
锌指蛋白家族在乳腺癌发生发展中的作用及机制
2021-04-29 11:12:22综述何咏茵孙朝晖李林海审校
福建医科大学学报 2021年1期
雷 婷(综述),肖 斌,何咏茵,孙朝晖,李林海(审校)
转录因子(transcription factor,TF)在真核生物中广泛存在,是一类能与特异DNA序列结合的核内蛋白质因子,可与启动子顺式元件作用或与其他TF的功能区域相互作用来调控基因的转录表达。根据不同的DNA结合基序,TF主要分为经典锌指、同源域和基本的螺旋-环-螺旋。近期研究表明,大量锌指蛋白在乳腺癌组织和细胞中异常表达,与乳腺癌发生、侵袭和转移等进程相关[1-3]。因此,揭示锌指蛋白在乳腺癌发生、发展中的作用机制有助于开发乳腺癌新的诊断或治疗靶点。本研究旨在探讨锌指蛋白的结构、分类及其在乳腺癌发生发展中的作用机制。
1 锌指蛋白结构及分类
锌指蛋白是指含有稳定“指状结构”的一类蛋白质,该结构通过结合Zn2+自我折叠形成。锌离子的存在是锌指蛋白发挥调控作用的关键:当锌离子缺乏时,锌指蛋白与DNA/RNA序列结合的特异性会被显著抑制,同时,蛋白本身的结构稳定性也会被破坏,影响基因表达。
根据与锌离子结合的保守性氨基酸残基Cys和His的不同组合,Krishna等[4]将锌指蛋白分为8个折叠群,包括类C2H2型锌指(C2H2-like)、塞结状锌指(gag knuckle)、高音谱号锌指(treble clef)、带状锌指(zinc ribbon)、Zn2Cys6锌指、类TAZ2型锌指(TAZ2-domain like)、锌离子结合短环锌指(short zinc binding loops)及金属硫蛋白锌指(metallothionein);也可根据锌指结构序列和功能的不同将其分为C2H2型、RING型(C3HC4)、LIM型(C2HC5)、C4型等9大类别[5]。
C2H2型是研究最为广泛的锌指蛋白,以单体形式与核酸发生相互作用,可根据其N端结构域进一步分为KRAB型(krüppel-associated boxes)、SCAN型(SRE-ZBP,CTfin51,AW-1 and number 18 cDNA)、BTB(broad-complex,tramtrack,and bric-a-brac)/POZ(poxvirus and zinc finger)型等[6],这些功能结构域通过调控TF之间或与其他细胞成分间的选择性结合来控制亚细胞定位、DNA结合和基因表达。
2 与乳腺癌相关的锌指蛋白
2.1锌指蛋白217(zinc finger protein 217,ZNF217)
ZNF217位于染色体20q13.2区域,其mRNA表达水平可作为ER(+)乳腺癌患者内分泌治疗反应的预测因子[7]。Bellanger等[8]通过体内外研究表明,ZNF217是乳腺癌发生骨转移的重要中介和指标。一方面,ZNF217 mRNA表达水平高的原发性乳腺癌发生骨转移的风险更高;另一方面,非侵入性多模态体内成像发现,ZNF217增加了骨转移生长速度,加速了严重的溶骨性病变的发展。此外,ZNF217过表达可驱动异常分化和激活信号通路,促进自我更新能力、间充质标记的表达、运动和转移,并抑制成人组织干细胞基因在癌症中的下调。硅片筛选发现,核苷类似物三西利宾(triciribine)可抑制ZNF217诱导的肿瘤生长和化疗耐药,并可抑制AKT和MAPK等信号通路磷酸化。
本研究为在临床环境中使用三西利宾治疗过表达ZNF217的乳腺癌患者提供理论基础,同时也证明ZNF217可作为乳腺癌进展过程中的潜在预后生物标志物和治疗靶点[9]。
2.2ZEB1(zinc finger E-box-binding homeobox)
ZEB1位于10p11.2号染色体上,是上皮间质转化(epithelial-mesenchymal transition,EMT)相关TF,在间质瘤(如胶质瘤、肺癌、乳腺癌、胰腺癌)中高表达,且与肿瘤侵袭性显著相关[10-12]。ZEB1诱导EMT的作用机制是与上皮细胞标记蛋白E-cadherin的启动子区连接,抑制其转录表达,导致细胞失去上皮性质[13]。Liang等[14]证实,敲除ZEB1可通过下调Flk-1抑制MDA-MB-231细胞血管拟态的形成。透明质酸(hyaluronan,HA)主要由透明质酸合酶2(hyaluronan synthase 2,HAS2)合成,是一种细胞外基质蛋白多糖,在乳腺肿瘤中富集[15]。研究发现,胞外HA与CD44s协同触发ZEB1表达而促进ZEB1介导的EMT,而ZEB1也可直接与HAS2启动子结合并激活其表达,促进更多的HA合成,从而进一步增强ZEB1,形成HA/CD44 s、HAS2和ZEB1正反馈回路[16]。
ZEB1/p53轴作为一种基质特异性信号通路,可促进乳腺上皮肿瘤进展[17]。间质成纤维细胞中ZEB1的缺失可增加成纤维细胞生长因子2/7(fibroblast growth factor 2/7,FGF2/7)、血管内皮生长因子(vascular endothelial growth factor,VEGF)和白细胞介素6(interleukin-6,IL-6)启动子的乙酰化及p53的表达和募集,从而减少其在周围间质中的产生和分泌。而在ZEB1基质缺失的乳腺肿瘤中,p53消融能充分恢复受损的肿瘤生长和进展。
2.3Snail 锌指蛋白Snail是一种E-cadherin表达的转录抑制因子,其通过与上皮性肿瘤抑制基因(cadherin 1,CDH1)结合启动EMT,是EMT的关键诱导因子之一。Maturi等[18]对Snail1在三阴性乳腺癌细胞中进行全基因组染色质免疫沉淀测序(ChIP-seq)分析,发现Snail1通过多种新机制参与乳腺癌细胞的恶性表型。Bossart等[19]证实,它莫西芬(tamoxifen)可诱导Snail抑制侵袭性小叶乳腺癌生长。Snail在细胞迁移和转移中也起着至关重要的作用[20]。具体来说,Snail在乳腺癌细胞中诱导PAPS合酶2(3′-phosphoadenosine 5′-phosphosulfate synthase 2,PAPSS2)和多能聚糖(versican,VCAN)的表达,而PAPSS2的缺失以及PAPSS抑制剂氯酸钠的加入导致MCF7和MDA-MB-231细胞迁移减少,裸鼠肺转移率和微转移结节数量也明显降低。此外,Snail、PAPSS2和VCAN在乳腺癌组织中的表达呈正相关。这些结果体现了Snail1作为转移性乳腺癌治疗靶点的潜力。
3 锌指蛋白在乳腺癌中的作用及机制
锌指蛋白由于其特殊的指状结构,可通过识别并结合DNA、RNA等序列或与蛋白质相互作用,从而实现转录调控。作为人体内重要的TF,锌指蛋白广泛参与乳腺癌的发生发展过程(表1)。

表1 锌指蛋白在乳腺癌中的作用机制
3.1增殖、生长和凋亡 ZFP57(zinc finger protein 57)是KRAB-ZNF家族成员,在胚胎干细胞(embryonic stem cell,ESC)中可以维持DNA甲基化。研究发现,ZFP57为乳腺癌抑制因子,可通过调控MEST启动子区的甲基化,下调MEST介导的Wnt/β-catenin信号通路,从而抑制乳腺癌细胞增殖[21]。Wang等[22]发现,ZNF496通过C2H2结构域与ERα的DNA结合域相互作用,选择性抑制ERα靶基因,最终抑制ERα(+)乳腺癌细胞的生长。Han等[23]的研究表明,ZIC1(zinc finger of the cerebellum 1)抑制乳腺癌细胞增殖,并通过抑制Akt/mTOR/P70S6K通路进而抑制生存素(survivin)表达,促进细胞凋亡。
CTCF(CCCTC-binding factor)是一种11-锌指多功能蛋白,具有转录激活、抑制和染色质屏障活性等分子功能。研究发现,CTCF的缺失致使乳腺癌细胞发生增殖减少、凋亡增加、细胞周期阻滞等表型改变,这与p53的激活密切相关。CTCF作为p53的转录抑制因子,其结合位点位于TP53第一个外显子上游约800 bp处。CTCF的缺失会激活p53及其下游基因p21和Bax(BCL2 associated X),导致细胞周期阻滞和凋亡[24]。
TRPS1(transcriptional repressor GATA binding 1)常在乳腺癌中发生扩增和失活突变。Cornelissen等[25]通过体内外功能丧失的方法分析TRPS1在乳腺发育和侵袭性小叶型乳腺癌中的作用,发现其导致E-cadherin功能丧失。E-cadherin和TRPS1的联合失活导致小鼠乳腺类器官的持续增殖和加速乳腺肿瘤的形成。可见TRPS1在乳腺癌中能够作为一种依赖于环境的肿瘤抑制因子,同时对正常乳腺上皮细胞的生长和分化起着至关重要的作用。
SALL1(spalt like transcription factor 1)是一种调控器官发生和干细胞发育的多锌指TF,可通过介导核小体重构和去乙酰化酶(NuRD)复合物的募集来诱导癌细胞衰老[26]。Cui等[27]发现,Znhit1(zinc finger HIT-type containing 1)可通过上调同源性磷酸酶(phosphatase and tensin homolog,PTEN)致使PI3K/Akt/mTOR通路失活,进而抑制乳腺癌的发生。
综上所述,锌指蛋白主要通过转录调控一系列下游靶基因的形式,激活或抑制Wnt/β-catenin、Akt/mTOR等乳腺癌相关通路,进而影响乳腺癌细胞的增殖、生长和调亡。
3.2侵袭、迁移和转移 Liu等[28]发现,GGNBP2(ZNF403)通过抑制IL-6/STAT3信号通路的激活,进而抑制三阴性乳腺癌的侵袭性。高通量乳腺癌表达数据集分析表明,ZNF367在转移性乳腺癌中特异性过表达,同时高表达ZNF367的患者无转移生存期和总生存期较短[29]。体内外实验表明,ZNF367促进肿瘤转移,同时伴有循环肿瘤细胞(circulating tumor cell,CTCs)数目的增加。由于CTCs的存活与其抵抗失巢凋亡的能力相关,而Hippo通路的中下游效应器YAP1(yes associated protein 1)已证实在乳腺癌细胞中发挥抑制失巢凋亡和促进转移的作用。ZNF367通过招募染色质重构蛋白BRG1激活CIT(citron rho-interacting serine/threonine kinase)和TP53BP2(tumor protein p53 binding protein 2)的转录,而CIT和TP53BP2可以抑制YAP1的磷酸化,进而抑制Hippo通路,激活YAP1信号,并抑制乳腺癌的失巢凋亡和促进CTCs的存活,最终导致乳腺癌的远处转移。Lee等[30]发现,A20在人基底样乳腺癌中显著上调,其表达水平与无转移患者的生存期呈负相关。进一步研究表明,A20可在3个赖氨酸残基上磷酸化Snail1,促进TGF-β1诱导的EMT,从而促进侵袭性基底样乳腺癌的转移。
Osx(Osterix)是一种锌指TF,对成骨细胞分化和骨形成至关重要。Yao等[31]证实,Osx上调与淋巴结转移和患者预后不良有关,且Osx通过上调参与侵袭、血管生成和骨溶解的基质金属蛋白酶9/13(matrix metallopeptidase 9/13,MMP9/13)、血管内皮生长因子(vascular endothelial growth factor,VEGF)、IL-8和甲状旁腺激素样激素(parathyroid hormone like hormone,PTHrP),促进乳腺癌的侵袭和骨溶解转移。Chen等[32]确定了乳腺癌细胞中低氧诱导的表观遗传阅读器ZMYND8(zinc finger MYND-type containing 8)。ZMYND8与低氧诱导因子(hypoxia inducible factor,HIF)相互作用,并通过募集BRD4(bromodomain containing 4)并增加RNA聚合酶Ⅱ磷酸化共激活HIF诱导的癌基因,从而增加血管生成和细胞运动性,减少癌细胞死亡,同时促进乳腺癌ZMYND8被p300乙酰化,乙酰化的ZMYND8对于HIF激活及乳腺肿瘤的进展和转移是必需的。
3.3耐药 ZNF703为常见乳腺癌致癌基因,属于NET/NlZ锌指TF家族成员,其表达上调和细胞凋亡耐药、化疗耐药之间存在联系。Marzbany[33]通过RT-PCR比较ZNF703在人乳腺癌组织、正常乳腺组织和MCF-7乳腺癌细胞株中的表达水平,发现其在乳腺癌组织和MCF-7细胞中的表达分别增加了93.3%和4倍。他们还研究了抗ZNF703 RNAi干扰和布洛芬单独或联合对MCF-7细胞存活和凋亡的抑制作用,结果表明,布洛芬联合抗ZNF703 siRNA比单独治疗更能促进细胞凋亡[33],可见抗ZNF703 RNAi联合布洛芬作为环氧化酶-2的抑制剂,对乳腺癌细胞具有高度抑制作用,显示出治疗乳腺癌的潜力。
ZEB1也是乳腺癌对抗雌激素治疗产生耐药性的关键因素[34]。ZEB1通过在ERα启动子上形成ZEB1/DNMT3b/HDAC1复合物,抑制ERα转录,导致DNA甲基化和ERα沉默。因此,ZEB1会下调ERα表达并减弱雌激素对细胞生长的抑制。大量的乳腺癌标本检测表明,ZEB1与ERα蛋白质表达存在显著的负相关性。此外,乳腺癌高度表达ZEB1抑制ERα启动子甲基化。裸鼠异种移植模型证实ZEB1表达下调可在体内恢复乳腺癌细胞对抗雌激素治疗的反应性。最终结果表明,ZEB1可在乳腺癌中诱发ERα启动子甲基化和授予雌激素抗性。Zhang等[35]发现,ZEB1通过在ATM启动子上形成ZEB1/p300/PCAF复合物,激活ATM激酶表达,增加乳腺癌中同源重组介导的DNA损伤修复和DNA断裂清除,导致化疗耐药。
3.4干细胞特性 肿瘤干细胞因其自我更新和治疗抵抗等特性,被认为是造成肿瘤发生发展、耐药耐辐射和复发的主要原因。Li等[36]发现,ZNF32可通过上调膜相关雌激素受体(G protein-coupled estrogen receptor 1,GPER)的表达,激活ERK信号,诱导干细胞样亚群的扩增和增加耐药性。他们还证明ZNF32通过与GPER启动子结合诱导GPER表达,ZNF32/GPER表达与移植瘤小鼠模型中肿瘤发生率和肿瘤负荷的增加之间存在相关性。综上所述,ZNF32参与GPER/ERK信号传递,并赋予乳腺癌干细胞样特性。Hong等[37]的研究表明,TFRUNX1(RUNX family transcription factor 1)通过抑制干细胞活性和直接抑制ZEB1表达来抑制乳腺癌肿瘤生长。Kuo等[38]分析位点突变,发现Myeloid Zinc finger-1磷酸化驱动骨桥蛋白介导的癌相关成纤维细胞表型,进而增加乳腺癌细胞的干性。
3.5自噬 自噬是细胞内膜结构、蛋白复合物和溶酶体作为溶细胞自噬体降解和更新细胞质成分的主要分解代谢过程,虽然机制尚未完全阐明,但大量研究表明其在乳腺癌中发挥重要作用。ZNF32被认为是一种有效的自噬抑制剂,可保护乳腺癌细胞免受过度刺激-自噬诱导的细胞死亡[39]。乳腺癌细胞中,ZNF143低表达可通过调控p53-Beclin1轴的自噬过程表现出更好的生存力。与普通MCF7细胞比较,经ZNF143敲除的MCF7细胞在饥饿状态下表现出更好的存活率和更多的自噬小泡,且Beclin1、p62和ATGs等自噬相关蛋白也在ZNF143敲除细胞中表达发生改变。此外,ZNF143敲除影响p53的稳定性,表现出对蛋白酶体抑制剂MG132的依赖性。而对ZNF143敲除的乳腺癌细胞进行蛋白质组分析,发现NAD(P)H醌脱氢酶1[NAD(P)H Quinone Dehydrogenase 1,NQO1]对p53的稳定性起作用[40]。
4 总结与展望
作为真核生物TF中的一个大家族,锌指蛋白在多个生物学领域发挥着重要作用。虽然其机制尚未完全阐明,但转录调控的特性已得到广泛证实和初步应用,不仅在基因表达调控等基础研究领域展现了广阔前景,还在临床肿瘤诊断和治疗等领域展露出巨大的潜力。锌指蛋白广泛参与乳腺癌生长、侵袭、转移等一系列发生发展过程:一方面,锌指蛋白可通过锌指结构域招募相互作用蛋白,从而激活或抑制下游基因及相关通路;另一方面,锌指蛋白在不同锌指基序组合下表现出不同的特异性DNA结合能力,从而通过结合直接调控下游基因及通路,影响乳腺癌进程(图1),这为探究一些新型锌指蛋白如何在乳腺癌发生、发展中发挥转录调控功能,为乳腺癌诊断、靶向治疗及耐药,发掘其临床应用价值。此外,随着对锌指蛋白结构和功能研究的不断深入,未来有望通过设计更多人工锌指蛋白来调控相应的基因转录,实现根据需要利用锌指蛋白调控真核生物中基因的表达,为基因治疗开拓新领域。
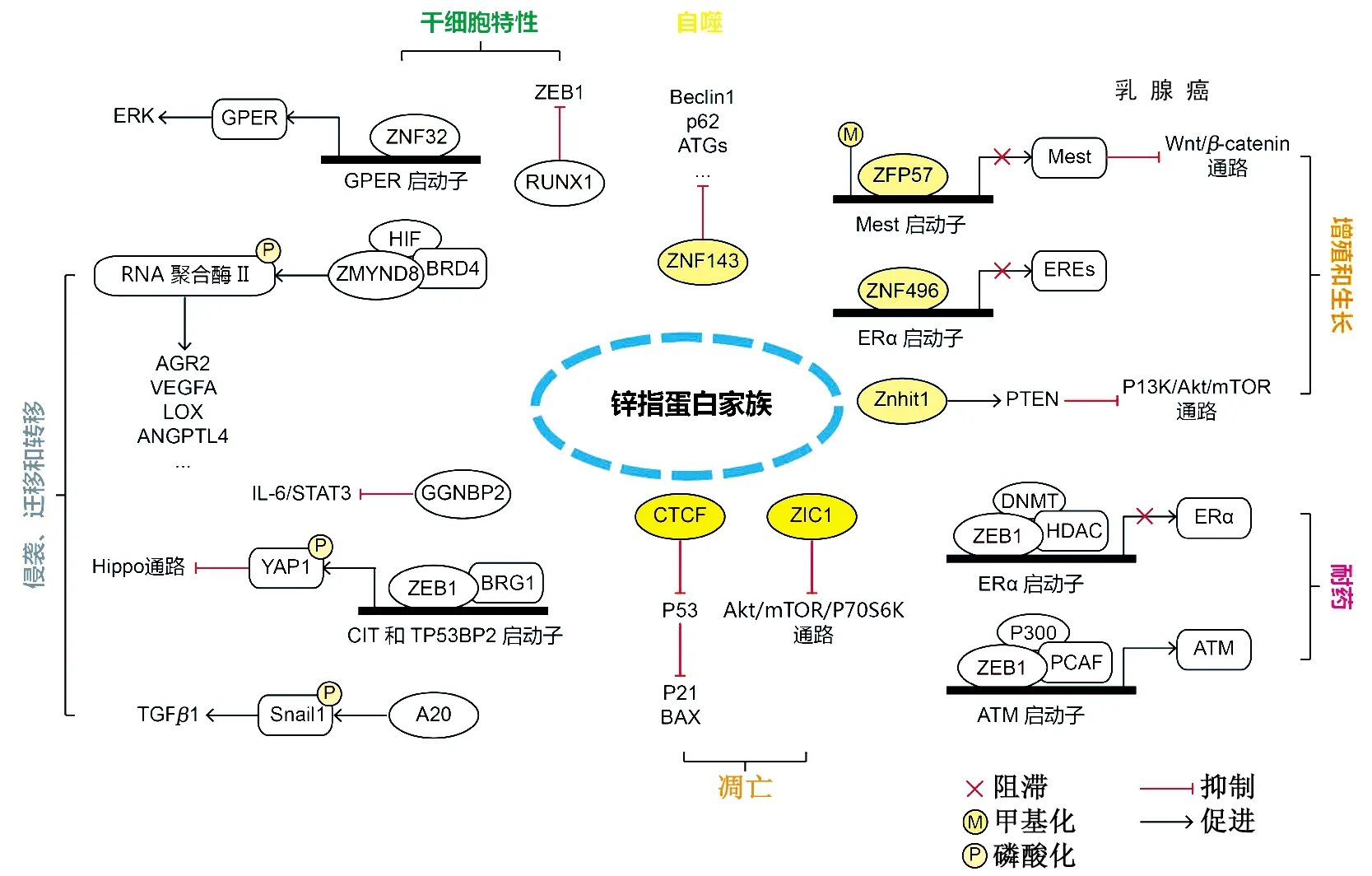
ERK:胞外信号调节激酶;GPER:G蛋白偶联雌激素受体;ZNF:锌指蛋白;ZEB1:转录抑制因子锌指E-盒结合同源异形盒;RUNX1:侏儒相关转录因子1;Beclin1:人卷曲螺旋肌球蛋白样BCL2结合蛋白;ATGs:自噬相关蛋白;Mest:中胚层特异性转录同源蛋白;ERα:雌激素受体α;Znhit1:HIT型锌指蛋白1;PTEN:同源性磷酸酶;DNMT:DNA甲基转移酶;HDAC:组蛋白去乙酰化酶;PCAF:赖氨酸乙酰转移酶2B;ATM:毛细血管扩张性共济失调突变激酶;ZIC1:小脑锌指1;CTCF:CCCTC结合因子;BAX:BCL2结合X蛋白;TGFβ1:转化生长因子β1;Snail1:蜗牛家族转录抑制因子1;CIT:枸橼相互作用丝氨酸/苏氨酸激酶;TP53BP2:肿瘤蛋白p53结合蛋白2;BRG1:染色质重构蛋白1;YAP1:Yes1相关转录调控因子;GGNBP2:配子生成素结合蛋白2;STAT3:信号转换器和转录激活因子3;AGR2:前梯度蛋白2同源物;VEGFA:血管内皮生长因子A;LOX:赖氨酰氧化酶;ANGPTL4:血管生成素样蛋白4;ZMYND8:MYND型锌指蛋白8;BRD4:溴结构域蛋白4;HIF:低氧诱导因子。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28 07:46:10
实用临床医药杂志(2021年13期)2021-01-10 19:52:18
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
作文教学研究(2016年1期)2016-07-05 12:22:47
中国病理生理杂志(2015年8期)2015-12-21 12:38:06
中华胰腺病杂志(2015年5期)2015-12-08 12:18:10
医学研究杂志(2015年8期)2015-06-22 14:00:57
医学研究杂志(2015年3期)2015-06-10 06:41:52
创业家(2015年1期)2015-02-27 07:52:02
