞大,且MGF 多基因家族中MGF-505 蛋白是很重要的结构蛋白。王晓丽等[10]研究表明,MGF-505 家族成员 pA528R 能通过抑制Poly(I:C)诱导的IRF3 和NF-ΚB 活性来抑制IFN 的表达。CORREIA 等[11]研究表明,MGF-530 / 5 05 的多个基因缺失后,可导致Ⅰ型IFN 在感染巨噬细胞中的诱导水平增加,且缺失后的毒株对Ⅰ型IFN 敏感,表明这些基因具有抑制IFN 应答和抗病毒状态的功能。研究表明,大部分基因缺失疫苗针对亲本毒株的攻击具有同源保护作用,部分也具有一定的异源保护作用[10,12-13]。因此,ASFV 基因缺失疫苗免疫保护效果较理想,是短期内最有望研制成功的疫苗[10,14-16]。
敲除MGF-505-3R 所获得的基因缺失毒株能够诱导机体产生抵抗亲本强毒株的保护作用,因此,以ASFV MGF-505-3R 基因为靶标开发基因缺失减毒活疫苗具有较大的可行性及实用价值[2,17-19]。本研究根据SY-18 株病毒基因组成功克隆了在ASFV 中稳定存在的MGF-505-3R 基因,构建了重组原核表达质粒并成功表达了重组蛋白,对研制非洲猪瘟抗体检测试剂盒具有重要意义。但表达的重组蛋白表达量不高,后续将对试验方法进行改进,以提高重组蛋白的表达量及纯度,为我国非洲猪瘟免疫血清学诊断试剂的制备及预防控制奠定基础。
猜你喜欢
科学大观园(2022年2期)2022-01-23
幸福(2019年12期)2019-05-16
猪业科学(2018年8期)2018-09-28
兽医导刊(2016年6期)2016-05-17
安徽医科大学学报(2015年9期)2015-12-16
动物医学进展(2015年10期)2015-12-07
动物医学进展(2015年10期)2015-12-07
动物医学进展(2015年10期)2015-12-07
现代检验医学杂志(2015年5期)2015-02-06
特产研究(2014年4期)2014-04-10
