
miR-885-3p抑制胃癌细胞的增殖和迁移
2021-03-26 09:22李亚洲韩瑞旸卫飞鹏
南昌大学学报(医学版) 2021年1期
李亚洲,韩瑞旸,卫飞鹏
(1.宝鸡高新人民医院介入疼痛科,陕西 宝鸡 721000; 2.兵器工业五二一医院肝胆与血管外科,陕西 西安 710065; 3.中国空军军医大学唐都医院介入放射科,陕西 西安 710038)
胃癌是常见的消化道恶性肿瘤,在我国胃癌发病率位于所有恶性肿瘤第2位,死亡率位于第3位,其主要治疗方式为手术治疗;但是晚期胃癌仍然缺乏有效的治疗方式,即使积极采取各种治疗方式,其五年生存率仍然低于30%[1-2]。由于胃癌细胞生长、增殖快且易转移,严重影响患者生命健康[3]。因此找到阻断胃癌进展的分子靶点,对挽救患者生命具有极大的意义。
近年来,microRNA(miRNA)的发现为肿瘤的治疗提供新的解决途径和思路。miRNA是一类内生的、长度约为20~24个核苷酸的小RNA,由发夹结构的双链RNA前体剪切而形成,在细胞内具有多种重要的调节作用。miRNA可以和靶基因3′非翻译区(3′UTR)互相结合,在转录后对目的基因的翻译产生影响,调控目的基因表达,对不同的分子信号通路进行调控,进而影响细胞增殖、分化、凋亡、炎症和应激反应,单个miRNA可对数个目的基因产生作用,而数个miRNA也可以作用于相同的目的基因,形成复杂的调控网络[4-5]。miRNA还能作为肿瘤抑制因子或致癌基因影响肿瘤发生、发展、侵袭和转移[6-7]。
miRNA-885-3p是由miRNA-885的3′端剪接而成的。有研究[8]表明:miRNA-885-3p的表达程度可能与肺癌耐药性、胶质瘤进展等有关,但是miRNA-885-3p对于胃癌增殖和迁移影响未见报道。因此,本研究通过比较miRNA-885-3p在胃癌细胞及正常细胞的表达,探讨miRNA-885-3p对胃癌细胞增殖和迁移的影响。
1 试剂与方法
1.1 试剂及仪器
人胃癌细胞系SGC-7901、MGC-803、BGC-823、AGS以及人胃正常黏膜细胞GES-1均购自中国生物化学与细胞生物学研究所。Lipofectamine TM 2000以及逆转录试剂盒(TaKaRa公司,批号:1708113、A180302),PCR引物序列由浙江格鲁斯特生物科技公司合成。青霉素和链霉素(伊力特公司,批号:1805161、1809012),基础培养基和胎牛血清(Biological Industries公司),酶标仪(Invitrogen公司),CCK-8细胞计数检测试剂盒(CST公司),Trizol试剂(TaKaRa 公司),Transwell小室 (Corning 公司),Matrigel基质胶(BD 公司,批号:356234),CCK-8 试剂(同仁公司,批号:180109),CO2细胞培养箱、荧光定量PCR仪(Thermo公司),倒置显微镜(重庆奥特光学仪器有限公司),IDTE 缓冲液(Integrated DNA Technologies公司)。
1.2 细胞培养与转染
将人胃癌细胞系SGC-7901、MGC-803、BGC-823、AGS以及人胃黏膜细胞GES-1细胞株,置于添加青霉素100 U·mL-1和链霉素100 mg·mL-1的含10%胎牛血清RPMI 1640培养基中,并在5%CO2、37 ℃、饱和湿度环境中培养。
选取对数生长期的SGC-7901细胞胰酶消化后,用基础培养基调整细胞密度为1×106个·mL-1,将细胞接种到6孔板后37 ℃孵育24 h,按照制造商说明书分别使用Lipofectamine TM 2000将mimic或miRNA-NC转染SGC-7901细胞,分为miR-mimic组和miR-NC组,检测转染效率,并将未经转染的SGC-7901细胞株作为空白对照组。
1.3 qRT-PCR检测miRNA-885-3p表达
使用qRT-PCR分别检测SGC-7901、MGC-803、BGC-823、AGS、GES-1细胞miRNA-885-3p表达;并检测miR-mimic组、miR-NC组及空白对照组miRNA-885-3p表达。miRNA-885-3p及U6的逆转录引物按文献[7-8]报道设计,由浙江格鲁斯特生物科技公司合成。
选择状态良好的细胞,当细胞生长密度达到80%~90%时,弃去培养基,用PBS溶液清洗2~3次后将1 mL Trizol试剂加入到每个孔中,室温条件下裂解5 min。将细胞裂解液转移至1.5 mL EP管中,每管加入200 μL氯仿(4 ℃预冷30 min),剧烈振荡15 s混匀,室温下静置10~15 min。将混合液在4 ℃、2000 r·min-1离心15 min,液体明显分层,将上层水相转移至1.5 mL EP管中,加入等体积的异丙醇(4 ℃预冷30 min),-20 ℃静置30 min。将混合液置于4 ℃、2000 r·min-1离心10 min,弃上清,加入适量75%乙醇(4 ℃预冷,DEPC水配制)重悬RNA沉淀。4 ℃、7500 r·min-1将重悬液离心5 min,弃上清晾干,以20 μL灭菌RNase-free水溶解,并适当吹打均匀。紫外分光光度法检测RNA浓度和纯度。
qPCR反应体系:25 μL SYBR Premix Ex Taq Ⅱ,20 ng cDNA和上下游引物(表1)共40 μmol·L-1,ddH2O补充至50 μL。TaKaRa试剂盒逆转录RNA为cDNA。将1 μg总RNA反转录为cDNA后用IDTE 缓冲液(pH=8.0)稀释至10 ng·L-1。依据qRT-PCR说明书进行检测,反应条件为:95 ℃预变性30 s;95 ℃变性5 s;60 ℃ 45 s; 70 ℃ 45 s,共40个循环。相应miRNA表达量检测以参考基因(U6)进行归一化处理。实验重复6次。结果以2-△△Ct方法计算[9]。
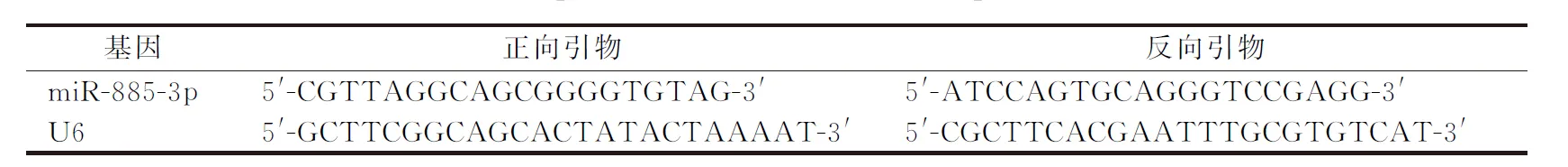
表1 qRT-PCR检测miRNA-885-3p的序列
1.4 细胞增殖能力检测
采用CCK-8检测各组不同时间点细胞活力,细胞转染后48 h,将miR-mimic组、miR-NC组及空白对照组细胞分别消化,以每孔3000个细胞种于96孔板内,每个分组设立5个复孔。分别在24、48、72、96 h进行CCK-8测量。测量时每孔加入100 μL培养基和10 μL CCK-8试剂混合液,并增加空白对照孔,放入细胞孵箱内培养1 h后,在避光条件下,酶标仪检测450 nm波长处的光密度值,并记录分析。以上实验重复6次。
1.5 细胞迁移能力检测
采用伤口愈合实验检测各组迁移能力,6孔板中接种miR-mimic组、miR-NC组,空白对照组细胞株,细胞密度调整为5×105个·孔-1,细胞单层铺满后用PBS液冲洗1次,然后用10 μL枪头尖端在6孔板细胞上垂直划痕,PBS液冲洗2~3次,之后继续培养24 h,用显微镜观察0 h和24 h的细胞划痕宽度并拍照。实验重复6次。
1.6 细胞侵袭能力检测
各组侵袭能力采用Transwell实验检测。将冻存的基质胶提前融化,然后以1:8 的比例将其与不含血清的Matrigel基质胶混匀,每个小室加入100 μL 混合液,放入37 ℃孵箱内孵育24 h。3组细胞消化离心,制成细胞悬液,调整细胞密度为3×108个·L-1。Transwell小室上室加入200 μL细胞悬液,下室加入700 μL含血清的完全培养基。细胞培养箱中培养24 h,取出小室后用PBS轻轻洗涤,棉签擦去边缘和内侧细胞。之后用4%多聚甲醛固定并晾干,加入0.1%结晶紫染色,固定时间20 min,染色时间15 min,PBS清洗掉染色液后自然风干,显微镜下拍照。实验重复6次。
1.7 统计学方法

2 结果
2.1 miRNA-885-3p在胃癌细胞中低表达
SGC-7901、MGC-803、BGC-823、AGS胃癌细胞株中miRNA-885-3p表达分别为(0.21±0.03)(0.46±0.06)(0.38±0.04)(0.42±0.05),均显著低于GES-1细胞的(1.24±0.33),差异有统计学意义(P<0.05),见图1。

*P<0.05与GES-1比较。图1 胃癌细胞和正常胃黏膜细胞miRNA-885-3p表达水平比较
2.2 miRNA-885-3p过表达抑制胃癌细胞增殖
miR-mimic组、miR-NC组、空白对照组SGC-7901细胞miRNA-885-3p表达水平分别为(7.27±1.22)(1.02±0.05)(1.05±0.07),miR-mimic组miRNA-885-3p表达水平显著高于miR-NC组、空白对照组(P<0.05),见图2。CCK-8检测结果显示,miR-mimic组SGC-7901细胞活力显著低于空白对照组和miR-NC组(P<0.05),见图3。

2.3 miRNA-885-3p过表达抑制胃癌细胞迁移
miR-mimic组、miR-NC组、空白对照组SGC-7901细胞迁移距离分别为(0.27±0.04)(0.52±0.06)(0.51±0.05),miR-mimic组SGC-7901细胞迁移距离显著低于miR-NC组、空白对照组(P<0.05),见图4—5。

2.4 miRNA-885-3p过表达抑制胃癌细胞侵袭
miR-mimic组、miR-NC组、空白对照组SGC-7901侵袭细胞数分别为(115±15)(212±27)(203±25),miR-mimic组SGC-7901侵袭细胞数显著低于miR-NC组、空白对照组(P<0.05),见图6—7。

*P<0.05与miR-mimic组比较。图6 3组SGC-7901侵袭细胞数比较
3 讨论
胃癌是世界范围内常见的恶性肿瘤,是我国癌症死亡的主要原因之一,其致病分子机制非常复杂且相关研究较少[10-11]。胃癌晚期患者通常采取癌症组织的扩大根治性切除,同时辅助化疗或放化疗。据文献[12]报道:晚期胃癌患者具有高复发率、低生存率的特点。因此,探索胃癌增殖、转移的分子机制,寻找特异性标志物或建立准确、低危害的治疗策略,已成为临床胃癌研究重点。
非编码RNA(ncRNAs),包括miRNAs和long ncRNAs(lncRNAs),约占基因组的98%,已被发现在生理和病理条件下参与蛋白编码基因的调控[13-14]。miRNAs通过调控多种信号通路,在细胞生长和凋亡、细胞分化、胰岛素分泌、胚胎后期发育等诸多重要的生理过程中发挥调控作用[15]。其中,恶性肿瘤的发生可能与miRNA的异常表达相关,miRNA表达异常引起目的基因表达水平上调或下调,进而调控肿瘤的发生和进展,涉及的可能机制包括对细胞增殖、细胞凋亡、细胞周期、细胞内酶、药物转运等的异常调控。近年,miR-885-3p被报道参与了多种癌症的发病机制,但是其作用在不同的肿瘤并不一致[16-18]。miR-885-3p在肉瘤细胞中,通常呈过表达,但其具体分子机制尚未明确;在涉及γ-突触蛋白的癌症中已经证明上调mir-885-3p可导致其发生[19]。与此相反,miR-885-3p在结肠癌中可通过靶向BMPR1A和阻断BMP/Smad/Id1信号通路来阻断血管生成,在乳腺癌中通过靶向免疫调节蛋白B7-H337来发挥抑癌作用[20-21]。但是miRNA-885-3p与胃癌的调控网络关系及其在癌变发展过程中的作用仍未涉及。因此,本研究通过比较研究miRNA-885-3p在胃癌细胞及正常细胞的表达,为胃癌发病机制研究提供新的靶点。
本研究qRT-PCR检测结果发现,较正常人胃黏膜细胞,miRNA-885-3p在SGC-7901、MGC-803、BGC-823、AGS胃癌细胞中呈低表达。而在miR-mimic组的细胞中miRNA-885-3p呈高表达。伤口愈合实验、transwell实验、CCK-8检测各组细胞增殖、迁移能力,发现miR-mimic组细胞增殖、迁移能力均显著低于空白对照组和miR-NC组。这提示miRNA-885-3p可能是胃癌的抑制基因,可能成为胃癌潜在的治疗靶点。miRNA-885-3p过表达,可能抑制胃癌细胞增殖、迁移。这可能与周期蛋白依赖性激酶-4(CDK4)有关,CDK4在各肿瘤细胞中均有上调,主要集中在细胞生长,可促进肿瘤发生、进展、迁移[22-23]。如在乳腺癌中miR-885-3p表达水平的增加可降低CDK4的表达,进而起到抑癌作用[24]。近年来研究[25-26]发现:miRNA或lncRNA均能通过CDK通路调控乳腺癌、卵巢癌进展,通常可导致细胞增殖延迟、细胞周期G1/S期阻滞或细胞凋亡增强。但是本研究仍然存在一定的局限性,如在胃癌细胞中,miRNA-885-3p过表达影响胃癌进展的具体机制,需要进一步的研究阐述。
猜你喜欢
理财周刊(2022年4期)2022-04-30
智慧农业导刊(2022年7期)2022-04-21
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年1期)2022-02-28
抗癌(2021年3期)2021-11-30
昆明医科大学学报(2021年5期)2021-07-22
昆明医科大学学报(2021年5期)2021-07-22
看世界·学术下半月(2020年7期)2020-09-10
学苑创造·A版(2020年12期)2020-01-07
中国医药导报(2018年3期)2018-03-07
