
基于水声学探测的洪潮江水库鱼类资源空间分布及其与环境因子的相关性*
2021-03-19 06:13李育森韩耀全匡天旭吴伟军雷建军何安尤
渔业科学进展 2021年1期
周 磊 李育森 施 军 韩耀全匡天旭 吴伟军 雷建军 何安尤①
(1. 广西壮族自治区水产科学研究院 广西水产遗传育种与健康养殖重点实验室 南宁 530021;2. 华南农业大学海洋学院 广州 510642)
水库是我国重要的淡水资源库,也是我国内陆重要渔业基地和淡水生物种质资源库。因其拥有资源丰富、鱼类物种繁多、品质优良等优势,在渔业生产和供应中起到重要的作用(徐跑等, 2017)。广西地处祖国南疆,属亚热带季风气候区,气候温暖,降水充沛,水域广阔,渔业资源丰富,为淡水渔业养殖提供了得天独厚的条件(周解等, 2006)。据广西水利发展统计公报记载,2015 年广西全区已建成各类水库4545 座,其中,大型水库57 座,中型水库229 座,水库总库容658.23 亿m3,正常水位条件下,水面面积为24.7 万hm2,适宜水产养殖面积为10.67 多万hm2。
与我国大水面渔业的发展一致,广西水库渔业的发展大致经历了天然捕捞—资源增养殖—网围、网拦、网箱养殖的历史轨迹,水库渔产量得到了大幅度的提高,为提升淡水鱼产业和解决库区移民生产、生活问题做出了贡献。很多水库因此成为当地重要的淡水鱼生产基地(刘其根等, 2003)。然而,近10 年来,随着库区水质富营养化进程的加速,传统的网箱、网围投饵养殖模式作为水域污染源之一,已经与库区周边的工业污染源、城镇排污口等点源污染一起被列为取缔的对象。但在许多库区及周边点源污染得到充分治理的情况下,仍无法解决富营养化问题,究其原因是由于流域非点源污染、上游自来水携带的营养盐及库区水体前期累积的营养盐,无法在库区生态系统中转化为生产力,导致累积性富营养化(张益峰等, 2014)。为了控制湖泊、水库水质以及减少生态系统能量的损失,挖掘水体生产潜力,鲢(Hypophthalmichthys molitrix)、鳙(Hypophthalmichthys nobilis)被广泛地投放于各类水体中。然而,由于传统方法的限制,对渔业资源评估只是根据渔民渔获物的数量进行粗略的估计,如何科学快捷地评估渔业资源分布特征及其与环境变化的关系,为增殖放流提供科学支撑是大水面渔业保护与发展的基础。
鱼类资源量调查的常用方法有网捕、钓鱼和水声学探测等(Elliott et al, 2001; Coll et al, 2007; Jurvelius et al, 2016; 张俊等, 2014; 刘鸿等, 2019)。通过钓鱼调查鱼类资源量时,受外界的干扰较大,其本身也难以提供规律和严谨的数据记录。一些网捕的方法虽然可计算被调查水域的鱼类丰度和鉴别出鱼类种类,但这类数据只能代表网捕的一小片区域,若扩大样本量,则不可避免地对鱼体甚至生态群落造成伤害。与上述传统的鱼类资源评估方法相比,水声学探测的方法可轻松地获得连续、长期和丰富的渔业资源数据,且成本相对较低,不会对鱼体造成伤害和给水域带来污染。这种方法更加适用对开阔水域进行长期、大规模的调查(Mowbray, 2002; Mason et al, 2005; Close et al, 2006; 张吉昌等, 2012)。目前,水声学方法已广泛应用于鱼类资源量评估、行为学分析、时空分布动态研究以及底质结构、水深植被和浮游生物等方面的研究。
本研究采用水声学探测技术结合渔获物调查,对广西洪潮江水库鱼类资源的昼夜差异、空间分布进行了探测,评估环境因子对鱼类分布的影响,以期为洪潮江水库渔业资源科学管理提供科学参考。
1 材料与方法
1.1 研究地点
洪潮江源于广西灵山县鹤龟山,洪潮江干流全长为106 km,流域面积为458 km2。洪潮江水库又名星岛湖,位于洪潮江下游,距合浦县城23 km (图1)。该水库是以灌溉为主,结合防洪、发电、旅游、渔业等综合利用的水利工程。水库水面面积为6666.67 hm2,集雨面积为400 km2,多年平均入库水量为3.17 亿m3,总库容7.16 亿m3,有效库容为3.0 亿m3,死库容为2.54 亿m3,为多年调节水库。为了研究方便,将全库划分为9 个区域(图1)。

图1 洪潮江水库及采样点划分示意Fig.1 Location of Hongchaojiang Reservoir and sampling sites
1.2 水声学探测
本研究采用的声学设备为 SIMRAD ES200-7C的 EY60 科研Ⅰ型分裂波束鱼探仪(挪威),换能器为圆形,频率为200 kHz,–3 dB 波束角为7°,SIMRAD公司的MX-500 GPS 导航仪(挪威)确定渔船位置。运用ER60 软件进行水声学数据及地理坐标数据的采集。联想R61 笔记本电脑存储数据。为消除各航次水环境的差异对声学探测的干扰,获得准确的回波信号,本研究参照标准校正流程,采用直径为13.7 mm的原厂标配钨铜金属球(Conti et al, 2006)于每航次探测前对SIMRAD EY60 设备进行校正。
为避免昼夜探测差异对鱼类资源评估的影响,选择一个风浪较小的库汊区,于2018 年4 月8 日晚和4 月9 日白天进行水声学调查评估洪潮江水库鱼类昼夜分布的差异。全库区空间探测选择在2018 年4 月10~11 日进行。探头通过不锈钢管进行固定,置于调查船右前方水下 0.5 m 处,方向垂直向下,采用“之”字形走航式探测采样,船速约为8~10 km/h。调查期间,仪器的脉冲长度为64 μs,分辨精度为0.012 m,脉冲频率设置为最大值。
1.3 渔获物调查与环境因子监测
水声学探测时,同步对洪潮江水库(图1,S1~S9站)进行理化因子监测,监测指标有水深(m)、水温(℃)、透明度(cm)、浊度、pH、溶解氧(DO, mg/L)、总磷(TP,mg/L)、总硬度(TH, mg/L)、总氮(TN, mg/L)、亚硝态氮(NO2–-N, mg/L)、硝酸盐氮(NO3–-N, mg/L)、氨氮(NH4+-N, mg/L)、高锰酸盐指数(CODMn, mg/L)、叶绿素a (Chl-a, μg/m3)、总有机碳(TOC, mg/L)等。其中,水温、pH 和DO 采用YSI PRO PLUS 现场测量;透明度和浊度分别使用塞氏盘和浊度计进行现场测量;水深通过鱼探仪导出;其他参数带回实验室采用标准方法测量。
为了了解洪潮江水库鱼类组成,采用刺网和地笼网结合,对S2、S3、S6、S7 和S9 站位进行采样。每个采样点放置25 m 地笼网5 张(网框高为33 cm,宽为45 cm,网目为0.7 cm)、60 m 刺网3 张(网高为1.5 m,网目分别为6、8、10 cm)。于19:00~21:00 放置,第2 天07:00~09:00 取出,放置时间约12 h。鱼类根据《广西淡水鱼类志(第二版)》(周解等, 2006)鉴定到种,计数并测量重量。
1.4 数据处理与分析
使用Sonar5 数据后处理软件转换和分析获得的声学数据。转换时,40logR 的噪音阈值设置为–60 dB,通过Sonar5 软件中Bottom detection 自动识别水底,有明显识别错误的地方手动进行矫正,表层线设置为探头以下1.5 m,以排除表层噪音及近场效应的影响。水底线在Bottom detection 和人工矫正的基础上提高0.5 m,以避免水底噪音的干扰。
数据分析的相关声呐参数分别是最小回波长度,为0.6,最大回波长度为1.8;最大相差0.5;声学截面的最大增益补偿(Max Gain Compensation) 设置为6 dB。采用回声计数法计算鱼类的体积密度。
目标强度和鱼体体长之间的关系采用 Foote(1987)提出的公式:

式中,TS 为目标强度(dB),L 为全长(cm)。据此,本研究阈值–60 dB 对应鱼类全长为3.94 cm。
运用方差分析对鱼类昼夜及空间差异进行显著性检验;Pearson 相关性检验用于分析各环境变量和鱼类密度、目标强度之间的相关性。上述分析均采用R 3.5.1 软件完成。
将获取的鱼类密度数据导入Arcgis 软件,采用反距离加权法(Inverse Distance Weighting, IDW) 进行插值运算,以进行空间可视化。
2 结果
2.1 洪潮江水库鱼类组成
网捕共采集鱼类1299 尾,重为19711 g,共计18 种。其中,罗非鱼(Oreochroms mossambcus)无论从数量还是生物量都占有绝对优势,分别占比79.68%和 79.69%。在数量上占比超过 1%的还有须鲫(Carassioides cantonensis)(5.08%)、高体鳑鲏(Rhodeus ocellatus)(4.85%)、海南似(Toxabramis houdemeri)(4.08%)、台细鳊(Rasborinus formosae)(3.54%)、子陵吻虾虎鱼(Rhinogobius giurinus)(1.46%)。在生物量上占比超过1%的种类还有须鲫(12.58%)、海南似(1.79%)、露斯塔野鲮(Labeo rohita)(1.44%)、海南鲌(Culter recurviceps)(1.35%)(表1)。
2.2 水声学探测昼夜差异
在S9 站选择一个风浪较小的库汊,对鱼类的昼夜水声学探测差异进行分析。研究发现,鱼类密度昼夜差异不显著(P>0.05)(图2),昼夜密度平均值分别为(105.02±12.59)(n=36)和(106.71±16.28)ind./1000 m3(n=29)。但目标强度平均值白天>夜间(P<0.05),昼夜目标强度平均值分别为(–43.30±0.32) 和(–44.50±0.26) dB,主要是夜晚声学探测覆盖区域的(–60 ~ –54 dB)小型鱼类增加(–60~ –57 和–57~ –54 dB 分别增加9.25%和4.76%),而白天声学探测覆盖区域的(–54 ~ –48 dB)鱼类比例增加所致(–54 ~ –51 和–51 ~ –48 dB 分别增加7.42%和5.27%)(表2 和图2)。
2.3 鱼类空间分布特征
对洪潮江水库鱼类密度的空间分布格局分析发现,洪潮江水库鱼类分布呈现显著的空间变化特征(P<0.05)。总体趋势上,鱼类密度均呈上游密度高于下游密度的特征。密度平均值为109.52 ind./1000 m3,最大值出现在上游的S3 站,平均密度为330.21 ind./1000 m3。最小值为下游的S8 站,平均密度为21.50 ind./1000 m3(图3 和表3)。

?

图2 昼夜水声学探测的鱼类密度和目标强度(TS)比较Fig.2 Comparison of the density and target intensity (TS) of hydroacoustic detection in day and night
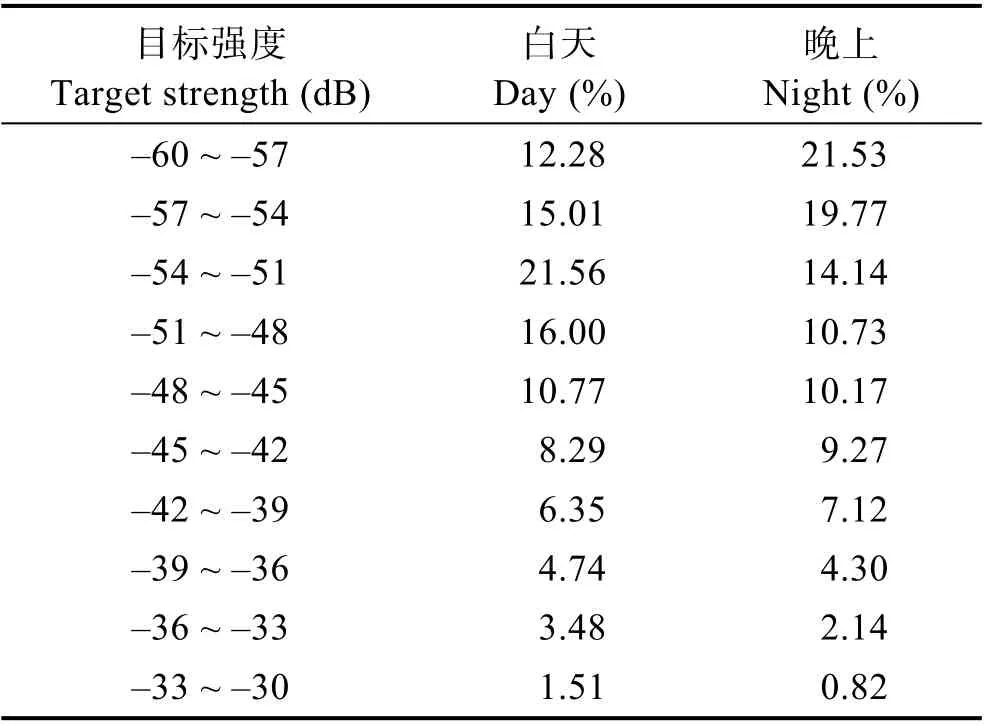
表2 昼夜水声学探测目标强度分布的比较Tab.2 Comparison of target strength distribution by hydroacoustic detection between day and night

图3 鱼类空间分布(IDW 插值)Fig.3 Fish spatial distribution by IDW interpolation

表3 洪潮江水库鱼类密度空间分布Tab.3 Spatial distribution of fish density in Hongchaojiang Reservoir
目标强度则表现出相反的特征,最大值出现在S9 站,平均值为–45.10 dB,对应全长为21.88 cm。最小值为S3 站,平均值为–47.90 dB,对应全长15.87 cm(表4)。相关性分析表明,鱼类密度与目标强度存在一定的负相关,但相关性不显著(R=–0.553, P=0.122)。
2.4 环境因子关联分析
相关性分析表明,鱼类密度分布与TP、CODMn、Chl-a 呈显著正相关,与水深呈显著负相关(P<0.05)。鱼类目标强度与WT、浊度、TP、CODMn 呈显著负相关(图4)。

表4 洪潮江水库鱼类目标强度(TS)空间分布Tab.4 Spatial distribution of target strength (TS)in Hongchaojiang Reservoir
3 讨论
3.1 昼夜差异对水声学评估的影响
与传统采样工具相比,水声学方法由于具备直接采样、采集迅速、空间上采样水体大、时间上可连续采集数据、不损伤鱼体等多重优势(Elliott et al,2001),因而,在鱼类资源和生态学的研究中应用越来越广泛。然而,水声学评估的可靠性会受仪器性能、鱼类行为(如集群行为)、环境因素的影响(Simmonds et al, 2005)。由于水体流速较缓,通常认为鱼类行为节律是影响湖泊、水库鱼类资源水声学评估结果准确性的首要考虑因素。
为了获取更准确的水声学调查结果,本研究对洪潮江水库鱼类昼夜差异进行了评估。结果显示,洪潮江水库鱼类昼夜差异不显著,白天和晚上鱼类密度探测结果分别为(105.02±12.59)和(106.71±16.28)ind./1000 m3。但目标强度白天显著大于夜晚,目标强度平均值分别为–43.30 dB 和–44.50 dB,夜晚小型鱼类的比重明显增加。已有大量研究表明,小型鱼类夜晚活动增加与其逃避敌害有关。水声学调查不可能覆盖全部水体,鱼类的集群行为会降低鱼类被水声学仪器探测到的几率,从而低估鱼类资源密度。大部分鱼类一般在白天集群,夜晚分散;同时,由于鱼类存在昼夜垂直迁移现象,有些鱼类白天多躲避于水底,夜晚向表层迁移觅食,水底探测盲区的存在造成白天探测结果偏低(Ye et al, 2013; 连玉喜等,2015)。但这种昼夜差异也不是绝对的,Djemali 等(2009)对突尼斯4 个水库的水声学探测表明,在多鱼种共存的水库,虽昼夜差异存在,但无法得出相同的规律。有些水库白天鱼类探测密度较高,而有些水库黑夜探测密度较高。因此,综合考虑结果的可靠性和走航安全性,洪潮江水库后续水声学调查可选择白天进行。

图4 鱼类空间分布与环境因子的关联Fig.4 Correlation between fish spatial distribution and environmental factors
3.2 洪潮江水库鱼类空间分布特征及其对环境因子的响应
鱼类空间分布特征及其与环境因子作用关系的研究有助于了解鱼类的行为规律与资源变动趋势。Dauble 等(1989)通过对大马哈鱼(Oncorhynchus keta)幼鱼在哥伦比亚河空间分布研究分析其早期的迁徙规律。Silvano 等(2000)研究发现,茹鲁阿河(Juruá River)干流鱼类资源的密度明显高于其附属湖泊鱼类资源的密度,并在此基础上提出了相应的保护措施。影响鱼类密度时空差异的主要因素有水深、离岸距离、底质特点、植被状况、浮游生物和捕食者等。正确认识这些因素对鱼类时空分布格局的影响,将为洪潮江水库鱼类资源的评估、保护及科学捕捞提供重要的参考。网捕法结果显示,洪潮江水库鱼类主要由罗非鱼、须鲫、高体鳑鲏、海南似、台细鳊、子陵吻虾虎鱼、露斯塔野鲮、海南鲌等鱼类组成。由于整个库区较小,对全库区5 个站点的抽样调查显示,种类组成的空间差异较小。但水声学探测结果表明,洪潮江水库鱼类密度却表现出了明显的空间分布差异性,鱼类密度呈上游大于下游、库汊大于库心的空间分布格局。谭细畅等(2002、2008)对东湖和三峡库首、陶江平等(2008)对三峡水库成库期间、Zeng 等(2015)对广西百色水库海南似的水声学调查也得出了类似的结论,即水库(湖泊)鱼类的空间分布特征表现出明显的斑块异质性。
环境因子的时空差异是造成鱼类资源时空变动的主要原因,鱼类往往沿着环境梯度有着对应的密度梯度分布特征(Prchalova et al, 2008)。本研究发现,对洪潮江水库鱼类分布影响最大的环境因子为TP、水深、叶绿素浓度、CODMn。其中,TP、叶绿素浓度、CODMn 与鱼类密度呈显著正相关,而水深与鱼类密度呈显著负相关。TP、叶绿素浓度、CODMn 反映的是水体营养物质的多寡或水体初级生产力的大小(蒋万祥等, 2010),营养盐、初级生产力高的水域有利于饵料生物的生长,尤其是叶绿素浓度与鱼类分布关系间接反映了饵料生物与鱼类分布的相关性。Godlewska 等(2003)研究发现,中营养化的Solina 水库鱼类资源量比富营养化水平Dobczyce 和Sulejow 水库资源量少一个数量级,表明一定程度的富营养化有利于水库容纳更多的鱼类。许多研究报道了水深和鱼类密度的相关性(Djemali et al, 2009; 陶江平等, 2008; Wang et al,2013)。幼鱼往往偏好栖息于浅水区,浅水区饵料食物更丰富,并且大型捕食者也较少(Blaber et al,1980)。Linehan 等(2001)研究表明,增加水深会增加鳕鱼(Gadus)幼鱼被肉食性鱼类捕食的风险。洪潮江水库浅水区不仅可以为鱼类提供必要的栖息地,也会使其免受掠食者的捕食,因而,鱼类密度往往高于深水区。
此外,本研究发现,鱼类的平均目标强度大小与水温、浊度、TP、CODMn 呈显著负相关,TP、CODMn间接反映了饵料的丰富度,因此,越丰富的区域往往幼鱼的比例更高,目标强度就越小。水温也是影响鱼类生长的主要因子,水温能改变代谢的速度。在鱼类生理限制的范围内,适当提高水温有利于鱼类的存活和快速生长,因此,鱼类尤其是幼鱼会趋向水温高的区域(Ribeiro et al, 2006; 黄良敏等, 2011)。浊度影响主要体现在索饵与避敌 2 个方面(Cyrus et al,1992)。鱼类利用较高的浊度来避免被捕食,增加鱼群的生存能力(Whitfield et al, 2012)。一般情况下,浊度较高的水域其底质的平均粒径相对较小,饵料生物的丰度相对较高,有利于鱼类的索饵(Islam et al,2006)。另一方面,浊度较高的水域环境有利于降低非肉食性被捕食的概率,提高种群存活率(Cyrus et al,1992),即增大了肉食性鱼类的捕食难度,因此,在高浊度水域中,小型鱼类或者仔、稚鱼丰度往往较高(North et al, 2003)。综上所述,洪潮江水库鱼类空间分布是多种非生物因子(如叶绿素a、TP 等)共同作用的结果,其在一定程度上也反映了生物因子(鱼类索饵、逃避敌害等)对鱼类分布的影响。由于本调查仅2018 年4 月1 个航次,还有待进行更长时间的监测,以掌握资源量的季节动态、年际变动趋势,以便提出更有效的渔业资源管理和保护措施。
猜你喜欢
中国三峡(2022年6期)2022-11-30
山西化工(2022年6期)2022-10-09
绿色科技(2022年8期)2022-05-25
建材发展导向(2021年10期)2021-07-16
药学与临床研究(2021年3期)2021-07-13
儿童时代·幸福宝宝(2020年9期)2020-09-08
知识就是力量(2019年9期)2019-09-09
绿色科技(2018年20期)2018-12-19
探索科学(2017年5期)2017-06-21
都市家教·上半月(2016年4期)2016-11-19
