
亚热带森林凋落叶分解过程土壤节肢动物群落的变化特征
2021-04-25 06:26:38刘静如郭可馨游成铭徐振锋
生态学报 2021年7期
刘静如,郭可馨,谌 亚,李 晗,张 丽,游成铭,徐振锋,谭 波,*
1 四川农业大学生态林业研究所,长江上游林业生态工程四川省重点实验室,水土保持与荒漠化防治省级重点实验室, 成都 611130
2 长江上游森林资源保育与生态安全国家林业和草原局重点实验室, 成都 611130
凋落物分解是森林生态系统物质循环和能量流动的核心环节[1]。已有的研究证实,森林生态系统所有植物性养分的再循环过程均通过土壤碎屑食物链能量流动渠道(细菌或真菌)进行[2]。尽管气候和凋落物基质质量是调控全球和区域尺度上凋落物分解速率的主要因子,但是土壤动物群落是构成凋落物分解速率变异的重要生物因素[3-4]。土壤动物群落不仅能通过吞噬、破碎、混合等直接作用改变凋落物形态和表面积加速凋落物养分流动速率[5],而且能通过选择性取食控制土壤微生物丰度、活性和多样性以及繁殖体接种传播间接影响土壤物质循环的速率和方向以及整个土壤养分库营养效率[6-7]。因此,研究土壤动物群落及其对凋落物分解的作用对深入认识地表生态系统物质循环至关重要。
凋落物分解是一个连续变化的过程,这一过程中凋落物基质质量的变化通常会影响参与分解的土壤生物群落,可能导致分解过程中土壤动物群落变化特征出现较大的变异性[8-9]。首先,新鲜凋落物在分解初期易分解组分(如N、P)含量较高,可吸引分布广、适应性强、取食新鲜凋落物的土壤动物参与凋落物的机械破碎,为微生物的定着创造条件,因而该阶段土壤动物群落结构较为简单[10]。其次,随着凋落物质地、基质质量的改变及微生物群落定着繁衍,丰富的食物来源和适宜的栖息生境使以植物碎屑和以微生物为食的土壤动物类群数和个体数显著增加,它们的参与不仅有利于凋落物难分解组分(如木质素、多酚类物质)的降解,而且可增加土壤食物链的复杂性[11-12]。第三,至凋落物分解后期,凋落物基质质量降低,难分解组分比例和腐殖化程度不断提高,不同取食功能类群的土壤动物以及部分稀有类群的参与可能进一步提升碎屑食物链的多样性[12-13]。与此同时,由于凋落物可食性降低,部分土壤动物类群迁移,也可能在一定程度上影响土壤动物群落结构。此外,不同树种凋落物的初始基质质量差异也会对分解过程中土壤动物的数量和群落结构产生影响[14]。然而,迄今为止关于凋落物长期分解过程中土壤动物群落结构变化的关注明显不足。
四川盆地亚热带常绿阔叶林是我国典型的地带性植被分布区。然而,该地区频繁的人类活动干扰致使自然森林植被受到严重破坏,正在被大面积的人工林和次生林逐渐取代[14]。林分结构的变化可影响凋落物归还的数量和质量,土壤动物群落结构和多样性也可能随之改变,进而影响凋落物分解速率和森林生态系统的养分循环[14]。目前,对于该区森林土壤动物群落及其生态功能的研究已有不少报道[8,15-16],主要集中在植被更替、氮沉降和凋落物分解等方面,并未涉及凋落物长期分解过程中土壤动物群落结构变化方面。因此,以亚热带典型阔叶树种麻栎(Quercusacutissima)和针叶树种柳杉(Cryptomeriafortunei)凋落叶为对象,采用分解袋法开展为期四年的凋落叶分解试验,研究不同树种类型凋落叶分解过程中土壤节肢动物群落结构和动态变化,以期为深入认识森林土壤动物与凋落物分解的相互联系与相互作用提供科学参考。
1 材料与方法
1.1 研究区域概况
试验地位于四川省都江堰灵岩山四川农业大学华西雨屏区生态环境监测站(31°01′N, 103°34′E,海拔896—1320 m),属亚热带湿润季风气候,年平均气温15.2℃,最高气温(7月)31.6℃,最低气温(1月)-1.4℃,年均降雨量约1243 mm。该区森林以亚热带常绿阔叶林为主,植物种类繁多,乔木树种主要有麻栎、含笑(Micheliafigo)、喜树(Camptothecaacuminata)、桢楠(Phoebezhennan)、柳杉等;林下灌木层主要有光叶海桐(Pittosporumglabratum)、胡枝子(Lespedezabicolor)、针刺悬钩子(Rubuspungens)和十大功劳(Mahoniafortune)等;草本层以扁竹根(Irisjaponica)、麦冬(Ophiopogonjaponicas)、莎草(Cyperusrotundus)和芒萁(Dicranopterispedata)等为主。土壤为山地黄壤,土层厚度70 cm—100 cm,pH 4.9±0.3,总有机碳(19.8±1.2)g/kg,全氮(1.4±0.2)g/kg,全磷(0.9±0.1)g/kg[14]。
1.2 样地设置和样品布设
2010年11月在监测站内设置1个面积为1 hm2的监测样地。在样地内3条水平线上每隔5 m安装一个面积约5 m2的凋落叶收集器,共15个,定期收集不同乔木树种凋落叶,带回实验室风干后分类。为探讨不同树种类型(针叶vs.阔叶)凋落叶分解过程中土壤节肢动物群落结构和动态变化,于2011年10月凋落叶高峰期后,将风干的麻栎和柳杉凋落叶装入大小为20 cm×20 cm凋落叶分解袋中,分解袋表面(与大气接触面)孔径为3 mm[17],底面(与土壤接触面)孔径为0.04 mm,每袋样品的重量为(10±0.05)g。2011年11月将装袋好的凋落叶分解袋埋置于样地内5个5 m×5 m样方中,共埋设凋落叶分解袋130个(11次采样×5样方×2树种+20备用),埋设深度与地表凋落叶层齐平。分解袋埋置完成后,随即取5袋凋落叶带回实验室,用以测定凋落叶初始质量特征(表1)。分解袋埋设的同时,分别在凋落叶分解袋和样地内距离地面1.5 m放置纽扣式温度传感器(DS1923-F5#, Maxim/Dallas semiconductor Inc., USA)连续监测凋落物和大气温度[15]。传感器1 h记录1次数据每隔2月用电脑下载一次数据。
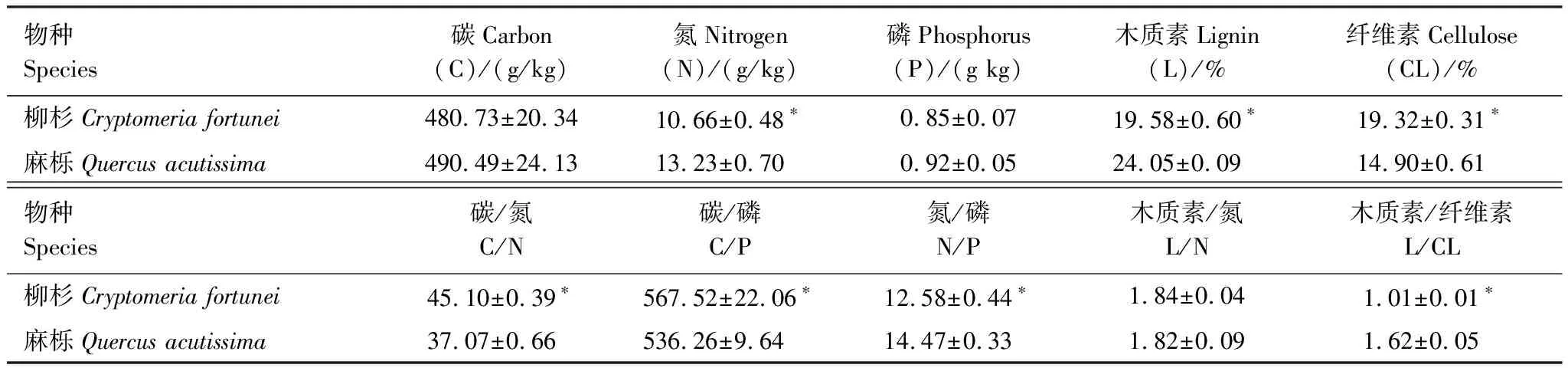
表1 柳杉和麻栎凋落叶初始质量特征(平均值±标注误差, n=5)
1.3 样品采集与土壤动物分离
样品采集于2011年12月27日、2012年4月27日、8月25日、12月26日、2013年4月24日、8月13日、12月27日、2014年4月27日、10月26日、2015年4月24日和10月26日进行,共11次采样,与之对应的凋落叶分解阶段为35 d、156 d、277 d、398 d、516 d、628 d、746 d、896 d、1079 d、1261 d、1444 d。采样时,从每个样方中采集分解袋2袋,将分解袋装入透气的黑布袋中低温保存,12 h内带回实验室。采用Tullgren漏斗(4 mm)分离收集的土壤节肢动物,分离周期为48 h。采用体式解剖镜和生物显微镜镜检计数和分类,参考《中国土壤动物检索图鉴》[17]、《昆虫分类检索》[18]鉴定到科水平。
1.4 数据统计与分析
根据捕获个体数量占捕获总量百分比,将个体数量10.0%以上的类群划分为优势类群,1.0%—10.0%的类群划分为常见类群,小于1.0%的类群划分为稀有类群[17]。由于土壤动物区系种类繁多、食性和营养级类型复杂多样,且取食食性易随季节和生境条件变化,将其系统分类为某一特定的功能类群难度较大。因此,基于已有的文献资料和各类群的总体特征将捕获的土壤动物划分为捕食性、植食性、菌食性、腐食性四个功能类群[14,19]。
采用Shannon-Wiener多样性指数(H)[20]、Pielou均匀度指数(J)[21]、Simpson优势度指数(C)[22]、Margalef丰富度指数(D)[23-24]计算残留凋落物中土壤动物群落多样性随分解进程的变化特征,计算公式如下:
式中,S为土壤动物群落的类群数;Pi=ni/N,ni为每个类群的个体数,N为总的个体数。
独立样本T检验(Student′s independent-sample t-test)凋落叶初始基质质量以及同一采样时期两个树种凋落叶土壤节肢动物个体密度、类群数量和多样性指数的差异;重复方差分析(repeated measures ANOVA)检验凋落叶树种、采样时间及其交互作用对土壤节肢动物群落特征的影响,统计分析由IBM SPSS 22.0(IBM SPSS Statistics, Chicago, IL, USA)完成。采用Cluster聚类和非度量多维尺度方法(non-metric multidimensional scaling analysis)对比两种凋落叶土壤节肢动物群落结构,检验分析由R软件(version 3.5.2)完成。
2 结果
2.1 土壤节肢动物组成随凋落叶分解的变化特征
整个分解期间,共捕获土壤节肢动物3855只,隶属16目51科(表2和表3),其中麻栎凋落叶中1924只,柳杉凋落叶中1931只。2种凋落叶中土壤节肢动物的优势类群均为弹尾目,麻栎凋落叶以等节跳科(Isotomidae)和棘跳科(Onychiuriclae)为优势类群,柳杉凋落叶以等节跳科,棘跳科和圆跳科(Sminthuridae)为优势类群;它们共有的常见类群有隐翅甲科(Staphylinidae)、摇蚊科(Chironomidae)、土革螨科(Ologamasidae)、蚁科(Formicidae)等10科,稀有类群有蚁甲科(Pselaphidae)、长角毛蚊科幼虫(Hesperinidae)、爪跳科(Paronellidae)、三甲螨科(Oribotritiidae)、原铗虫八科(Projapygidae)等14科。稀有类群长须螨科(Stigmaeidae)、赤螨科(Erythraeidae)、隐颚螨科(Cryptognathidae)等8科仅在麻栎凋落叶中检出,长角跳科(Entomobryidae)、罗甲螨科(Lohmanniidae)、狼蛛科(Lycosidae)等11科只在柳杉凋落叶中捕获。
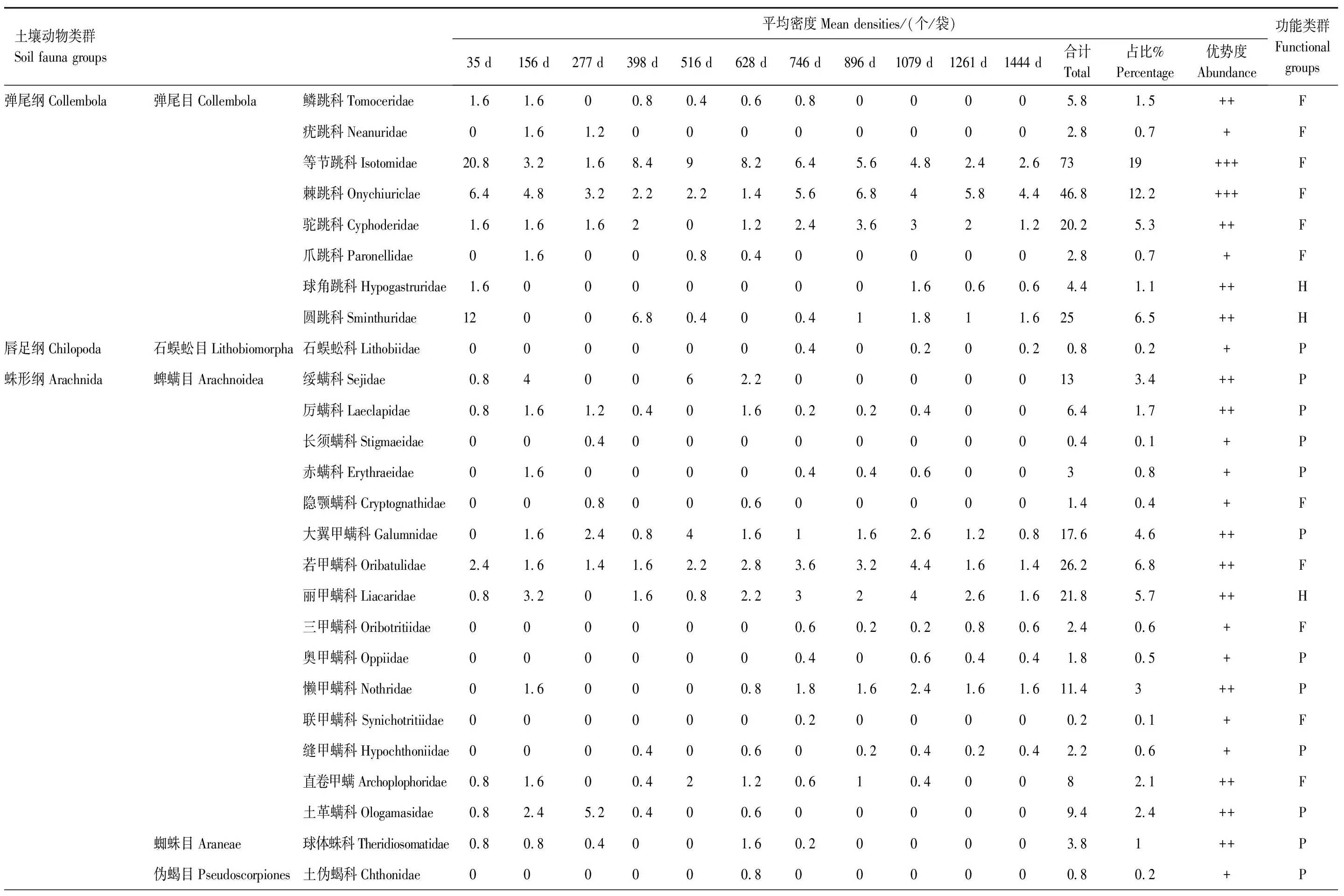
表2 麻栎(Quercus acutissima)凋落物中土壤节肢动物组成及动态变化
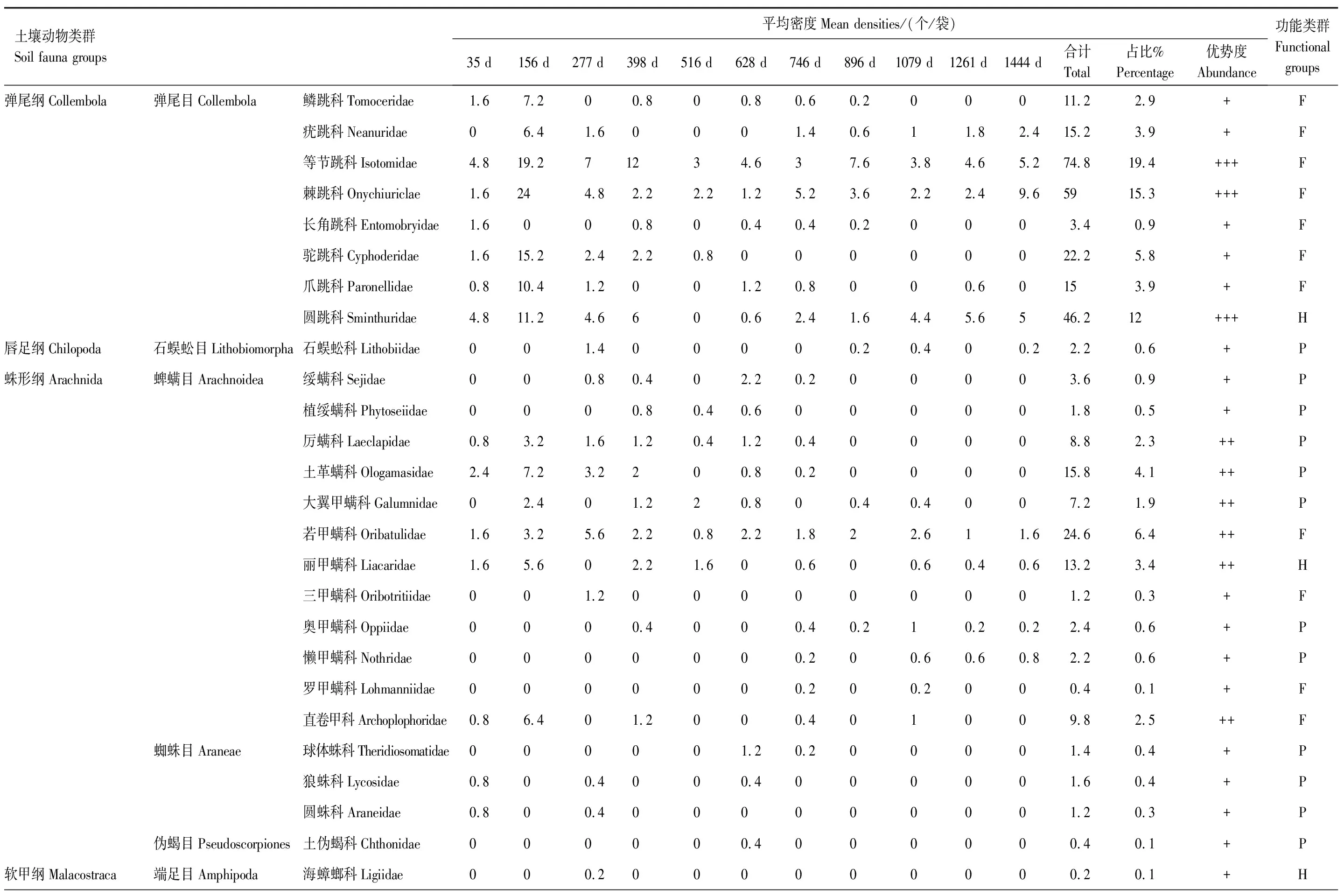
表3 柳杉(Cryptomeria fortunei)凋落物中土壤节肢动物组成及动态变化
凋落叶分解袋埋设后,土壤节肢动物快速定殖,个体密度和类群数量随凋落叶分解进程不断变化(图1)。麻栎凋落叶土壤节肢动物的个体密度随分解进程呈现增加趋势,在分解的1079 d达最高值后降低;而柳杉凋落叶则在分解的156 d急剧增加后快速降低。2类树种凋落叶土壤节肢动物类群数量具有相似的动态变化过程,但麻栎凋落叶土壤节肢动物类群数量显著高于柳杉凋落叶,麻栎和柳杉凋落叶土壤节肢动物类群数量分别在分解的746 d和1079 d达到最高值。凋落叶种类、分解时间及其交互作用均显著影响土壤节肢动物的个体密度和类群数量。整个分解阶段,麻栎凋落叶土壤节肢动物总体上以常见类群占比最高,而柳杉凋落叶土壤节肢动物以优势类群占比最大(图2)。柳杉凋落叶中稀有类群占比高于麻栎凋落叶,且随着分解进程,柳杉凋落叶中优势类群占比呈现上升趋势。
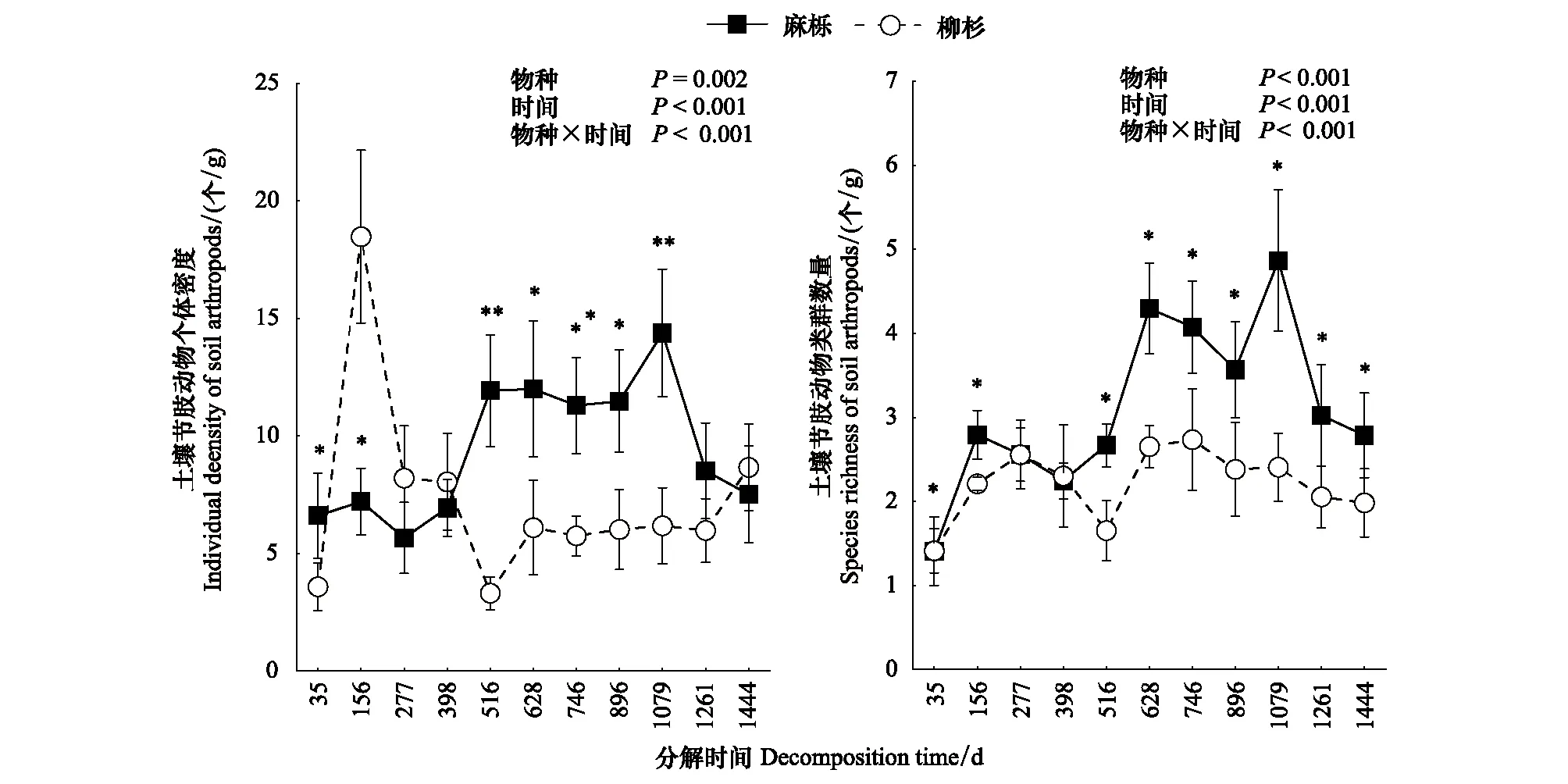
图1 麻栎和柳杉凋落叶分解过程中土壤节肢动物个体密度和类群数量动态(平均值±标准误,n=5)
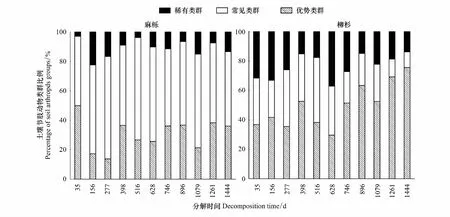
图2 麻栎和柳杉凋落叶分解过程中土壤节肢动物类群(优势类群、常见类群、稀有类群)动态(平均值±标准误,n=5)
2.2 土壤节肢动物功能类群随凋落叶分解的变化特征
土壤节肢动物功能类群随凋落叶类型和分解进程变化明显不同(图3)。两种凋落叶中捕获的土壤节肢动物以菌食性占比最高,腐食性最低,且随凋落叶分解进程,植食性土壤节肢动物占比明显下降,菌食性则上升。在麻栎凋落叶分解过程中,捕食性土壤节肢动物比例仅在277 d有明显降低,植食性土壤节肢动物在分解的277 d数量占比最大。柳杉凋落叶分解过程中,捕食性土壤节肢动物占比明显波动,在分解的628 d占比最低,与之相反植食性土壤节肢动物占比在分解的628 d增至最大值后明显降低。
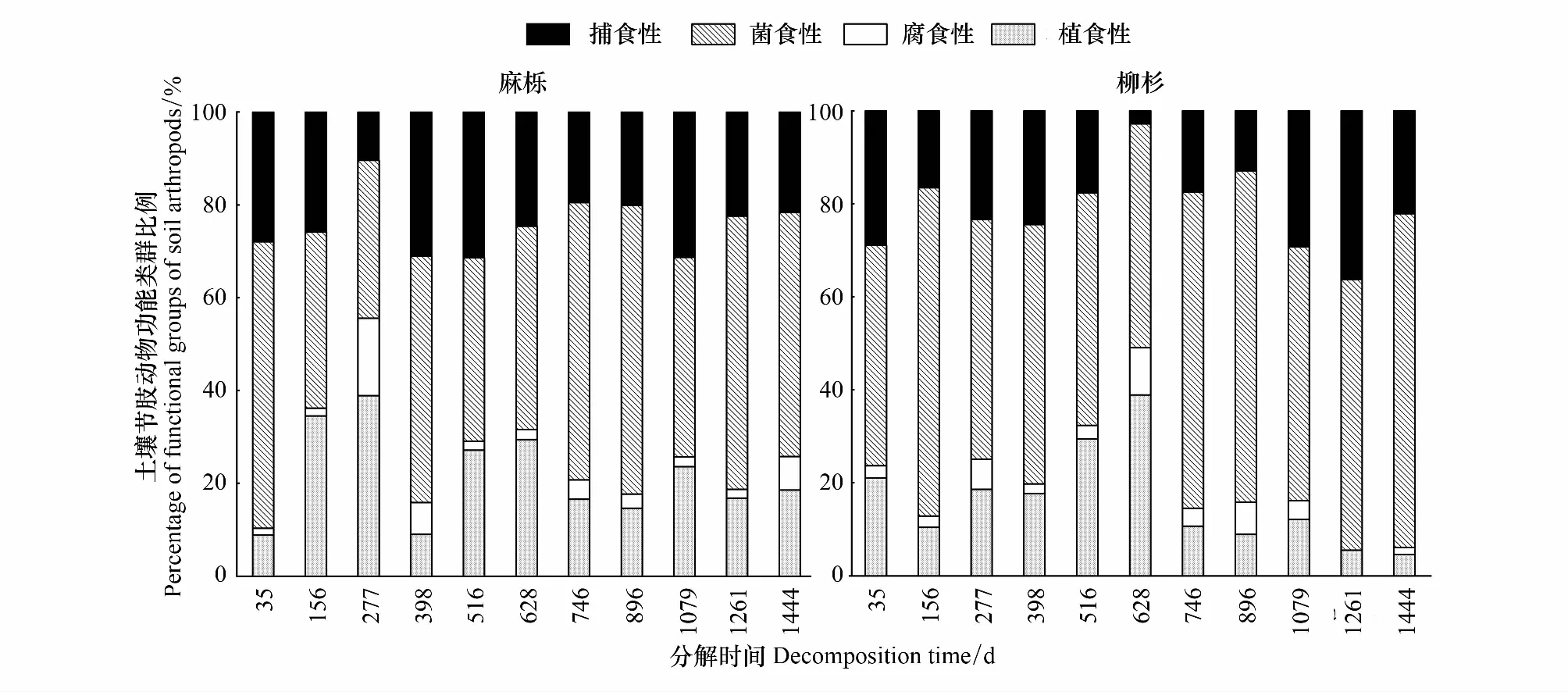
图3 麻栎和柳杉凋落叶分解过程中土壤节肢动物功能类群(捕食性、菌食性、腐食性和植食性)动态(平均值±标准误,n=5)
2.3 土壤节肢动物多样性随凋落叶分解的变化特征
由图4可见,两种凋落叶分解过程中土壤节肢动物多样性总体呈现出相似的动态变化。除均匀度指数外,2种凋落叶土壤节肢动物优势度、多样性和丰富度指数差异显著(P<0.05)。在多数时期,柳杉凋落叶中捕获的土壤节肢动物优势度指数大于麻栎,总体呈上升趋势,多样性指数与丰富度指数都呈现下降趋势,且多数时期麻栎大于柳杉,均匀度变化波动不大。非度量多维尺度(NMDS)分析表明,两种凋落叶土壤节肢动物群落组成具有显著差异(P=0.02),聚类分析显示,两种凋落叶土壤节肢动物群落结构随分解进程相似性不断降低(图5)。

图4 麻栎和柳杉凋落叶分解过程中土壤节肢动物多样性指数动态(平均值±标准误,n=5)
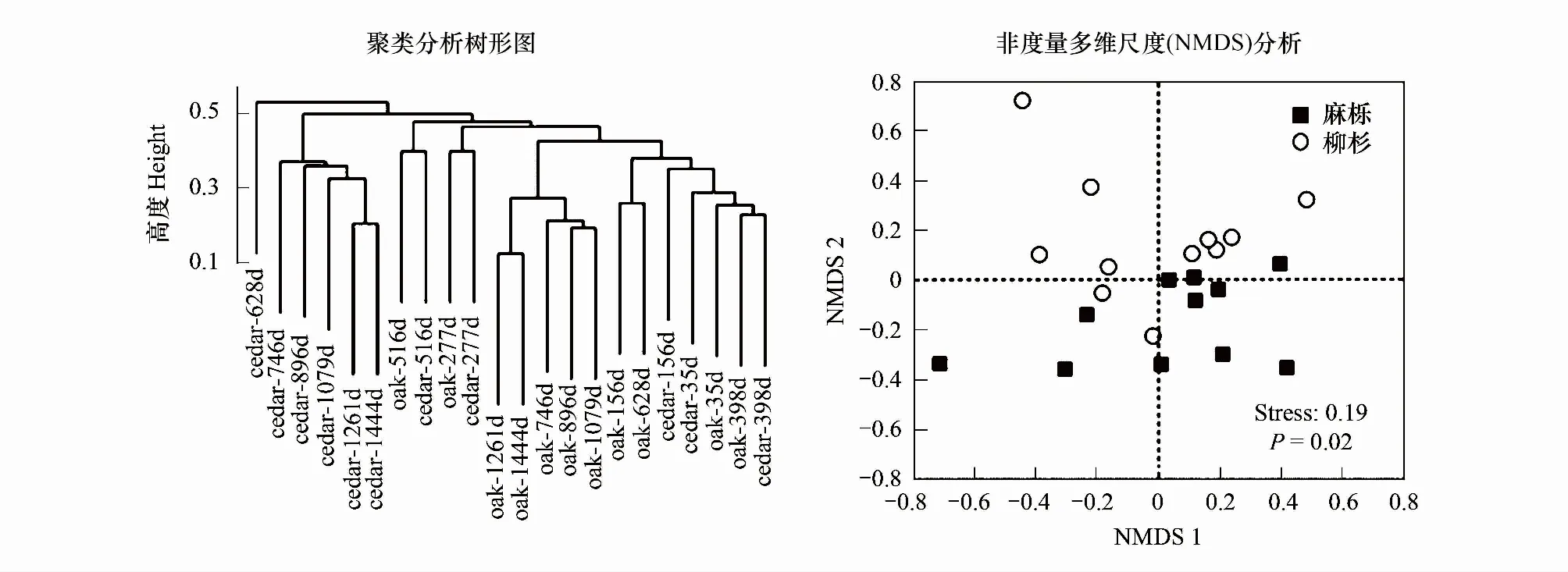
图5 麻栎和柳杉凋落叶分解过程中土壤节肢动物组成Cluster聚类和非度量多维尺度(NMDS)分析
3 讨论
土壤动物通过取食、破碎、搬运等方式参与凋落物分解,是构成凋落物分解速率变异的重要生物因素[25]。研究表明,土壤动物群落空间自相关性随类群和生境不同,其组成和结构随凋落物类型和基质质量变化而异[26]。本研究中,柳杉和麻栎凋落叶分解袋中捕获的土壤节肢动物总数量无显著差异,土壤节肢动物类群组成与我国亚热带/热带森林研究结果较为相似[27-28]。整个研究期间,捕获土壤节肢动物以蜱螨目和弹尾目为优势类群(表2和表3),这与易兰等[29]在亚热带森林中的研究结果一致,这是由于它们取食真菌、细菌以及植物凋落物碎屑,少部分也可以捕食其他小型土壤动物,食性相对复杂的原因[30]。2种凋落叶中捕获的土壤节肢动物个体密度和类群数量随分解时期不同差异显著,麻栎凋落叶的土壤节肢动物个体密度和类群数量在分解后期显著高于柳杉,这与短期凋落物分解的研究结果存在不一致性[10,31]。通常,针叶凋落物的C/N、Lignin/N、萜类及酚类物质含量比阔叶凋落物高,这种凋落物基质质量会降低了自身的分解速率,也会限制土壤动物群落在凋落物中的生长和繁衍[17,32-33]。同时,Bradford等[34]研究指出,C/N值是决定凋落叶分解速率及参与分解土壤生物丰度的重要因子。与麻栎凋落叶基质质量相比,柳杉凋落叶高的C/N值可能是造成参与凋落物分解土壤节肢动物密度和类群数量降低的原因(表1)。值得注意的是,两种凋落叶获得的优势类群和常见类群相似度较高,但稀有类群差异较大,如赤螨科、球角跳科、叶蝉科仅在麻栎凋落叶中出现,长角跳科、罗甲螨科、海蟑螂科仅出现在柳杉凋落叶中。这是因为不同稀有类群土壤动物取食特性和获得食物源不同的原因,而分解过程中食物源和凋落叶基质特性变化也会导致稀有类群组成和数量的变化。
不同土壤动物类群的体形大小和活动能力相差悬殊,但它们在土壤生态系统中常常占据着相同的生态位[19],所以目前还无法将所有土壤动物类群的生态习性细化分类,而是按各类群的总体特征将土壤动物划分成不同的营养级,如本研究中的腐食性、植食性、菌食性和捕食性[19,35]。已有的研究表明[12,29],土壤动物功能类群在凋落叶分解初期以菌食性为主,中期以植食性和菌食性为主,后期食物链和功能类群复杂化,将导致捕食性和腐食性类群的增加。这在本研究中也得到较好的证实,两种凋落叶分解过程中土壤节肢动物功能类群总体以菌食性为主,其次是捕食性和植食性,腐食性最少(图3)。同时,植食性土壤节肢动物比例在麻栎凋落叶分解初期(35—277 d)迅速上升,而其在柳杉凋落叶中至分解的516 d才显著增加,这与王小平等[36]的短期研究结果不一致。麻栎凋落叶质地柔软,初始养分含量高等特征更易于植食性土壤节肢动物快速定着,使得土壤动物的破碎作用在分解前期就开始发挥作用[37]。相比之下,柳杉凋落叶的针叶表面革质,角质层发达,且含有较多难分解物质(木质素、纤维素、单宁等),阻碍了微生物和土壤动物的定着和繁殖[35],且淋溶作用也不及阔叶,所以土壤动物前期的取食破碎作用在35 d后开始受到限制,植食性类群在分解中期才开始大量出现。
物种多样性指标能反应群落组成的复杂程度以及群落稳定性[31]。本研究发现,凋落叶长期分解过程中,树种、时间及两者的交互作用对凋落叶中捕获的土壤节肢动物多样性指数、优势度和丰富度指数影响显著。丰富度指数是表征土壤生境优越与否的参数[28],在两个树种间该指数存在明显差异,说明两者分解过程中的微生境条件差异较大。在柳杉和麻栎凋落叶分解过程中,土壤节肢动物丰富度指数总体呈现下降趋势,但个别分解时期(如分解的156 d,628 d和1261 d)柳杉凋落叶丰富度指数显著低于麻栎凋落叶,与类群密度变化一致。这主要是针叶凋落物在分解初期速率缓慢,土壤动物类群种类较少,且主要以菌食性为主的原因。多样性指数和均匀度的高低可以体现生态系统食物链长短和共生现象的多少,从而分析该生态系统群落结构的稳定性[38]。本研究中,不同树种间土壤节肢动物多样性指数的差异主要集中在分解后期,柳杉叶土壤节肢动物的多样性指数在分解一年后迅速降低,这与短期的研究结果明显不同[39]。而非度量多维尺度(NMDS)和聚类分析显示,两种凋落叶土壤节肢动物群落组成和结构在分解过程中具有显著差异,相似程度随分解进程不断降低(图5)。这种土壤动物组成和结构在分解过程中的差异可能是造成不同物种或类型凋落叶分解速率变异的重要因素。值得注意的是,土壤节肢动物的均匀性指数在两种凋落叶中差异不显著,说明两类树种土壤动物群落结构的稳定性表现较为一致。此外,优势度指数是在群落类群组成基础上进一步推算出来以表达群落组成状况的指标,优势度指数高就表明群落内不同类群的分布越不均匀,相比之下,优势类群就更能发挥自身的生态功能[40]。柳杉凋落叶的土壤节肢动物优势度在分解过程中逐渐增加,这与柳杉的优势类群占比变化一致,但与短期的研究相比[14],两类凋落叶的优势度总体较低,表明研究区域土壤节肢动物优势类群的生态功能可能未得到充分发挥,针叶凋落物的降解可能更加依赖某些特定土壤动物类群的作用。
综上所述,土壤节肢动物群落的组成和结构随凋落叶分解过程不断变化,且与短期分解的研究结果存在明显差异。土壤节肢动物的稀有类群组成、群落结构和多样性因凋落叶类型和基质质量差异而不同。这些结果表明凋落物分解过程中土壤动物的群落结构和多样性与凋落物类型密切相关,土壤动物群落结构差异是构成不同类型凋落物分解速率差异的重要生物因素。
猜你喜欢
大自然探索(2024年1期)2024-03-19 19:01:03
广东蚕业(2022年4期)2022-11-23 13:31:55
安徽农学通报(2022年6期)2022-04-07 21:30:29
科学大众(中学)(2021年3期)2021-05-17 17:07:01
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
浙江林业(2020年7期)2020-08-06 14:55:32
课外生活(小学1-3年级)(2018年10期)2018-11-19 07:26:10
中成药(2018年3期)2018-05-07 13:34:25
喜剧世界(2016年7期)2016-08-31 06:21:06
