
胆管细胞型肝癌lncRNA-mRNA 共表达网络的高通量基因芯片筛查分析
2021-03-08 03:25唐津天王瑞宾王伯庆
中国医药导报 2021年3期
唐津天 王瑞宾 王伯庆▲
1.新疆医科大学附属肿瘤医院肝胆胰外科,新疆乌鲁木齐 830000;2.新疆医科大学第三临床医学院,新疆乌鲁木齐 830000
胆管细胞型肝癌是来源于肝内胆道上皮细胞的恶性肿瘤,发病率较低,没有特异性的肿瘤标志物用于血清学诊断,糖类抗原(CA)19-9 和胚胎抗原(CEA)的增高可能有助于诊断[1]。病理表现为腺癌,肝外受侵表现为淋巴结转移和周围神经侵犯[2]。大多数胆管细胞型肝癌发病呈散发性,没有特征性的风险因素[3]。目前,胆管细胞型肝癌的发病机制尚不明确。
人类基因组研究发现93%的DNA 被转录为RNA,其中2%可翻译成蛋白转录产物,其余98%均为无编码功能的非编码RNA(non-coding RNA,ncRNA)[4-5]。长链非编码RNA 是非编码RNA 的一种,在表观遗传调控、转录调控以及转录后调控等多层面调控基因的表达。研究证实lncRNA 与人类多种重大疾病的发生发展密切相关[6-10]。本研究借助于高通量生物芯片进行筛选分析,构建共表达网络,获取与肿瘤相关lncRNAmRNA,为胆管细胞型肝癌的诊断和治疗提供依据。
1 资料与方法
1.1 标本收集
选取2017 年1 月—6 月新疆医科大学附属肿瘤医院(以下简称“我院”)肝胆外科手术切除的5 例胆管细胞型肝癌新鲜组织标本。标本自手术中离体后立即取样,注明标识后置入液氮中保存。
5 例经病理证实均为胆管细胞型肝癌患者,女3 例,男2 例;年龄37~55 岁,中位年龄45.2 岁;肿瘤均单发,最大径介于2.2~4.7 cm,平均(4.2±0.3)cm,病理分期均为T1N0M0(AJCC,第8 版)。本研究经医院医学伦理委员会的批准,并获得患者及其家属的同意。
1.2 RNA 的提取和数据处理
RNA 提取试剂盒提取样本总RNA,通过琼脂糖凝胶电泳检测RNA 完整性。总RNA 质检合格后,样本依次进行标记、芯片杂交、洗脱和染色,最后扫描获得原始数据图像。样本数据分析由北京康普森生物技术有限公司支持完成。
1.3 差异mRNA 的筛选
使用Affymetrix 软件对获取的数据进行分析。根据筛选条件log2|FC|≥1 且P ≤0.05,对得到的标准化信号值的FC 值和差异显著性P 值进行筛选。
1.4 lncRNA-mRNA 共表达网络的构建
Affymetrix 软件进行统计学分析后获取差异lncRNA。对Top10 差异lncRNA 进行共表达分析,获得相应的靶基因。依据Gene Ontology 数据库和KEGG 数据库进行分析,预测lncRNA 的功能。再次选取Top10 差异lncRNA 中|FC|值最高的Top1 lncRNA进行GO 富集和Pathway 分析。
1.5 qRT-PCR 验证分析
另选取我院肿瘤标本库保存的胆管细胞型肝癌的组织标本30 例。随机抽取Top10 差异mRNA 中的1 个差异mRNA 进行qRT-PCR 验证;对Top1 lncRNA及其共表达mRNA 进行qRT-PCR 验证。根据Real-Time PCR 原始检测数据,按照F=2-△△Ct计算RNA 的相对表达量。组织RNA 的提取使用天根生化的动物组织总RNA 提取试剂盒(生产批号:DP451)。使用Takara 的PrimeScriptTMRT reagent Kit with gDNA Eraser反转录试剂盒(生产批号:RR047A)进行反转录获取cDNA,利用Takara 的SYBR Premix Ex Taq Ⅱ(Tli RNaseH Plus)(生产批号:RR420)进行Real-Time PCR反应。从NCBI 数据库和PrimerBank 数据库查找引物序列,委托生物公司设计合成。
1.6 统计学方法
使用SPSS 16.0 统计软件进行数据分析。正态分布计量资料采用均数±标准差(±s)表示,比较采用t检验;偏态分布计量资料采用中位数表示。多组间比较选择单因素方差分析。以P <0.05 为差异有统计学意义。
2 结果
2.1 差异mRNA 和lncRNA 的筛选
在筛选阈值为P ≤0.05,log2|FC|≥1 的情况下,共筛选获得差异mRNA 475 个,其中上调mRNA 213 个,下调mRNA 262 个;共筛选获得差异lncRNA 共计438 个,其中上调lncRNA 131 个,下调lncRNA 307 个。根据差异表达的mRNA 和lncRNA 进行聚类分析,分析结果见图1 和图2(封四)。根据log2|FC|值,其中差异表达的Top10 mRNA 和lncRNA 见表1~2。
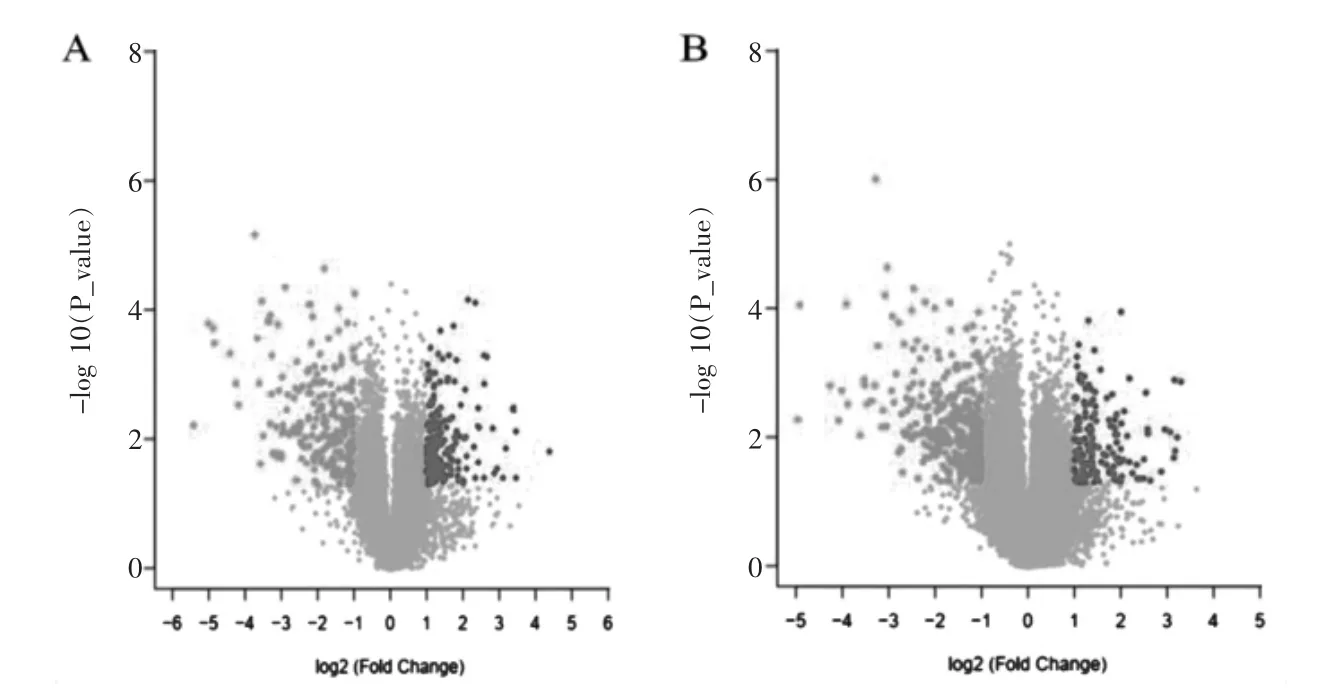
图1 差异表达mRNA 和lncRNA 的火山图分布
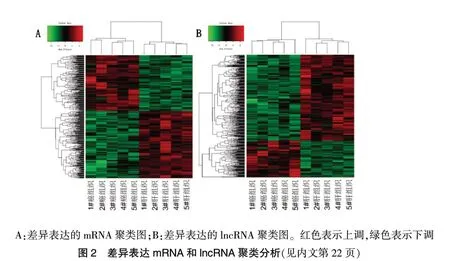
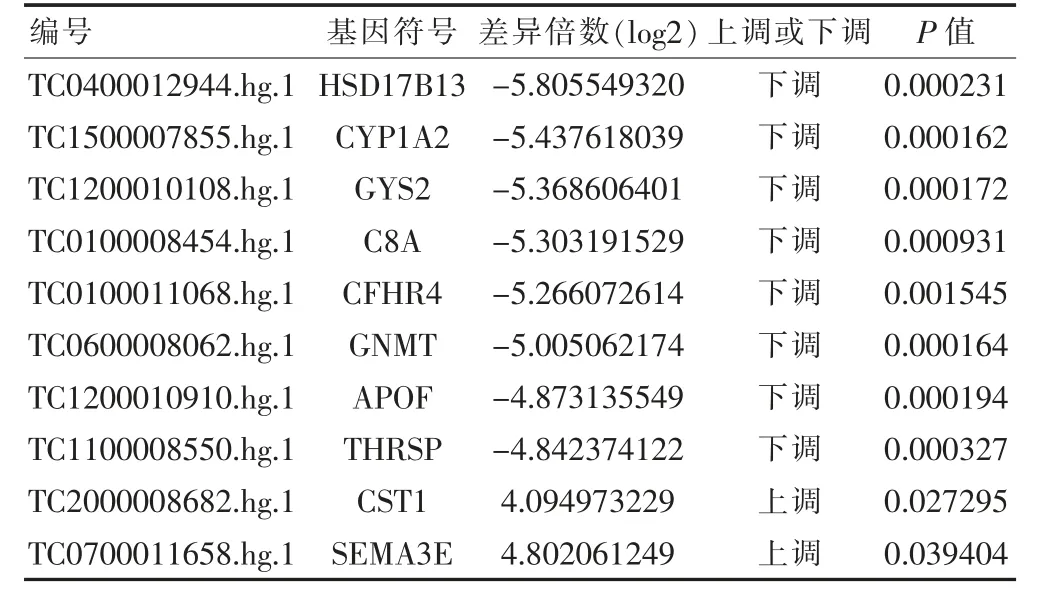
表1 排名前10 的差异表达的mRNA
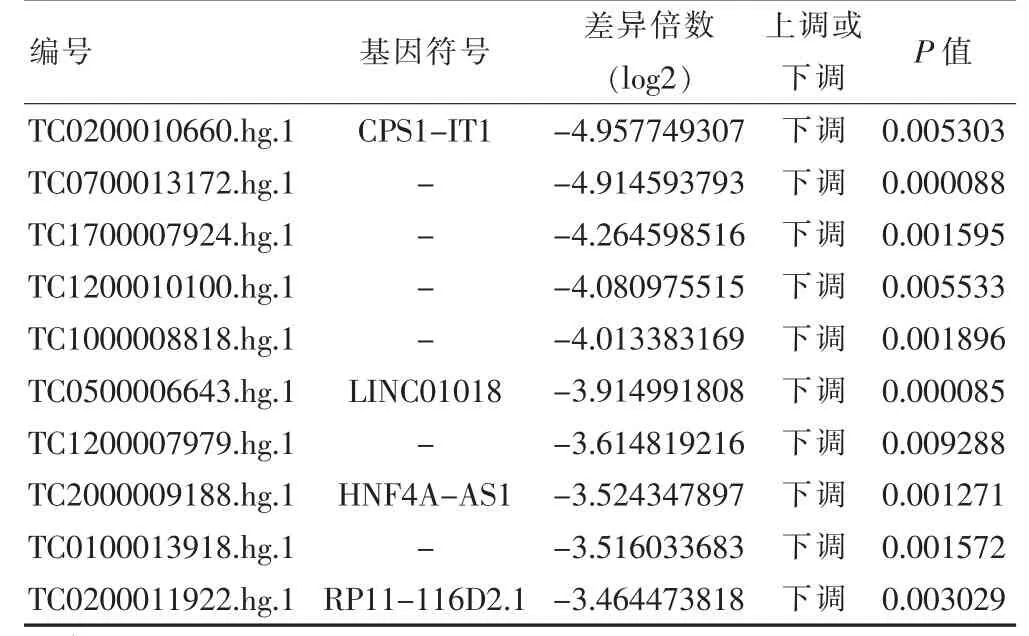
表2 排名前10 的差异表达的lncRNA
2.2 mRNA-lncRNA 共表达分析
对Top10 差异lncRNA 进行lncRNA 与mRNA的共表达分析,获得相应的靶基因。在筛选阈值为P ≤0.05,log2|FC|≥1,相关系数|cor|≥0.95 的情况下,Top10 差异lncRNA 共表达的mRNA 数目见表3。对Top10 lncRNA 相应靶基因的GO 富集分析和Pathway分析,因获得的数据信息量巨大,仅选择Top1 lncRNA进行GO 富集分析和Pathway 分析。

表3 排名前10 的lncRNA 共表达mRNA 数目
2.3 CPS1-IT1 的共表达分析
选择|FC|最大值的lncRNA CPS1-IT1 进行共表达分析。通过GO 分析最终筛选获得与之关系密切的共表达基因是CSNK1E,同时对CPS1-IT1 的功能进行分类(图3)。通过Pathway 分析,获得CPS1-IT1 相关的信号通路(图4)。
2.4 LncRNA CPS1-IT1 的验证
实时荧光定量PCR 检测30 例胆管细胞型肝癌的癌组织和癌旁正常肝组织中靶基因mRNA 和lncRNA 的表达丰度。随机抽取Top10 mRNA 中的TC1200010108.hg.1(GYS2)和CPS1-IT1 的共表达基因CSNK1E 进行验证分析。结果显示TC1200010108.hg.1(GYS2)在癌组织中呈相对表达下调,癌组织2-△△Ct=0.5413,癌旁正常肝组织2-△△Ct=1,P <0.05;CSNK1E在癌组织中呈表达上调,癌组织2-△△Ct=1.8907,癌旁正常肝组织2-△△Ct=1,P <0.05(图5)。同时对Top1 lncRNA 进行验证,结果显示CPS1-IT1 在癌组织中呈相对表达下调,癌组织2△△Ct=0.2973,癌旁正常肝组织2-△△Ct=1,P <0.05(图5)。
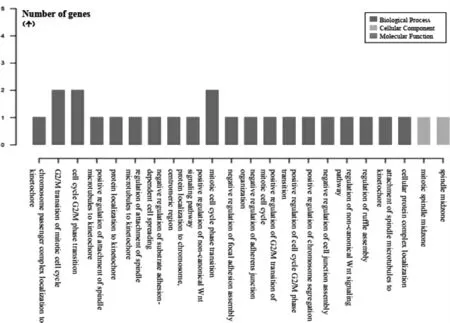
图3 lncRNA CPS1-IT1 的功能分类

图4 lncRNA CPS1-IT1 共表达基因Pathway 分类图
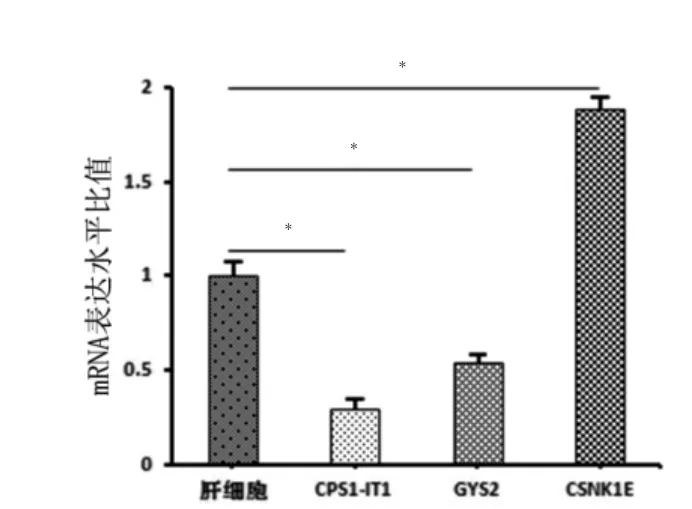
图5 CPS1-IT1、GYS、CSNK1E 在组织RNA 中的表达水平
3 讨论
本研究通过生物芯片对胆管细胞型肝癌标本组织进行高通量分析,结果显示在筛选阈值为P ≤0.05,log2|FC|≥1 的情况下,下调表达的差异mRNA 多于上调表达的差异mRNA。同样的情况在lncRNA 中更为明显,在共计438 个差异lncRNA 中,下调表达的差异lncRNA 为307 个,上调表达的差异lncRNA 仅为131 个,这种表达趋势在Top10 差异mRNA 和lncRNA中更加明显。对Top10 差异mRNA 进行分析发现,大多数mRNA 涉及糖代谢和脂代谢。例如,HSD17B13为蛋白编码基因,与脂质代谢相关[11]。GYS2 为肝糖原合成酶,是糖代谢信号通路中的关键蛋白之一,在胰岛素信号通路和糖原代谢信号通路中发挥重要作用。GYS2 表达下调导致下游肝糖原合成不足,加速细胞对葡萄糖的摄取,符合恶性肿瘤对葡萄糖高代谢的特征[12]。CFHR4 则与脂质的运输密切相关。APOF 参与脂质转运,与脂蛋白代谢密切相关[13]。THRSP 在脂质合成中发挥调节作用,特别是对中长链脂肪酸的三酰甘油合成具有重要意义[14]。由此可见,在肝胆管细胞型肝癌中,肿瘤组织的糖代谢和脂质代谢紊乱相对明显。
因高通量分析后获得的数据信息量巨大,因此本研究依据差异倍数,选择Top1 lncRNA 进一步分析。CPS1-IT1 位于编码氨甲酰磷酸合成酶1 基因的内含子,是近年来确定的新的肿瘤相关分子,主要功能角色尚不清楚,目前已经在部分肿瘤中进行了探索性研究。研究发现,CPS1-IT1 在肺癌中具有抑制肿瘤的作用,其过表达可以抑制细胞增殖、迁移和侵袭[15]。此外,CPS1-IT1 在肝细胞癌组织中的表达显著降低,通过调节HIF-1α 活性抑制上皮间质转化抑制肝癌转移[16]。进一步研究表明,Foxa2 表达增加促进CPS1-IT1上调,降低HIF-1α 的活性,抑制上皮-间质转化[17]。在卵巢癌的研究中也证实CPS1-IT1 是一种保护性的预后因子[18]。
对CPS1-IT1 进行共表达分析,最终筛选获得CPS1-IT1 的共表达基因为CSNK1E。通过CPS1-IT1的功能分析发现,CPS1-IT1 及其共表达mRNA 网络可能参与有丝分裂细胞周期转换、Wnt 信号通路的调控、昼夜节律基因表达的调节等。此外,通过Pathway分析发现,CPS1-IT1 和其共表达基因CSNK1E,通过FOXO 信号通路参与糖酵解、葡萄糖异生等;通过Wnt 信号通路参与细胞周期的调控;通过Hedgehog 信号通路参与细胞增殖;通过Hippo 信号通路参与蛋白质水解调控;通过昼夜节律通路参与生物钟基因的调控。
酪蛋白激酶1E(casein kinase 1 epsilon,CSNK1E)可以作用于底物酪蛋白使其磷酸化,主要参与Wnt信号通路的转导,也是生理性昼夜时钟调控的核心成员之一[19-20]。生物钟基因突变常与代谢缺陷有关,包括糖酵解途径、脂肪酸合成、脂肪酸氧化和细胞解毒等,特别是在脂质和葡萄糖代谢方面[20-25]。这些数据表明,昼夜节律和代谢途径的相互协调对于细胞内稳态和机体健康是至关重要的。
本研究显示,胆管细胞型肝癌的糖代谢和脂质代谢紊乱相对明显,其机制之一可能是通过lncRNA CPS1-IT1 和共表达基因CSNK1E 介导下调控肿瘤的葡萄糖和脂质代谢,具体的信号通路仍需进一步验证。
猜你喜欢
昆明医科大学学报(2020年12期)2021-01-26
中成药(2018年9期)2018-10-09
生物医学工程学进展(2018年1期)2018-03-26
中成药(2018年1期)2018-02-02
山东医药(2017年20期)2017-07-01
中成药(2017年4期)2017-05-17
中国学术期刊文摘(2016年1期)2016-02-13
山东医药(2015年44期)2015-02-28
中成药(2014年9期)2014-02-28
中国中西医结合外科杂志(2013年3期)2013-03-11
