
炒酸枣仁对状态性焦虑大鼠基底外侧杏仁核神经元信息编码的影响研究
2021-05-13 11:19华伊张正一黄莉莉王艳艳叶晓楠李廷利
中医药学报 2021年4期
华伊,张正一,黄莉莉,王艳艳,叶晓楠,李廷利
(黑龙江中医药大学,黑龙江 哈尔滨 150040)
酸枣仁为鼠李科植物酸枣(ZiziphusjujubaMill.var.spinosae(Bunge)Hu ex H.F.Chou)的干燥成熟种子,主治虚烦不眠、惊悸多梦、体虚多汗等症[1]。现代研究发现酸枣仁具有一定的抗状态性焦虑作用[2-3],酸枣仁在临床的应用普遍使用炒制品。状态性焦虑指机体面对某一特定情境而产生的暂时的紧张不安的情绪状态,旷场实验是一种被广泛应用的状态性焦虑动物模型建立及行为学评价方法[4-5]。在脑科学领域,杏仁核(Amygdala)被普遍认为是情绪调节尤其是焦虑、恐惧等负性情绪调节过程中的最关键脑区[6-7],杏仁核的基底外侧核团(Basolateral amygdala,BLA)是进行焦虑处理时接收外界刺激传入的主要部位[8]。目前为止,炒酸枣仁的抗状态性焦虑作用在神经元在体电活动方面的机制尚未清晰。基于以上,本文利用旷场实验方法和在体多通道神经同步记录技术,拟考察炒酸枣仁水煎液对状态性焦虑大鼠的旷场行为学及BLA区神经元信息编码的影响,探讨炒酸枣仁对状态性焦虑的干预作用及神经电生理方面的作用机理。
1 实验材料
1.1 实验动物
SD大鼠,雄性,体质量(220±20)g,无特定病原体(SPF)级,购于黑龙江中医药大学GLP实验动物中心,许可证号:SCXK(黑)2013-004。
动物饲养与监测环境维持为通风、避光、隔音、电磁屏蔽状态,并通过光控定时开关程序进行12 h/12 h明暗交替光照处理,室内温度:20~24 ℃,相对湿度:50%~60%,噪声≤40 dB。
1.2 实验药物
酸枣仁,原产地山西,依照文献[9]方法,制备炒酸枣仁和炒酸枣仁水煎液。
1.3 实验试剂
生理盐水(170104D5,哈尔滨三联药业有限公司),戊巴比妥钠(WS20140104,国药集团化学试剂有限公司),义齿基托聚合物(170308,上海Pigeon Dental有限公司),义齿基托树脂(20170408,上海New Century Dental Materials有限公司),多聚甲醛(20161130,天津基准化学试剂有限公司),无水乙醇(20160408,天津市天力化学试剂有限公司)等。
1.4 实验仪器
半微量分析天平(MS105/A,梅特勒-托利多仪器上海有限公司),旷场实验箱(100 cm×100 cm×40 cm,木制),标准脑立体定位仪(51600,美国Stoelting公司),在体多通道神经记录分析系统(OPX-32D,美国Plexon公司),低温冷冻组织切片机(CM1950,德国Leica公司),强制对流标准型烘箱(Venticell 111,德国 MMM 集团公司)等。
2 实验方法
2.1 在体多通道记录电极的埋植
选取大鼠20只,经戊巴比妥钠(40 mg/kg)腹腔注射麻醉后,借助脑立体定位仪,图谱[10],确定BLA区位置(坐标:前囟后2.5~2.8 mm,中缝线右4.5~4.8 mm,颅骨下7.5 mm)。将16(4×4阵列)通道在体记录微电极(铂铱合金,电极丝外周涂有绝缘层,直径35 μm)埋置于大鼠的BLA区域,用义齿基托树脂将整个电极部分固定在大鼠颅顶。手术后每只大鼠单独饲养恢复5 d。
2.2 动物的分组及给药
将已植入电极并经过术后恢复的大鼠随机分为模型对照组与炒酸枣仁组,每组各10只。经灌胃途径给予炒酸枣仁组大鼠炒酸枣仁水煎液(17.5 g/kg),模型对照组灌服相同体积的蒸馏水。
2.3 模型的制备及行为学测试
依照文献[11]方法,利用旷场实验制备大鼠状态性焦虑模型。给药0.5 h后,将各组大鼠放入旷场箱内底面中心,任其自由探索,记录并分析10 min内大鼠在旷场环境中的行为学表现。测试指标为跨格次数、站立次数、进入中央格次数、中央格运动时间(s)和总运动时间(s),并以中央格运动时间(s)除以总运动时间(s)计算中央格/总运动时间(%)。
2.4 神经元放电信号的采集
给药0.5 h后,利用在体多通道神经记录分析系统,先记录各组动物在日常饲养环境中10 min内的神经元基础放电信号,再在进行旷场实验的同时记录旷场环境中10 min内的神经元旷场放电信号。利用Offline Sorter软件对采集到的神经元放电信号进行处理,过滤低频放电和环境干扰放电,设置信噪比大于3的信号为所需的动作电位(Spikes)信号,运用主成分分析(Principal Component Analysis,PCA)技术对分离出的动作电位进行聚类处理,测试指标为神经元动作电位的放电频率(spikes/s)。
2.5 组织学考察
电信号采集全部结束后,各组动物颅顶在体记录电极经电刺激器进行通电刺激(1 mA,10 s,5次左右),摧毁电极末端脑组织。灌流取脑,利用脑组织冰冻切片的HE染色技术判定记录电极是否准确植入大鼠的BLA区域,偏离位置的数据不予采用。
2.6 统计学分析
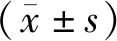
3 结果
3.1 炒酸枣仁对状态性焦虑大鼠行为学的影响
实验结果表明,与模型对照组相比,炒酸枣仁组大鼠在旷场中的跨格次数、站立次数及总运动时间无明显变化(P>0.05),中央格运动时间、进入中央格次数及中央格/总运动时间(%)明显增加(P<0.01)。见表1。
表1 各组大鼠旷场行为学比较
3.2 炒酸枣仁对状态性焦虑大鼠BLA区神经元信息编码的影响
3.2.1 组织学考察
大鼠脑组织冰冻切片的HE染色结果显示,在体多通道记录电极准确植入大鼠BLA区。切片见图1。
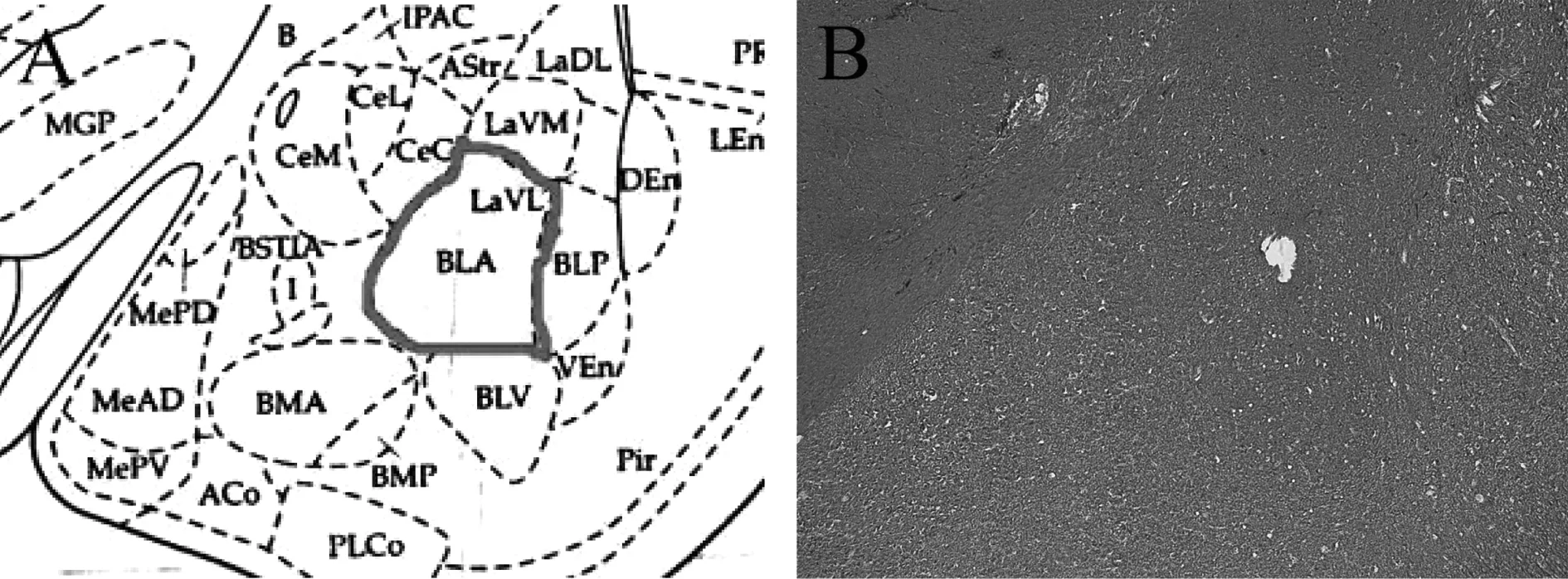
注:图A为大鼠脑立体定位图谱所示BLA区位置;图B为大鼠脑组织切片HE染色图,空腔指示为记录电极末端所处位置。图1 记录电极定位HE染色结果(50×)
3.2.2 大鼠BLA区神经元动作电位分类
本实验在大鼠BLA区记录到的神经元动作电位波形特征为:波长较长,波峰后曲线下降缓慢。按照放电模式的不同,把在BLA区记录到的神经元分为Ⅰ、Ⅱ两类(以下称BLA区Ⅰ类神经元为NeuronⅠ,Ⅱ类神经元为NeuronⅡ)。NeuronⅠ的时长较长、振幅稍低,自相关图中出现典型的尖峰、峰值后呈现非常陡的下降趋势。NeuronⅡ的时长较短、振幅较高,自相关图中峰值后下降较缓慢。两类神经元动作电位放电模式的特征图见图2。
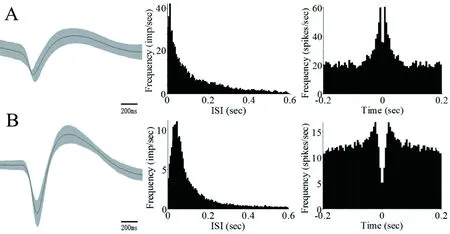
注:图A为NeuronⅠ动作电位的波形图、放电间隔直方图及自相关图,图B为NeuronⅡ动作电位的波形图、放电间隔直方图及自相关图。图2 BLA区两类神经元动作电位特征图
3.2.3 炒酸枣仁对状态性焦虑大鼠BLA区神经元动作电位放电频率的影响
实验结果表明,与自身的基础放电频率相比,模型对照组和炒酸枣仁组大鼠在旷场中的NeuronⅠ、NeuronⅡ动作电位放电频率均有所增加,放电频率直方图见图3。将每只大鼠的旷场放电频率减去其基础放电频率,得出的值定义为神经元动作电位放电频率增加值,以此值进行两组大鼠在旷场中神经元动作电位放电频率变化的对比。实验结果表明,与模型对照组相比,炒酸枣仁组大鼠BLA区NeuronⅠ的动作电位放电频率增加值明显减少(P<0.05),NeuronⅡ的动作电位放电频率增加值无明显变化(P>0.05),见表2。
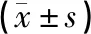
表2 各组大鼠BLA区神经元动作电位放电频率增加值比较
3.3 大鼠旷场行为学与BLA区神经元信息编码的相关性分析
实验结果表明,大鼠在旷场中的中央格/总运动时间(%)与BLA区NeuronⅠ的动作电位放电频率增加值之间存在明显的负相关(r=-0.614,P<0.01),与NeuronⅡ的动作电位放电频率增加值之间存在明显的负相关(r=-0.504,P<0.05)。见图4。
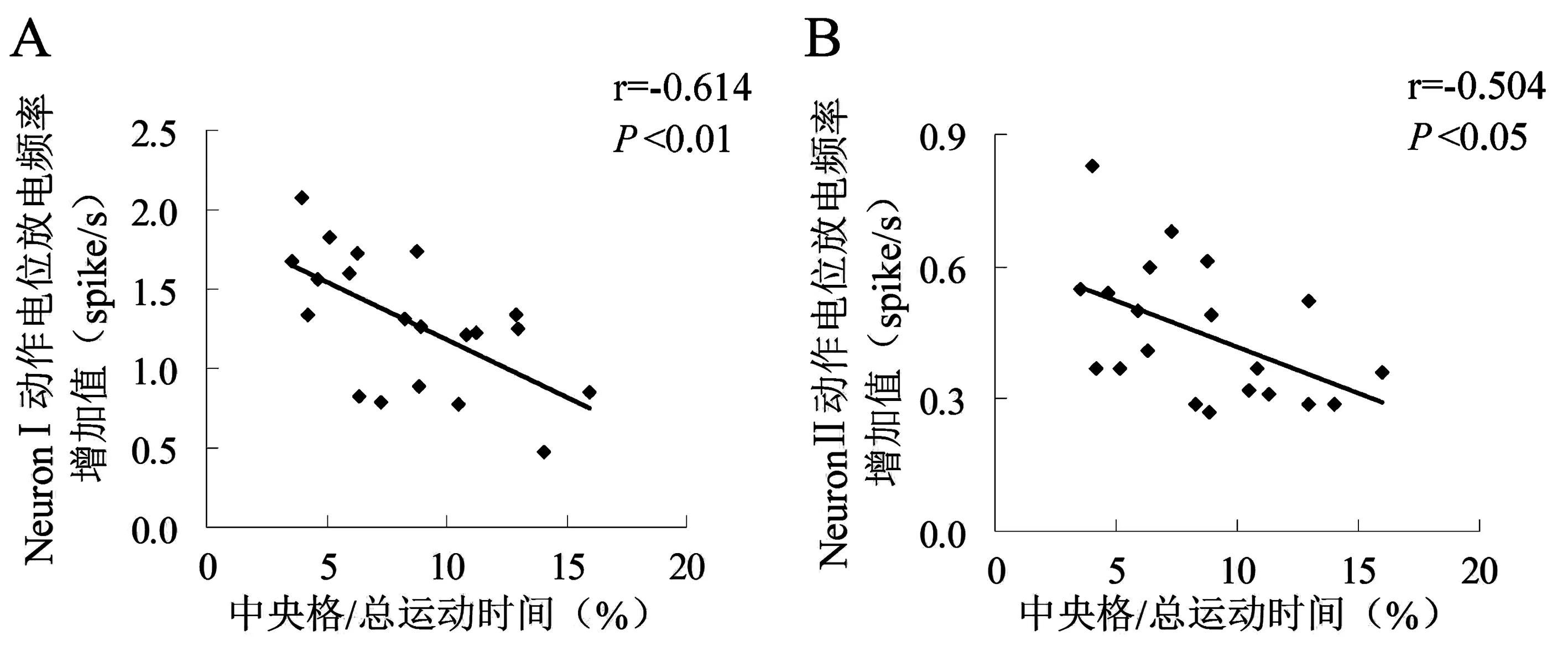
图4 旷场中央格/总运动时间(%)与BLA区神经元动作电位放电频率增加值的相关性分析
4 讨论
旷场实验利用了啮齿动物害怕空旷环境的天性,但又对陌生场地产生好奇而试图探索的矛盾心理[12],被广泛应用于状态性焦虑动物模型的复制与评价[13-14]。本文利用旷场实验建立大鼠状态性焦虑模型,运用行为学实验方法和在体多通道神经同步记录技术,给予炒酸枣仁水煎液的干预,记录并分析了炒酸枣仁对状态性焦虑大鼠的旷场行为学及BLA区神经元信息编码的影响。
对啮齿动物而言,旷场的中央格区域具有潜在威胁,周边格相对安全,大鼠的焦虑程度高时会更偏向于在周边格活动[15-16]。旷场中的跨格次数、站立次数和总运动时间等行为学指标可用于评价大鼠的活动性和探索行为,进入中央格次数、中央格运动时间及中央格/总运动时间则指示了大鼠的焦虑水平。在行为学实验中,给予炒酸枣仁水煎液干预后,大鼠的跨格次数、站立次数及总运动时间有所增多,运动性与探索行为增加,这与报道[17]相符。同时,炒酸枣仁能明显增加大鼠的中央格运动时间、进入中央格次数及中央格/总运动时间(%),大鼠的焦虑样行为减少,与前人研究[18]结果一致,提示炒酸枣仁能减轻旷场对大鼠造成的焦虑情绪。
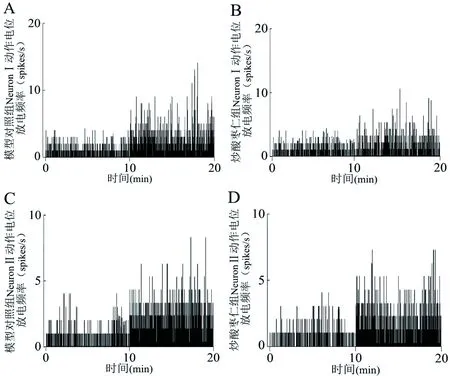
注:0~10 min为大鼠在日常饲养环境中的基础放电,10~20 min为在旷场环境中的放电。图3 旷场对大鼠BLA区神经元动作电位放电频率的影响
大脑对信息的处理是通过神经元群的协同工作实现的,动作电位则是神经元进行信息传递的基本语言[19]。神经元动作电位的发放具有“全或无”的特点,外界刺激信息发生改变时,动作电位的放电频率也会发生相应的改变。神经元信息编码的研究本质上就是对神经元动作电位的放电频率进行的研究,这对于探索大脑进行各种智能行为的神经机制、揭秘大脑对外部事件的编码机制具有重要意义[20]。杏仁核神经元网络的同步活动在大脑情绪功能中发挥重要的作用[4-5]。大脑进行焦虑调节的过程中,感觉信息从丘脑和感觉皮质传递到基底外侧杏仁核(BLA),然后投射到中央杏仁核,再传递到下丘脑和脑干结构,产生自主神经等反应[21]。焦虑发生的同时伴有杏仁核功能的改变[21-22],焦虑反应的增加与BLA区的异常活动有关[23]。
我们运用在体多通道神经同步记录技术,通过铂铱合金阵列记录微电极,监测大鼠的在体BLA区神经元同步电信号。本文记录到的BLA区神经元动作电位的波形特征为波长较长,波峰后曲线下降缓慢,与文献[24]报道一致。按照发放特征的不同,将记录到的BLA区神经元分为NeuronⅠ与NeuronⅡ。本研究发现,与自身的基础放电相比,各组大鼠在旷场中NeuronⅠ、NeuronⅡ的动作电位放电频率都有所增加。这一现象反映了大鼠面对旷场环境时,其BLA区神经元信息编码发生了改变,与前人报道[25]一致。我们将各只实验动物的旷场放电频率减去其基础放电频率,得出的值定义为神经元动作电位放电频率增加值,以此值进行两组大鼠神经元动作电位放电频率变化的对比。结果显示,炒酸枣仁明显降低了大鼠BLA区NeuronⅠ的动作电位放电频率增加值,提示炒酸枣仁通过降低BLA区NeuronⅠ的动作电位放电频率,抑制旷场对大鼠造成的焦虑状态。
由于旷场的中央格区域对动物而言存在潜在危险,动物的焦虑程度越高,越不喜爱在中央格运动,实验动物在旷场中的中央格/总运动时间(%)这一指标与动物的焦虑程度呈负相关[14]。相关性分析结果显示,大鼠在旷场中的中央格/总运动时间(%)与BLA区NeuronⅠ、NeuronⅡ的动作电位放电频率增加值间均存在明显的负相关,证明BLA区神经元的活动情况确与动物的焦虑状态密切相关,且本实验采集到的两类BLA区神经元的活动性均随焦虑程度的增加而增加。由于旷场兴奋了大鼠的Ⅰ、Ⅱ两类BLA区神经元,进一步证实了旷场中的大鼠处于焦虑状态。同时我们发现,BLA区NeuronⅠ和NeuronⅡ均为焦虑相关神经元,但是炒酸枣仁只降低了NeuronⅠ的动作电位放电频率,而NeuronⅡ的放电情况无明显变化。推测炒酸枣仁并不是对BLA区所有类型的神经元均有干预作用,炒酸枣仁对焦虑的拮抗作用也并不是只通过某一个脑区,而是多个脑区共同作用的结果,具体原因还需进一步研究。
综上所述,炒酸枣仁具有拮抗状态性焦虑的作用,其神经电生理作用机制与降低BLA区神经元动作电位放电频率、改变BLA区神经元信息编码有关。本研究结果在神经电生理层面阐述炒酸枣仁的抗状态性焦虑作用,进一步明确焦虑样行为与BLA区神经元在体活动的关系,为焦虑相关的新药研发和临床应用奠定理论基础。
猜你喜欢
——从一道浙江选考生物学试题谈起
中学生物学(2022年9期)2022-11-11
食品安全导刊(2021年21期)2021-08-30
电子产品世界(2021年8期)2021-01-16
心肺血管病杂志(2020年5期)2021-01-14
中国计算机报(2019年49期)2019-02-07
今日农业(2019年5期)2019-01-03
中国新闻周刊(2017年36期)2017-10-21
妇女之友(2017年6期)2017-08-01
绿色科技(2017年10期)2017-07-05
老年世界(2017年2期)2017-03-16
