
3种无脊椎动物对近江牡蛎Crassostrea ariakensis和熊本牡蛎C.sikamea的捕食研究
2021-04-25 06:26:50孙兆跃范瑞良隋延鸣唐保军修玉娜李楠楠李雪英全为民
生态学报 2021年7期
孙兆跃,范瑞良,隋延鸣,唐保军,姜 伟,修玉娜,李楠楠,李雪英,全为民,*
1 农业农村部东海渔业资源开发利用重点实验室,中国水产科学研究院东海水产研究所,上海 200090
2 水产科学国家级实验教学示范中心,上海海洋大学, 上海 201300
牡蛎礁(oyster reef)是由大量牡蛎聚集生长所形成的一种生物礁系统,广泛分布于河口和滨海区[1],具有净化水体[2-3]、提供栖息地[4-5]、能量耦合[6-8]和防止岸线侵蚀[9-10]等生态功能。历史上中国沿海分布有大面积的牡蛎礁[11-12],但由于自然海岸带变迁和人为干扰(环境污染、过度捕捞和生境破坏),大部分牡蛎礁已经退化或消失,亟需进行生态修复与保育。
捕食是影响牡蛎种群生存与牡蛎礁发育的重要生物因子之一。目前,国内外学者开展了较多蟹类和螺类捕食牡蛎的相关研究。McDermott[13]测定了扇蟹(Panopeusherbstii)对1-2龄美洲牡蛎(Crassostreavirginica)的捕食效率。Menzel和Nichy[14]比较了石蟹(Menippemercenari)和蓝蟹(Callinectessapidus)对不同规格的美洲牡蛎选择性捕食行为。Brousseau等[15]研究肉球近方蟹(Hemigrapsussanguineus)对3种双壳贝类(蓝贻贝Mytilusedulis、软壳蛤Myaarenaria和美洲牡蛎)的捕食偏好。Kennedy[16]比较了2种蟹类捕食者对美洲牡蛎与近江牡蛎(C.ariakensis)的捕食选择及效率。Newell等[17]观察了1种蓝蟹和4种泥蟹对切萨皮克湾美洲牡蛎和近江牡蛎的捕食偏好。楼小康[18]进行了蛎敌荔枝螺(Thaisechinate)对近江牡蛎的捕食研究。于瑞海[19]开展了脉红螺(Rapanavenosa)对长牡蛎(C.gigas)的捕食实验。这些研究结果表明:牡蛎种类及其规格大小显著影响着消费者的捕食效率。
近江牡蛎和熊本牡蛎(C.sikamea)共同分布于长江口及邻近的苏浙沿岸,既是重要的牡蛎养殖种类,也是沿岸牡蛎礁的造礁物种。日本蟳(Charybdisjaponica)、脉红螺和黄口荔枝螺(T.luteostoma)是该海区野生或养殖牡蛎的主要捕食者。日本蟳属节肢动物门、甲壳纲、梭子蟹科、梭子蟹亚科、蟳属,俗称赤甲红、石蟹等,以底栖动物为食,食物范围广泛[20]。脉红螺隶属于软体动物门、腹足纲、骨螺科,红螺属,为食肉性动物,喜食双壳贝类,通过捕食泥沙中的埋栖型贝类和岩石上的附着型贝类生存[21]。黄口荔枝螺属软体动物门、腹足纲、骨螺科、荔枝螺属,以一些行动迟缓的小型腹足类以及牡蛎、藤壶和苔藓虫等为食[22-23]。
本研究通过室内受控实验测定日本蟳、脉红螺和黄口荔枝螺对不同规格的近江牡蛎和熊本牡蛎的捕食效率,揭示了3种无脊椎动物对不同规格近江牡蛎和熊本牡蛎的捕食偏好性,研究结果可为牡蛎礁的保护和修复提供理论基础及实践指导。
1 材料与方法
1.1 实验材料
本实验于2019年5月在中国水产科学院研究院东海水产研究所水生生物实验室进行。实验用的近江牡蛎和熊本牡蛎均取自上海市芦潮港,带回实验室经清除壳表面的附着物(泥沙和藤壶等)后,将牡蛎放入养殖缸(规格72 cm×44 cm×48 cm)中暂养1周,每日投喂50 mL小球藻溶液(藻细胞浓度5.0×109个/mL)。实验用的日本蟳和脉红螺购于上海市东方水产中心,黄口荔枝螺采自江苏南通东灶港,使用游标卡尺测量日本蟳的甲壳宽(CW)和两种螺类捕食者的壳高(SH)。3种捕食者在室内暂养1周,每天投喂牡蛎1次,实验前24 h停止投喂。每种牡蛎随机选择90个牡蛎,使用游标卡尺测定其壳高和壳厚,壳厚为牡蛎左壳弧形瓣膜壁边缘5mm处的壳体厚度,结果精确到0.01mm。
1.2 实验方法
实验用的被捕食者(近江牡蛎、熊本牡蛎)和捕食者(表1,日本蟳、脉红螺、黄口荔枝螺)均单独放置于57L的循环养殖缸中。日本蟳单只饲养,防止种内残杀。采用室内循环水养殖系统开展实验,实验用水为人工配制的海水,盐度为21—23,水温在(21.0±0.5)℃。本研究开展3个独立实验,每种捕食者与两种牡蛎的捕食关系为1个独立实验。每个实验采用双因子随机区组实验设计,2个自变量为牡蛎种类(近江牡蛎和熊本牡蛎)和牡蛎规格(4组规格),因变量为捕食效率。
适应1周后,挑选体质健康,形态完整的近江牡蛎和熊本牡蛎进行实验。根据壳高,将每种牡蛎划分为4个规格处理组(表2),每个处理组设置5个重复,共计40个实验单元。对于每种牡蛎,每组规格处理挑选50个牡蛎用于实验,测量其壳高、壳厚和体重(湿重)。在每个实验单元中,随机选取相应规格处理组的10只牡蛎和对应的1只捕食者放入1个养殖缸(28 cm×25 cm×23 cm)中进行捕食实验(表1和表2)。每个实验周期为7d,保持充气,每2天换水1次,每天投喂5 mL小球藻溶液(藻细胞浓度5.0×109个/mL)。实验结束时,统计每个实验单元中牡蛎的被捕食比例,即为捕食效率。捕食成功的标准为牡蛎壳打开或者牡蛎壳被钻孔,牡蛎软体部分被捕食。由于实验对照组没有牡蛎死亡,因此在计算捕食效率时不考虑牡蛎自然死亡率。

表1 3种无脊椎动物捕食者的平均规格及体重
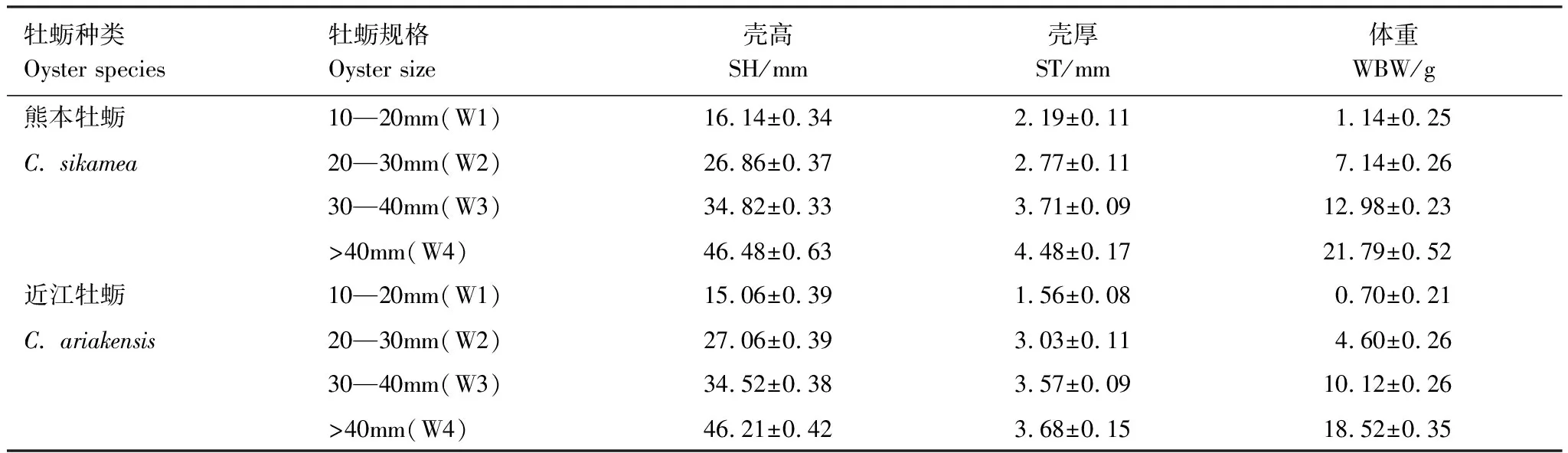
表2 近江牡蛎和熊本牡蛎4个规格处理组的平均壳高(SH)、壳厚(ST)及体重(WBW)
1.3 数据处理及分析
捕食效率表示为平均值±标准误,采用双因子方差分析(Two-way ANOVA)检验2个自变量(牡蛎种类和规格大小)对捕食效率的影响,运用Duncan多重比较检验不同处理间的差异(P<0.05)。利用线性回归检验每种牡蛎的壳厚与壳高之间的相关性,并采用协方差分析检验两组曲线斜率间的差异显著性。数据的统计和分析采用SPSS 25.0进行。
2 结果与分析
2.1 日本蟳对2种牡蛎的捕食效率
双因子方差分析结果表明:日本蟳对2种牡蛎的捕食效率没有显著性差异(表3,F=2.149,P=0.152),但牡蛎的规格大小显著影响着日本蟳的捕食效率(表3,F=5.472,P<0.05),牡蛎种类与其规格大小之间没有显著的互作效应(表3,F=0.543,P=0.657)。
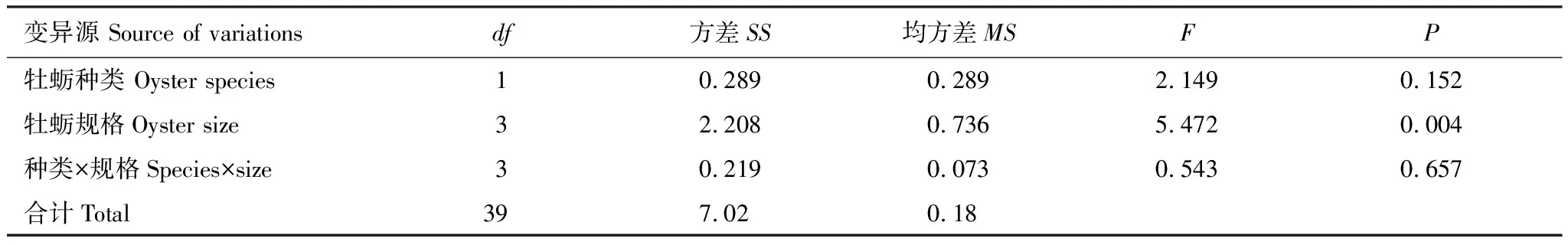
表3 牡蛎种类和规格大小对日本蟳捕食效率影响的双因子方差分析结果
表4显示了日本蟳对4组规格近江牡蛎和熊本牡蛎的捕食效率。据Duncan多重比较结果,日本蟳对W1组近江牡蛎的捕食效率显著高于W2和W4组(表4,P<0.05),对W3组的捕食效率介于中间(表4,P>0.05)。日本蟳对于W1组熊本牡蛎的捕食效率显著高于W2和W3(表4,P<0.05),W4组的被捕食效率与其他3组均没有显著性差异(表4,P>0.055)。
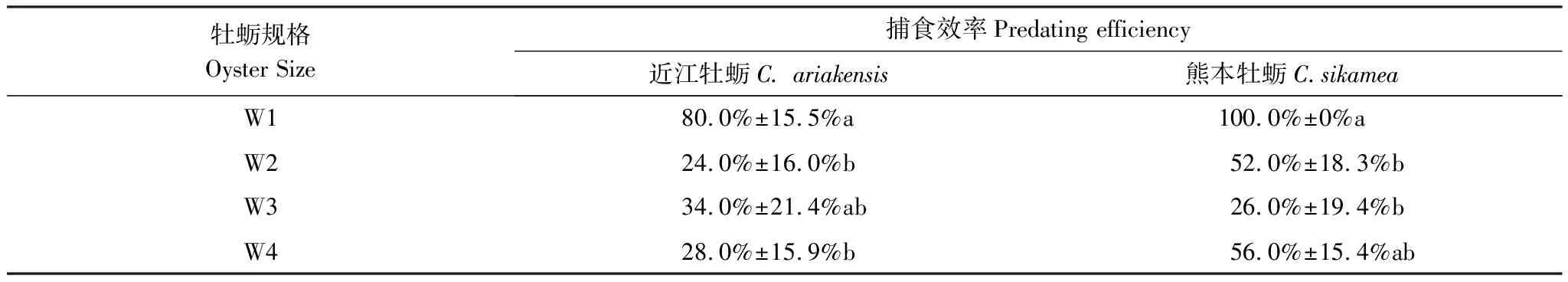
表4 日本蟳对4组规格近江牡蛎和熊本牡蛎的捕食效率(平均值±标准误)
2.2 脉红螺对2种牡蛎的捕食效率
双因子方差分析结果显示:牡蛎种类和规格大小对脉红螺的捕食效率均无显著性影响(表5,种类:F=0.296,P=0.590;规格:F=1.116,P=0.357),牡蛎种类与其规格大小之间没有显著的互作效应(表5,F=0.548,P=0.653)。表6显示了脉红螺对4组规格近江牡蛎和熊本牡蛎的捕食效率,总体上脉红螺的捕食效率很低。
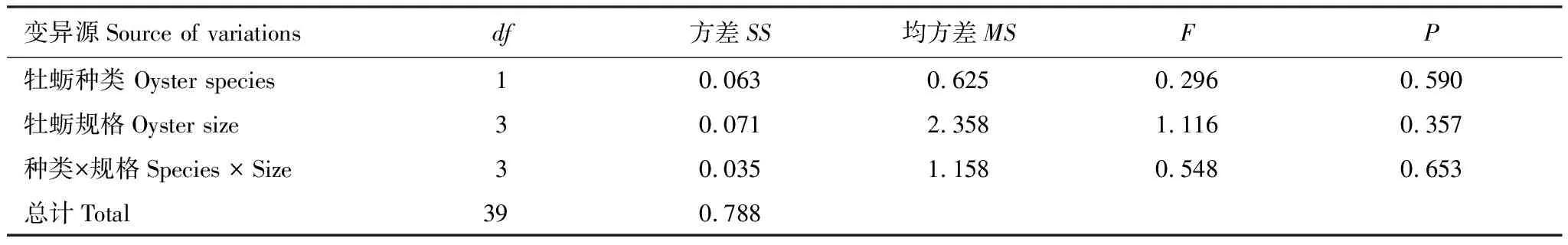
表5 牡蛎种类和规格大小对脉红螺捕食效率影响的双因子方差分析结果
2.3 黄口荔枝螺对2种牡蛎的捕食效率
双因子方差分析结果显示:牡蛎种类对黄口荔枝螺的捕食效率无显著性影响(表7,F=0.011,P=0.917),但规格大小影响着黄口荔枝螺的捕食效率(表7,F=3.232,P=0.035),牡蛎种类与其规格大小之间没有显著的互作效应(表7,F=2.551,P=0.093)。
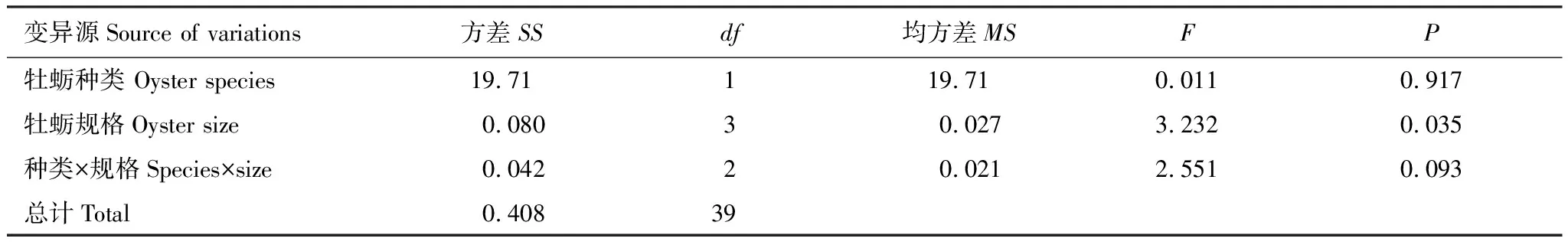
表7 牡蛎种类和规格大小对黄口荔枝螺捕食效率的双因子方差分析结果
表8显示了黄口荔枝螺对4组规格近江牡蛎和熊本牡蛎的捕食效率,总体上黄口荔枝螺的捕食效率很低。据Duncan多重比较结果,黄口荔枝螺对4组规格近江牡蛎的捕食效率没有显著性差异(表8,P>0.05),但对W1组熊本牡蛎捕食效率显著高于其他3个规格组(表8,P<0.05)。
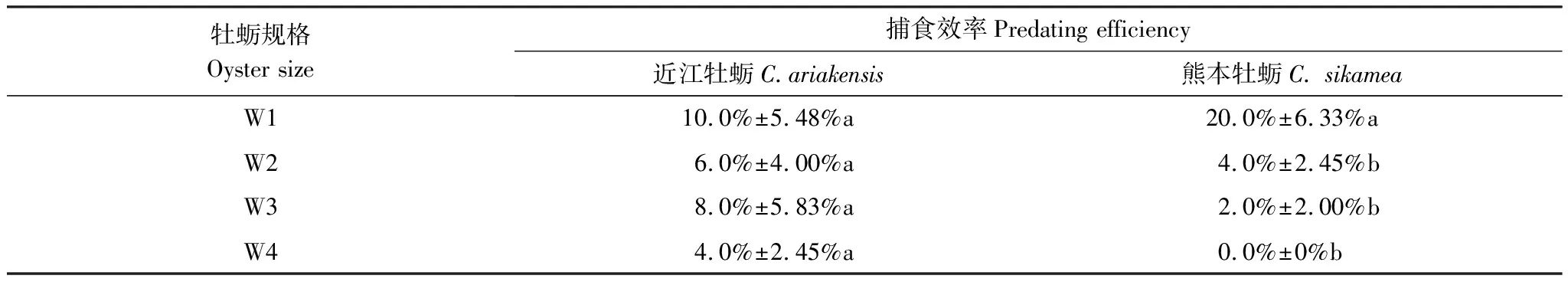
表8 黄口荔枝螺对4组规格近江牡蛎和熊本牡蛎的捕食效率(平均值±标准误)
2.4 牡蛎壳高与壳厚的相关性
2种牡蛎的壳高与壳厚之间均存在极显著的正相关关系(图1,P<0.001)。近江牡蛎壳厚与壳高之间的线性回归方程为ST=1.04+0.074 ×SH(r2= 0.650,P<0.001),熊本牡蛎为ST=1.30+0.051 ×SH(r2=0.45,P<0.001)。两组方程的线性趋势具有显著性差异,两条曲线斜率对数比0.3716(%95置信区间:0.3714—0.3718),同等规格下熊本牡蛎的壳厚显著高于近江牡蛎的壳厚。
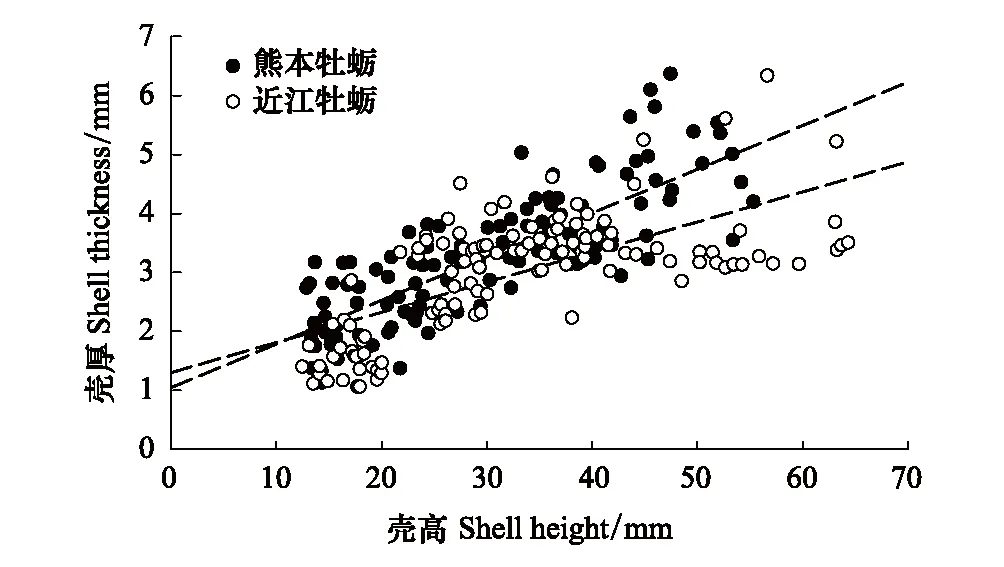
图1 近江牡蛎(CA)和熊本牡蛎(CS)的壳高(SH)与壳厚(ST)之间的线性相关性(样本量n=90)
3 讨论
3.1 3种捕食者对2种牡蛎的捕食行为及选择性
日本蟳捕食牡蛎主要依靠其两个螯足和它的吻,螯足用来捕捉牡蛎、钻孔及破碎牡蛎壳,吻部吸食壳破碎后的牡蛎肉,其捕食方式类似于首长黄道蟹(Cancermagister)[24-26]和以小型河口双壳类为食的珍宝蟹(Calappaphilargius)幼蟹[27]。脉红螺个体较大,用腹足包裹住牡蛎使其窒息,打开贝壳再用消化液将贝肉融化成胶质物质后用吻去吸食;荔枝螺个体小不足以包裹住整个牡蛎,通过分泌酸液腐蚀外壳形成小洞,再往壳内注射消化液使食物消化成液体状态后吸食。两种螺摄食过程的差异可能源自其规格大小的不同,因此采取了两种不同的策略。
Kennedy[16]在实验中发现,两种牡蛎钻(Urosalpinxcinereal,Eupleuracaudata)对美洲牡蛎的捕食效率显著高于近江牡蛎,表现出2种捕食者对美洲牡蛎的偏好捕食行为。然而,Newell[17]的实验研究却发现4种蟹类捕食者更喜欢捕食近江牡蛎而不是美洲牡蛎,原因是近江牡蛎的壳比同规格美洲牡蛎的壳更脆弱,打破近江牡蛎壳所需要的压缩力要比美洲牡蛎低64%;对于牡蛎钻更喜欢捕食美洲牡蛎的机制,他们的解释是牡蛎钻对来自本地美洲牡蛎的化学响应要高于近江牡蛎。赵静[28]研究了有无底质条件下日本蟳对3种贝类(菲律宾蛤仔Ruditapesphilippinarum,光滑河蓝蛤Potamocorbulalaevis及四角蛤蜊Mactraquadrangularis)的摄食选择性,结果显示日本蟳对光滑河蓝蛤有明显的摄食偏好,主要原因是光滑河蓝蛤个体较小,更易于被日本蟳的螯足夹碎取食。于瑞海[19]专门挑选了10种常见双壳贝类活体同时对脉红螺进行投喂,结果表明脉红螺更喜好摄食螠蛏、长竹蛏而不喜欢太平洋牡蛎,此结果表现出脉红螺对壳薄而脆的被捕食者具有偏好性。楼子康[18]的实验结果揭示蛎敌荔枝螺对于近江牡蛎、白纹脊藤壶(Balanusalbicostatus)和中华绿螂(Glauconomechinensis)没有捕食选择性。本研究结果表明日本蟳、脉红螺和黄口荔枝螺均未表现出对同规格的近江牡蛎和熊本牡蛎的摄食选择性,可能的解释是:尽管熊本牡蛎的壳厚显著大于近江牡蛎,但这种壳厚的差异水平还不足以对捕食者的捕食效率造成显著性影响。
3.2 3种捕食者对不同规格牡蛎的偏好与选择性
被捕食者的规格大小通常显著影响捕食者的捕食效率及选择偏好。Brousseau[15]研究表明肉球近方蟹更喜欢捕食规格小的贝类。Savini[29]的研究结果显示脉红螺更倾向于选择捕食小规格的不等壳毛蚶(Scapharcainaequivalvis)。刘吉明[30]等研究发现脉红螺喜食同种饵料生物中的小个体。楼子康[18]的实验证明,荔枝螺先捕食1龄牡蛎,然后再捕食2龄牡蛎。本实验中,牡蛎规格大小对日本蟳和黄口荔枝螺的捕食效率均有显著性影响,小规格组W1(10—20mm)组牡蛎更易被捕食。原因是个体小的牡蛎易于被捕食,捕食过程中捕食者消耗的能量较少。如规格小的牡蛎更容易被蟹钻孔,被螺包住,更利于捕食者用自己的捕食方式进行捕食。相反,对于规格较大的牡蛎来说,不容易用螯足钻孔或者钻孔时间要更长,螺类摄食难度要变大。因此,捕食者不会去花费更多的能量选择大牡蛎,而去选择易于获取,消耗能量较小牡蛎为食。但本研究却发现,脉红螺对4组规格牡蛎的捕食效率均很低,各规格组间的被捕食效率无显著性差异,可能与脉红螺生理生态习性有关,在其人工养殖条件通常表现低摄食行为。
3.3 3种捕食者对2种牡蛎的捕食效率比较
Menzel[14]和Nichy[14]的野外实验结果表明石蟹捕食美洲牡蛎的平均捕食效率为3.7个/d。Savini[29]实验结果显示脉红螺对双壳贝类(湿重约1.2g)平均摄食率约为1 个/d。本实验中,单个日本蟳对近江牡蛎和熊本牡蛎的平均捕食效率分别为0.975个/d和0.725个/d,单个脉红螺对近江牡蛎和能本牡蛎的平均捕食效率分别为0.136个/d和0.229个/d。Menzel[14]实验发现佛罗里达岩螺捕食牡蛎个数较少,在实验中仅有1个牡蛎被捕食,石蟹对牡蛎的捕食效率远高于其他几种捕食者。在本实验中,脉红螺和黄口荔枝螺对近江牡蛎和熊本牡蛎的捕食效率很低,日本蟳的平均捕食效率明显高于脉红螺及黄口荔枝螺,表明日本蟳是该海区牡蛎礁中的主要捕食者。但在实际评估各种捕食者对野外牡蛎种群的捕食压力时,需综合考虑捕食者的密度、规格大小及生态习性等因子。
猜你喜欢
基层中医药(2021年7期)2021-11-02 07:19:54
云南大学学报(自然科学版)(2021年1期)2021-02-05 08:04:04
学生天地·小学低年级版(2019年1期)2019-03-18 02:23:24
学生天地(2019年3期)2019-03-05 08:21:30
太原师范学院学报(自然科学版)(2018年2期)2018-08-17 12:21:50
东华大学学报(自然科学版)(2018年1期)2018-06-29 03:35:24
地震研究(2017年3期)2017-11-06 17:35:54
童话王国·文学大师班(2016年3期)2016-11-10 06:09:49
中外文摘(2016年13期)2016-08-29 08:53:27
大众考古(2015年6期)2015-06-26 08:27:16
