
三疣梭子蟹14-3-3 基因的克隆及其在低盐和病原胁迫后的表达分析*
2021-03-19 06:13题兴斌吕建建阎德平孙东方
渔业科学进展 2021年1期
题兴斌 吕建建 宋 柳 阎德平 孙东方 刘 萍①
(1. 上海海洋大学水产与生命学院 上海 201306;2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071;3. 农业农村部海洋渔业可持续发展重点实验室 中国水产科学研究院黄海水产研究所 青岛 266071)
三疣梭子蟹(Portunus trituberculatus)是我国重要的海水养殖经济蟹(戴爱云等, 1977)。盐度是三疣梭子蟹养殖中的重要环境因子,直接影响其生长、发育和存活(隋延鸣等, 2012)。夏季暴雨多发,养殖水体盐度急剧下降,使得三疣梭子蟹体内渗透压失衡,导致其生长缓慢、成活率较低(孙东方等, 2018)。病害发生也是影响三疣梭子蟹养殖的重要问题,其中,对虾白斑综合征病毒(WSSV)和副溶血弧菌(Vibrio parahaemolyticus)是导致三疣梭子蟹死亡的主要病原(Marques et al, 2011; Sullivan et al, 2017)。耐低盐和抗病性状是三疣梭子蟹重要的育种性状,深入研究其低盐适应和免疫防御的分子机制对培育三疣梭子蟹耐低盐及抗病良种具有重要意义。
14-3-3 基因是一种广泛分布于所有真核生物中的高保守基因(Chaudhri et al, 2003),编码一组酸性可溶的多功能蛋白,由Moore 等(1967)首次在牛脑细胞中发现。14-3-3 基因的功能具有多样性,参与生物细胞应激、免疫、细胞周期调控、细胞信号转导、代谢酶合成、细胞凋亡等多种生理过程的调控(Koskinen et al, 2004; Angrand et al, 2006; Neal et al, 2009; Matta et al, 2012; Rehman et al, 2014; Gómez-Suárez et al,2016; Lu et al, 2017; Yang et al, 2017)。由于其功能的重要性,该基因在拟穴青蟹(Scylla paramamosain)、凡纳滨对虾(Litopenaeus vannamei)、斑节对虾(Penaeus monodon)等多种经济甲壳类动物中已被克隆,比如在盐度胁迫过程中,斑节对虾中14-3-3 基因大量表达(Kaeodee et al, 2011);在病原感染过程中,凡纳滨对虾中14-3-3 基因呈显著上调表达(Wang et al, 2007),初步证明了该基因在抗逆、抗病过程中具重要作用。至今尚未见该基因在三疣梭子蟹中的相关研究报道。
本研究以三疣梭子蟹为对象,通过本实验室三疣梭子蟹转录组测序得到三疣梭子蟹14-3-3 基因片段,对14-3-3 基因在不同组织、低盐和病原胁迫下的表达情况进行研究。本研究利用RACE 技术,首次克隆三疣梭子蟹14-3-3 基因,通过qRT-PCR 分析其在三疣梭子蟹不同组织中的表达水平,并对其在盐度胁迫和病原胁迫下的表达变化规律进行研究,以此探讨Pt14-3-3 在三疣梭子蟹盐度适应和免疫反应中的功能,为三疣梭子蟹耐低盐和抗病品种培育提供依据。
1 材料与方法
1.1 实验材料
本实验所需三疣梭子蟹均取自中国水产科学研究院黄海水产研究所实验基地潍坊昌邑海丰水产有限公司,体重为(5.78±1.11) g。暂养于4 m×5 m×1.5 m的室内水泥池中,每池90 只,共10 池,暂养7 d。养殖水温为(25±1)℃,盐度为33,pH 8.7,持续充氧,每天换水1/3,于18:00 定时投喂蓝蛤(Potamocorbula laevis)。随机选取9 只暂养后健康有活力的三疣梭子蟹,分别取其血细胞、心脏、肝胰腺、鳃、肌肉和眼柄,存于液氮,设3 个平行,每个平行3 只,用于组织表达分布分析。
1.2 实验方法
1.2.1 盐度胁迫实验 随机挑取暂养7 d 的三疣梭子蟹,分为2 组,对照组(33)和低盐组(11)(72 h 半致死盐度)(Han et al, 2014; 隋延鸣等, 2012),每个处理组设3 个平行,每个平行60 只。使用自然海水和淡水配制低盐度实验用水,使用YSI 盐度仪进行盐度校准,具体方法见隋延鸣等(2012)。各组分别于胁迫后0、3、6、12、24、48 和72 h 取鳃和肝胰腺组织,液氮速冻保存,用于RNA 的提取,每个时间点取3 只。
1.2.2 病原胁迫实验 随机挑取暂养7 d 后体表完整、活力旺的三疣梭子蟹,分成副溶血弧菌注射组和WSSV 注射组,每组50 只,设3 个平行。分别于梭子蟹的游泳足第一关节基膜处注射浓度为3.8×108CFU/ml副溶血弧菌和7.6×107CFU/ml WSSV(谢建军等, 2011;Ren et al, 2017)。在人工感染后的0、3、6、12、24、48 和72 h 取血细胞和肝胰腺组织,于液氮中速冻保存,用于RNA 提取,每个时间点取3 只。
1.3 Pt14-3-3 基因全长cDNA 的克隆及测序
取三疣梭子蟹鳃、肝胰腺、肌肉等组织样品,采用TRIzol®Reagent(Invitrogen 公司)方法进行总RNA提取,经 1%琼脂糖凝胶电泳和紫外分光光度计(NanoDrop 2000, Thermo)对所提RNA 检测质量和浓度,将各组织的高质量RNA 均匀混合,使用SmartTMRACE Amplification Kit (TaKaRa 公司)进行3′和5′RACE cDNA 模板的合成。根据从三疣梭子蟹cDNA文库中获得的Pt14-3-3 基因的EST 序列,利用Primer Premier 5.0 软件设计3′RACE 和5′RACE 特异性引物及RACE 通用引物UPM(表1-1)。使用Advantage 2 PCR Kit(Clontech 公司)进行3′和5′末端的扩增。将获得的PCR 产物回收纯化、连接转化,挑取阳性单克隆,利用M13-47/48 引物进行菌落PCR 鉴定,筛选目的菌液进行测序。
1.4 序列生物信息学分析
利用Vector NTI 11.0 软件对测序后的序列进行冗余序列去除和再拼接,通过DNAStar 的EditSeq 程序进行开放阅读框(ORF)的预测和氨基酸的翻译。三疣梭子蟹Pt14-3-3 基因的核苷酸序列和推导氨基酸序列使用 BLAST(http://www.blast.ncbi.nlm.nih.gov/Blast.cgi)进行同源性比对。利用ProtParam tool 软件进行蛋白质理化性质预测,使用InterproScan 软件进行蛋白质功能结构域分析,TMHMM 2.0 软件进行蛋白质跨膜区分析。使用ClustalX 软件对三疣梭子蟹Pt14-3-3 基因的氨基酸序列与其他物种的14-3-3 氨基酸序列进行多序列比对,在此基础上采用MEGA 4.0软件,以邻接法构建系统进化树。
1.5 Pt14-3-3 的组织表达及各实验组的表达特征
利用Trizol 试剂提取不同实验组三疣梭子蟹鳃和肝胰腺组织的总RNA,使用PrimeScript RT reagent Kit 反转录合成cDNA。根据已获得的三疣梭子蟹内参基因β-actin 和Pt14-3-3 基因全长序列,利用Primer Premier 5.0 软件设计 2 对正反引物(β-actin-F 和β-actin-R;Q14-3-3-F 和Q14-3-3-R)(表1),使用SYBR Premix Ex Taq Ⅱ试剂在ABI 7500 Real Time PCR 仪上对不同处理组获得的Pt14-3-3 基因进行实时荧光定量检测。荧光定量PCR 反应体系为20 μl:10 μl SYBR Premix Ex TaqTMⅡ (2×),0.8 μl 10 μmol/L 的引物Q14-3-3-F,0.8 μl 10 μmol/L 的引物Q14-3-3-R,0.4 μl ROX Reference Dye Ⅱ(50×),2.0 μl cDNA 模板,6.0 μl 无菌ddH2O。反应程序:95℃ 30 s,95℃5 s,60℃ 34 s,40 个循环;95℃ 15 s,60℃ 1 min,95℃ 15 s。采用2-ΔΔCt计算Pt14-3-3 基因的相对表达量,使用SPSS 17.0 软件进行单因素方差(One-way ANOVA)分析。

表1 本研究中所用引物序列Tab.1 The sequence of primers used in this study
2 结果与分析
2.1 Pt14-3-3 基因cDNA 全长克隆及序列分析
获得的三疣梭子蟹基因全长为2510 bp,包括93 bp的5′端非编码区,1676 bp 的3′非编码区和744 bp 的开放阅读框(ORF)(图1)。3′端含有PolyA 尾,具有AATAAA 多聚腺苷酸加尾信号。氨基酸序列分析可知,Pt14-3-3 基因编码一个由247 个氨基酸组成,理论等电点为4.65,分子量为27.98 kDa 的蛋白。由于其较高的不稳定系数(41.28)和较低的GRAVY(Grand average of hydropathicity) (–0.657),推断其为不稳定的亲水蛋白。
2.2 Pt14-3-3 基因的同源性及系统进化分析
利用MEGA 4.0 软件进行系统进化分析显示,所有物种的14-3-3 共聚为两大类群:脊椎动物和无脊椎动物;在无脊椎动物中,三疣梭子蟹与中华绒螯蟹(Eriocheir sinensis)紧密聚为一支,同源性为94%,之后的聚类顺序依次为与脊尾白虾(Palaemon carinicauda)、日本对虾(Marsupenaeus japonicus)、拟穴青蟹、斑节对虾、墨吉明对虾(Fenneropenaeus merguiensis)。在脊椎动物和无脊椎动物中,14-3-3 基因在氨基酸序列上都具有高度的保守性,表明该基因在物种间非常保守。
2.3 Pt14-3-3 基因的组织表达
利用实时荧光定量 PCR 分析了三疣梭子蟹Pt14-3-3 基因在不同组织中的表达分布特征,结果显示,Pt14-3-3 基因在鳃、肝胰腺、肌肉、眼柄、血细胞和心脏中均有表达。其中,在肝胰腺中的表达量最高,其次为肌肉、鳃、心脏和眼柄,而在血淋巴中的表达量最低(图4)。
2.4 Pt14-3-3 基因在低盐胁迫中的表达
Real-time PCR 检测了三疣梭子蟹在低盐胁迫后不同时间点的鳃和肝胰腺中Pt14-3-3 基因的相对表达情况(图5)。低盐胁迫后,Pt14-3-3 基因在鳃中的相对表达量与对照组相比,于3~24 h 呈现显著下调表达,48~72 h 为显著上调表达,分别于6 h 和48 h达到最小值和最大值,为对照组的0.24 倍(P<0.05)和1.34 倍(P<0.05)。低盐胁迫后,Pt14-3-3 基因在肝胰腺中的相对表达量与对照组相比,除6 h 外,整体呈上调表达,表达趋势为先上升、下降后再上升、下降,于12 h 达到最大值,为对照组的7.54 倍(P<0.05)。

图1 三疣梭子蟹14-3-3 基因核苷酸序列及其推导的氨基酸序列Fig.1 Nucleotide sequence and deduced amino acids sequence of P. trituberculatus 14-3-3 gene
2.5 Pt14-3-3 基因在病原胁迫中的表达
实时荧光定量PCR 检测了三疣梭子蟹在病原胁迫后不同时间点的肝胰腺和血细胞中Pt14-3-3 基因的相对表达情况(图6)。感染副溶血弧菌后,Pt14-3-3基因在肝胰腺中相对表达量与对照组相比,整体呈显著上调表达,分别于12 h 和48 h 达到最大值和最小值,为对照组的17.52 倍(P<0.05)和1.77 倍(P<0.05);在血细胞中的相对表达量与对照组相比,除6 h 外,整体呈下调表达,分别于6 h 和12 h 达到最大值和最小值,为对照组的1.68 倍(P<0.05)和0.04(P<0.05)倍。感染WSSV 后,Pt14-3-3 基因在肝胰腺和血细胞中的相对表达量与对照组相比,除24 h 外,整体均呈上调表达,分别于72 h 和3 h 达到最大值,为对照组的13.35 倍(P<0.05)和3.25 倍(P<0.05),均于24 h 降至最小值,分别为对照组的0.52 倍(P<0.05)和0.42 倍(P<0.05)。
3 讨论

图2 三疣梭子蟹Pt14-3-3 氨基酸序列与其他物种14-3-3 氨基酸序列比对Fig.2 Amino-acid sequences alignment of P. trituberculatus with different animals´ 14-3-3 genes
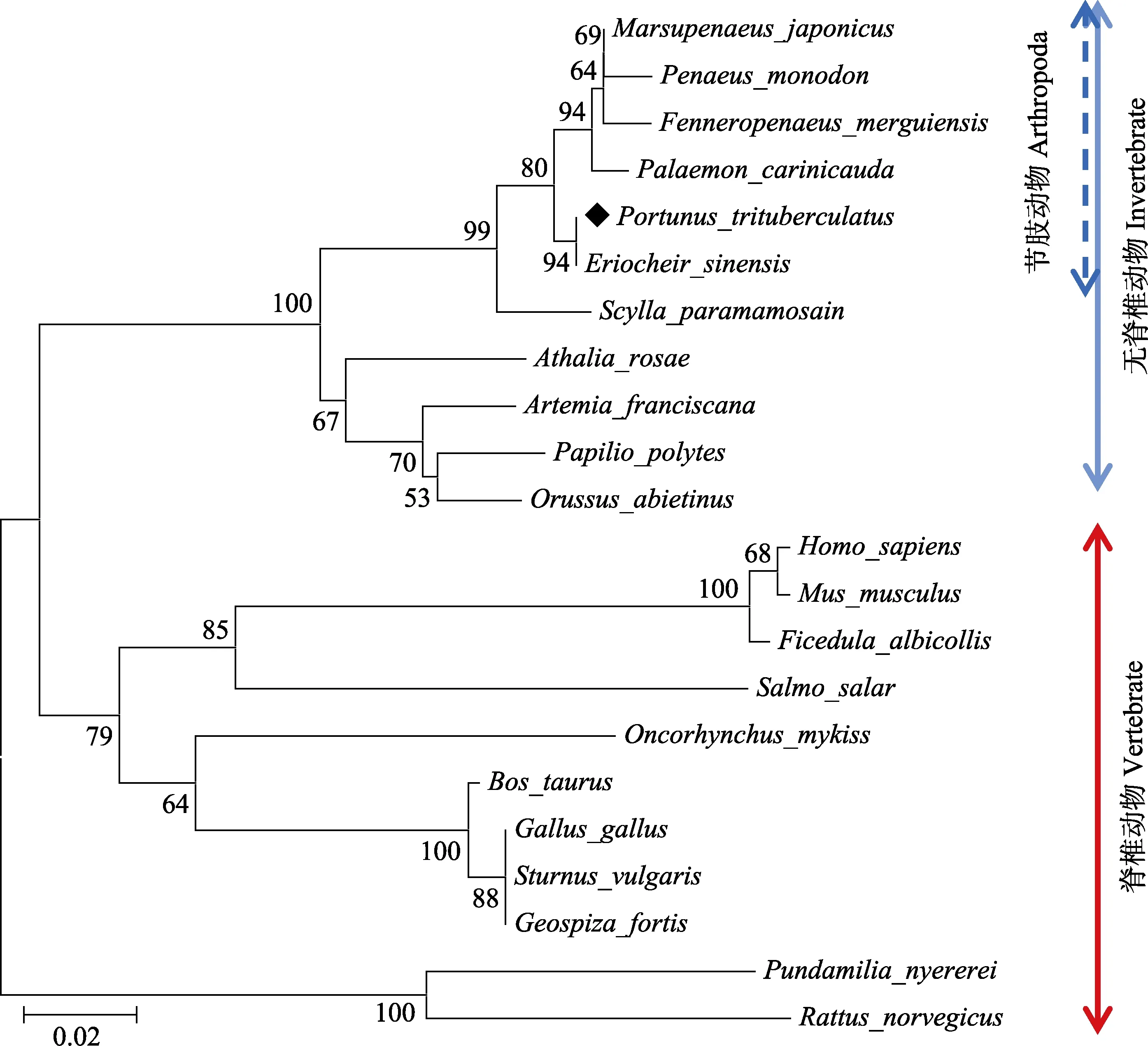
图3 利用MEGA 4.0 软件构建的基于14-3-3 氨基酸序列的NJ 系统进化树Fig.3 NJ tree based on 14-3-3 gene amino acid sequences using MEGA 4.0

图4 三疣梭子蟹Pt14-3-3 基因在各组织中的表达Fig.4 Distribution of Pt14-3-3 gene expression in different tissues of P. trituberculatus
本研究克隆得到三疣梭子蟹14-3-3cDNA 全长序列,编码由247 个氨基酸组成的蛋白质。14-3-3 基因具有高度的保守性,将本研究克隆得到的三疣梭子蟹14-3-3 基因与其他进行比对,Pt14-3-3 基因与中华绒螯蟹14-3-3 基因同源性最高,达到100%;聚类分析显示,三疣梭子蟹14-3-3 基因氨基酸序列与中华绒螯蟹紧密聚为一支,之后依次与脊尾白虾、日本对虾、拟穴青蟹、斑节对虾、墨吉明对虾聚在一起,同源性均在95%以上。研究发现,14-3-3 基因在哺乳动物中存在β、γ、ε、η、σ、ζ 和θ 7 种亚型(Hermeking, 2003)。根据NJ 系统进化树和氨基酸比对结果,对Pt14-3-3基因进行分析,推测该基因属于ζ 亚型,三疣梭子蟹是否存在14-3-3 基因的其他亚型有待进一步研究。
本研究采用荧光定量RT-PCR 方法对三疣梭子蟹14-3-3 基因mRNA 组织表达进行分析。显示14-3-3基因在多种组织中都有表达,暗示该基因功能的多样性。在肝胰腺中的表达量最高,推测肝胰腺为三疣梭子蟹14-3-3 基因发挥作用的重要器官,而肝胰腺是机体参与免疫防御的重要组织,暗示了该基因具有免疫功能。该结果与拟穴青蟹14-3-3 基因表达的研究结果基本一致(舒妙安等, 2012)。血细胞是三疣梭子蟹重要的免疫细胞,Pt14-3-3 基因在血细胞中有少量表达,同样说明其具有免疫功能(Kaimin et al, 2017)。例如,Chongsatja(2010)等研究发现,凡纳滨对虾病原感染后14-3-3 基因在血细胞中大量表达,证明了该基因的免疫功能。另外,在鳃中也有较多表达,鳃组织与渗透压调节密切相关(Lü et al, 2013; Genovese et al,2004),暗示其可能的在盐度适应中发挥一定的功能。例如,Kaeodee 等(2011)发现,斑节对虾从高渗到低渗的适应中14-3-3 基因大量表达,证明该基因参与机体渗透压的调节。
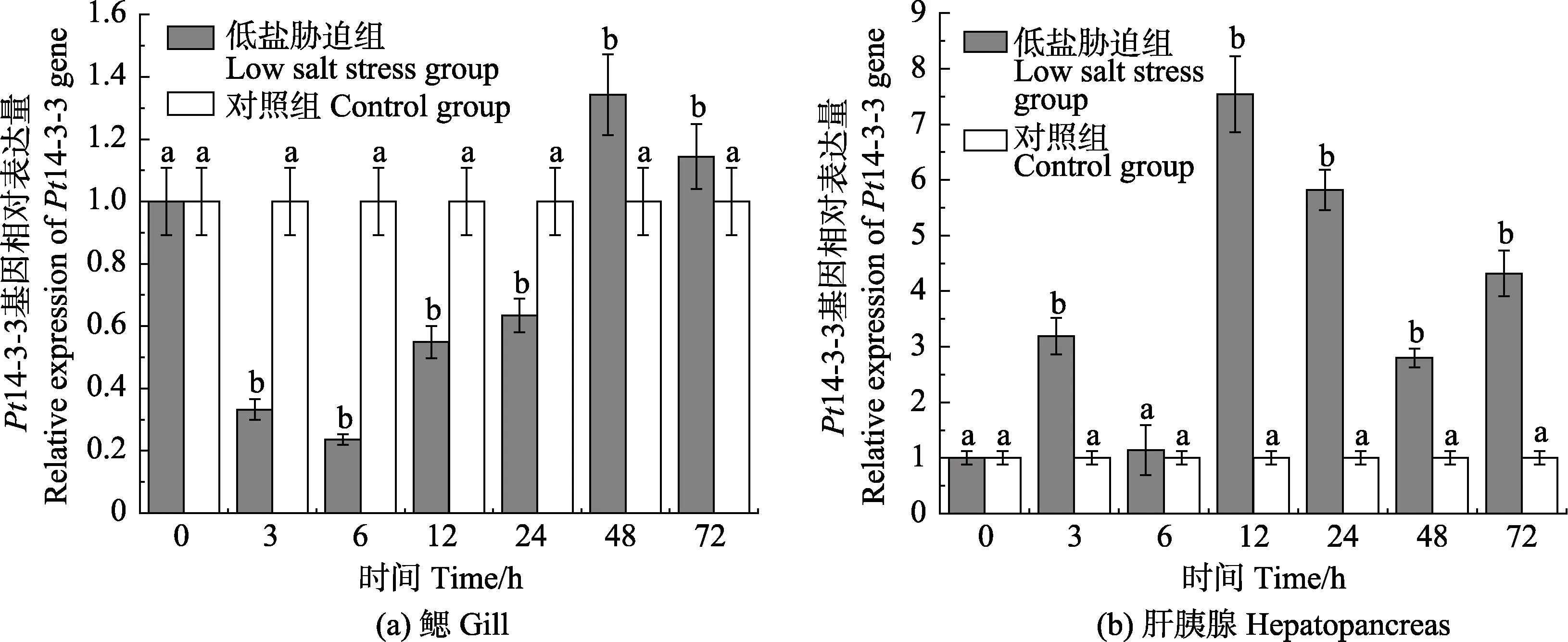
图5 Pt14-3-3 基因在低盐胁迫下鳃和肝胰腺中的表达情况Fig.5 Expression of Pt14-3-3 gene in gill and hepatopancreas under low salt stress
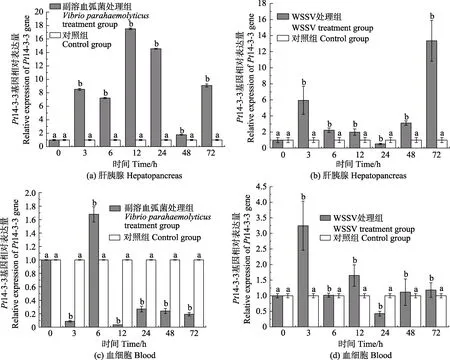
图6 Pt14-3-3 基因在病原胁迫下肝胰腺以及血细胞中的表达Fig.6 Expression of Pt14-3-3 gene in hepatopancreas and blood cells under pathogen stress
为了探究Pt14-3-3 基因在三疣梭子蟹盐度适应中是否具有一定功能,本研究分析了该基因在低盐胁迫下鳃和肝胰腺中的表达规律。发现低盐胁迫后,Pt14-3-3 基因在鳃组织中0~24 h 呈显著下调表达,推测原因可能是低盐胁迫导致鳃组织产生应激损伤,最终间接干扰或抑制了Pt14-3-3 基因的表达;48 h 后基因表达上调,暗示了三疣梭子蟹通过显著上调鳃组织中Pt14-3-3 基因的表达量来应对低盐胁迫。Kammerer等(2009)研究认为,盐度胁迫下罗非鱼鳃上皮细胞凋亡数量随时间的延长而增加。Pt14-3-3 基因属于ζ 亚型,具有凋亡抑制的功能(陈惠华等, 2005)。以上研究表明,三疣梭子蟹可能通过上调Pt14-3-3 基因的表达来抑制鳃组织中细胞凋亡的过程。低盐胁迫后,Pt14-3-3 基因在肝胰腺中于3 h 后呈上调表达,暗示三疣梭子蟹通过上调肝胰腺中Pt14-3-3 基因的表达量来抑制肝胰腺中的细胞凋亡进程,来参与环境的适应性调节。
注射副溶血弧菌和 WSSV 病毒后,肝胰腺中Pt14-3-3 基因的表达量整体上调,基本呈先上升后下降再上升的表达趋势,上升趋势表明Pt14-3-3 基因可能参与了三疣梭子蟹的免疫应答反应,通过增加Pt14-3-3 基因的表达协助清除受损细胞,来维持其正常生理功能(Chang et al, 2008)。下降趋势表明,机体可能处于感染后的恢复期,而再次的上升趋势则可能是三疣梭子蟹恢复期后继续清除残留的副溶血弧菌和WSSV 病毒的表现。2 种致病病原导致的Pt14-3-3基因在肝胰腺中的表达到达最大值的时间不同,可能是由于三疣梭子蟹对于2 种病原的敏感程度和耐受程度不同。感染副溶血弧菌后,三疣梭子蟹14-3-3基因在血细胞中的表达量呈先下降再上升再下降的趋势,该趋势与脊尾白虾感染WSSV 病毒后14-3-3基因(王有昆等, 2016)在血细胞中的变化趋势及凡纳滨对虾感染桃拉病毒后14-3-3 蛋白(Chongsatja et al,2010)的变化趋势相一致,表明三疣梭子蟹可能通过上调14-3-3 基因的表达来抑制肝胰腺和血细胞中的细胞凋亡进程,来参与三疣梭子蟹的免疫应答反应。
本研究成功克隆了三疣梭子蟹14-3-3 基因cDNA全长,初步探究了该基因在三疣梭子蟹低盐适应中鳃和肝胰腺中的表达模式以及感染副溶血弧菌和WSSV 病毒后在肝胰腺和血细胞中的表达模式,证明Pt14-3-3 基因在三疣梭子蟹低盐适应和免疫应答反应中发挥了作用,为进一步研究三疣梭子蟹14-3-3基因在低盐适应中的分子机制及其免疫机制研究和病害防治提供了理论依据。
猜你喜欢
河北渔业(2022年10期)2022-10-15
传染病信息(2022年2期)2022-07-15
中国药学药品知识仓库(2022年9期)2022-05-23
锦州医科大学报(2022年2期)2022-05-07
文萃报·周二版(2022年10期)2022-03-19
渔业科学进展(2021年3期)2021-05-12
现代临床医学(2021年2期)2021-03-29
海峡姐妹(2018年11期)2018-12-19
北京广播电视报(2018年39期)2018-11-23
现代营销·经营版(2017年3期)2017-05-17
