
不同施氮水平对柳枝稷光合特性及抗旱性的影响
2021-01-27 03:25何海锋闫承宏吴娜刘吉利贾瑜琀
草业学报 2021年1期
何海锋,闫承宏,吴娜,刘吉利,贾瑜琀
(1.宁夏大学农学院,宁夏 银川750021;2.宁夏大学资源环境学院,宁夏 银川750021;3.宁夏旱区资源评价与环境调控重点实验室,宁夏 银川750021)
柳枝稷(Panicum virgatum)是起源于北美洛基山脉以东、北纬55°以南大草原的多年生草本C4作物[1−3],因其适应性广、耐贫瘠、产量高、多年生和易于管理等特性被视为理想的生物能源作物[3]。柳枝稷木质素和纤维素含量极其丰富,最主要的乙醇转化来源是纤维素和半纤维素,同时水分和灰分含量少,具有较高的产量潜力和较强的耐旱耐瘠薄能力[3−5],相比其他多年生草本作物和传统农作物,柳枝稷能够适应粘壤土、砂壤土等多种土壤类型,具有较低的投入和维护成本、较高的净能源产出,一旦建植成功可持续利用至少15 年以上[6−7],是良好的生态修复和生物改良材料[8]。氮素营养是植物生长发育必不可少的大量营养元素之一,也是植物体内蛋白质、核酸、叶绿素等的重要组成部分,对植物的叶绿素荧光特性及产量和品质具有明显的调节作用,研究合理施用氮肥,是改善植物光合特性,促进植物生长的一项重要调控措施,对提高氮肥利用率和作物的产量、品质,保护生态环境均具有重要意义[8−9]。
叶片是作物进行光合作用的主要器官,其叶绿素含量和光合速率的高低可以反映出作物光合潜力和实际光合产量水平[9]。光合作用是作物生长发育和产量形成的生理基础,作物产量90%~95% 来自光合产物[10−11]。研究表明,施氮可减轻干旱胁迫对作物生长发育与光合作用的不良影响,改善作物叶片的气体交换状况、提高对光能的吸收利用,同时提高干旱环境下叶片光合特性的适应能力;提高作物渗透调节物质含量,从而降低活性氧对细胞结构和功能的伤害。通过施氮可在一定程度上提高作物抗旱能力,但是随着施氮量的増加,作物抗旱性呈现出下降的趋势,过量施氮可能会降低作物的抗旱性[11]。范希峰等[12]研究发现,盐碱胁迫下柳枝稷光合作用受到抑制,但依然可以存活,属于典型的耐盐碱作物。氮素是作物必需的大量营养元素之一,是作物生长的重要物质基础,对作物的器官建造、物质代谢、生化过程、作物产量及品质的形成等都有不可替代的作用,被称为“生命元素”[13]。黄瑾等[14]研究发现,适当的增施氮肥能有效改善柳枝稷光合特性,当土壤中的氮素不能满足作物需求时,要靠施肥来补充和调节,但是土壤中氮素含量过多,会对环境造成污染。宁夏银北地区是我国典型的盐碱地之一,凭经验施肥,偏施氮肥,严重影响了作物的品质和产量,同时容易造成环境污染[15−16],目前关于柳枝稷的研究大多集中在引种适应性、生物质产量等方面,对氮肥调控其光合特性及抗旱性的研究鲜有报道。因此,本研究旨在深入分析盐碱地柳枝稷氮肥施用效果,探讨其对柳枝稷叶片光合作用及抗旱性的影响,以期筛选出既能保证作物高产又能不污染环境的理想施氮水平,为盐碱地生物改良、生态修复及合理施肥提供科学依据。
1 材料与方法
1.1 试验地概况
试验在宁夏大学西大滩盐碱地综合改良试验站进行,该试验站位于宁夏石嘴山市平罗县前进农场(海拔1150 m,38°50′N,106°24′E)。试验地块选择中度盐碱地,研究区域属于典型的半干旱大陆性气候,年均气温在−20.5~33.8 ℃,平均气温为9.5 ℃,≥10 ℃的积温为3350 ℃,年降水量为205 mm,年均蒸发量1875 mm。试验地块为典型的龟裂碱土,耕层土壤(0~20 cm)的有机质及全盐含量分别为8.47 g·kg−1和3.78 g·kg−1,碱解氮、速效磷和速效钾含量分别为20.5 mg·kg−1、5.8 mg·kg−1和64.3 mg·kg−1,土壤碱化度为18.5%,pH 值为8.93。
1.2 试验设计
本试验选取的柳枝稷材料为Cave-in-Rock品种,其生态类型为高地型,染色体倍性为八倍体,原产地为Southern Illinois 38º N,种子由北京草业与环境发展研究中心提供。柳枝稷采用育苗移栽方式种植,于2016 年3月底在温室内育苗,待幼苗长到5 叶期,挑选出长势一致、生长健壮的植株幼苗进行移栽。建植当年每hm2施60 kg 纯N、50 kg P2O5和50 kg K2O,建植第二年每hm2施120 kg 纯N,从第3 年起,开始实施不同施氮量处理,按照当地旱作物进行灌溉管理。采用大田试验,进行随机区组设计,设置4 个施氮水平,无氮添加(无人工施肥,N0)、纯N 60(低氮,N60)、纯N 120(中氮,N120)和纯N 240 kg·hm−2(高氮,N240),供试肥料为尿素,每个水平设3 次重复,分小区种植,共计12 个小区。每个小区长8 m,宽5 m,小区面积为40 m2,种植行距为50 cm,株距为20 cm。
1.3 测定指标及方法
1.3.1 光合作用及叶绿素含量的测定 在柳枝稷每个生育时期内,早上9:00−11:00 及下午14:00−16:00自然光照下,采用美国的LI-6400 便携式光合仪测定柳枝稷倒二叶(从上数第2 片全展叶)的净光合速率(net photosynthetic rate,Pn,μmol·m−2·s−1)、气孔导度(stomatal conductance,Gs,mol·m−2·s−1)、细胞间隙CO2浓度(intercellular carbon dioxide concentration,Ci,μmol·mol−1)、蒸腾速率(transpiration rate,Tr,mmol·m−2·s−1)。采用标准叶室,开放式气路,流速均设定为500 μmol·s−1,当CO2摩尔浓度为400 μmol·mol−1,温度为25 ℃时进行气体交换参数的测定,每个小区获取5 次重复读数,取其平均值。根据以上测定结果计算叶片水分利用效率(water use efficiency)WUE=Pn/Tr;采用Minolta SPAD-502 型便携式叶绿素计(日本)测定柳枝稷叶片叶绿素相对含量(relative chlorophyll content,SPAD),每个小区测5 次重复,然后取其平均值。
1.3.2 主要渗透调节物质和抗氧化酶活性的测定 超氧化物歧化酶(superoxide dismutase,SOD)活性采用氯化硝基四氮唑蓝(NBT)光化还原法测定;过氧化物酶(peroxidase,POD)活性采用愈创木酚法测定;过氧化氢酶(catalase,CAT)活性采用紫外吸收法测定;游离脯氨酸(free proline,Pro)含量采用茚三酮比色法测定;丙二醛(malondialdehyde,MDA)含量采用硫代巴比妥酸法测定;可溶性糖(soluble sugar,SS)含量采用蒽酮乙酸乙酯比色法测定;相对电导率采用电导仪直接测定。具体测定方法见高俊凤[17]主编的作物生理学实验指导书。
1.4 隶属函数法进行抗旱性综合评价
参考张朝阳等[18]和石永红等[19]的方法,采用模糊数学隶属函数法,利用各抗旱指标对各柳枝稷材料进行隶属函数平均值的计算,对其抗旱性进行评价。该平均值代表柳枝稷的抗旱性,数值越大表示抗旱性越强[20]。隶属函数的具体计算方法如下:
A:若指标与抗旱性呈正相关关系,则:
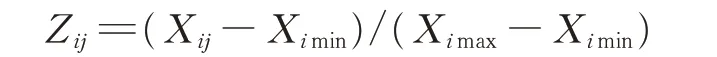
B:若指标与抗旱性呈负相关关系,则:
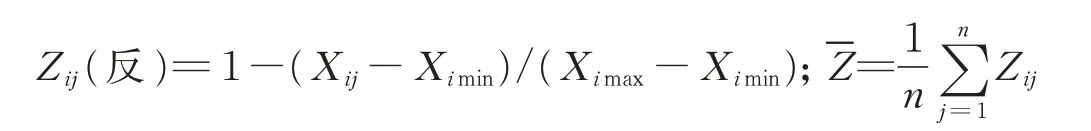
式中:i表示柳枝稷施氮水平,j表示抗旱指标,Zij为供试柳枝稷材料的抗旱性隶属函数值,Xij为各抗旱指标的实际测量值,Ximax为各抗旱指标实际测定的最大值,Ximin为各抗旱指标实际测定的最小值,n为指标数为隶属函数平均值。
1.5 数据处理
利用SPSS 23.0 软件进行数据分析,运用Excel 软件处理图表。各测定指标平均值采用单因素方差分析法,利用Duncan 法对相同处理下各指标进行差异显著性分析,显著水平为0.05。
2 结果与分析
2.1 柳枝稷叶片叶绿素相对含量的变化
柳枝稷各个生育时期内的叶片叶绿素相对含量(SPAD)值随着不同施氮水平的增加均呈现出先升后降的总趋势,其中在施中氮(N120)处理下达到峰值,分别为59.7(返青期)、63.0(拔节期)、69.0(抽穗期)、66.7(开花期)和54.9 mg·g−1(灌浆期),并且总体显著高于其他氮肥处理(图1)。其中,在柳枝稷开花期,施低氮(N60)、中氮(N120)和高氮(N240)处理下柳枝稷叶片SPAD 值之间均存在显著性差异,显著高于无氮添加(N0)处理(P<0.05),分别提高了3.69%、16.34%和8.08%。在柳枝稷返青期、抽穗期和灌浆期,施低氮(N60)和高氮(N240)处理之间SPAD 值变化不大,没有呈现出显著性差异(P>0.05)。在柳枝稷拔节期,无氮添加(N0)和施低氮(N60)处理之间,叶片SPAD 值之间没有显著性差异(P>0.05),但均显著低于施中氮(N120)和高氮(N240)处理(P<0.05)。施低氮(N60)、中氮(N120)和高氮(N240)处理较无氮添加(N0)处理分别提高了1.40%、18.87%和11.70%。
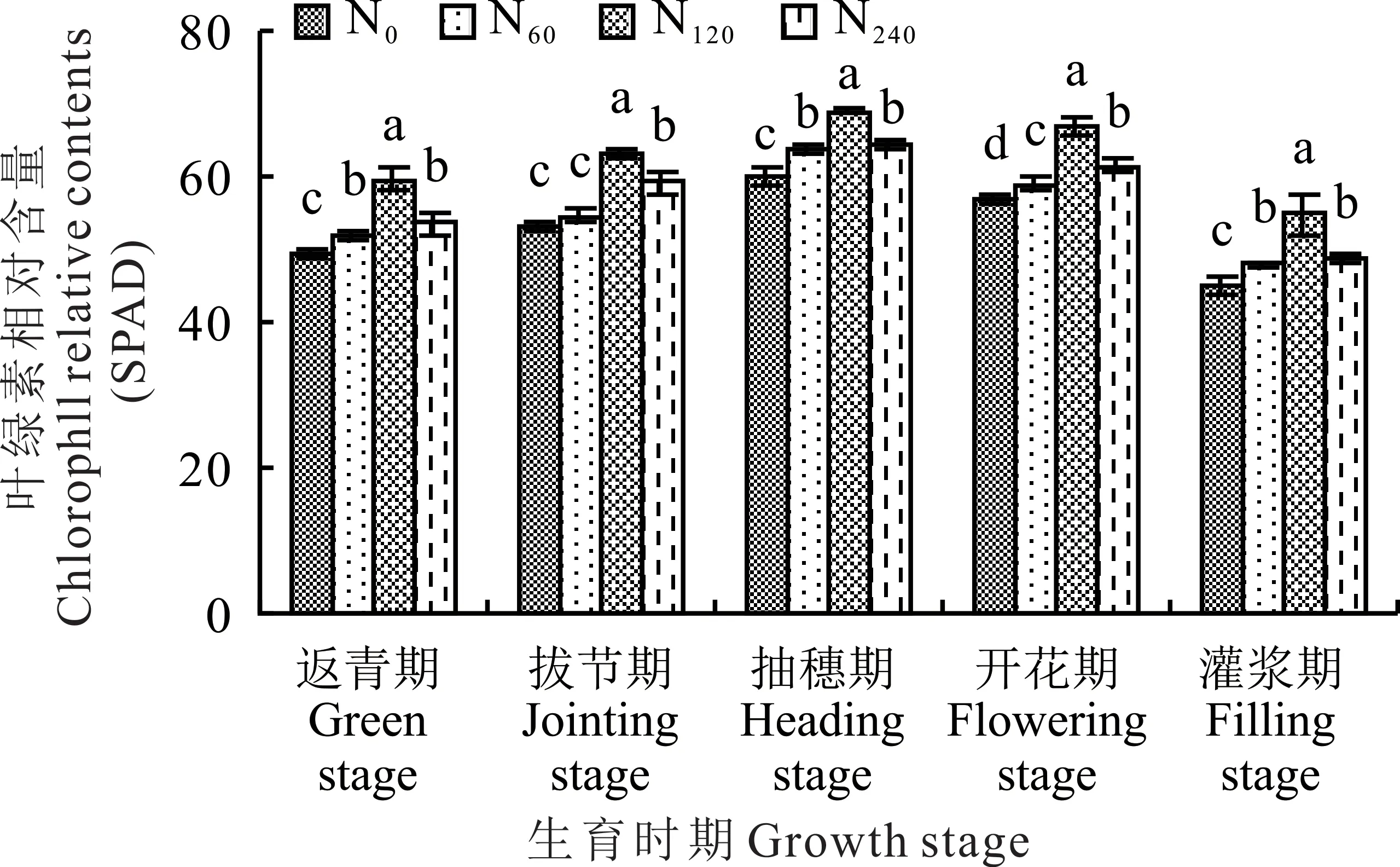
图1 不同施氮水平对柳枝稷叶片叶绿素相对含量的影响Fig.1 Effect of different nitrogen levels on chlorophyll relative content of switchgrass leaves
2.2 柳枝稷叶片净光合速率的变化
柳枝稷各个生育时期内叶片净光合速率在不同施氮水平呈现出先升后降的总趋势,在整个生育时期内,开花期柳枝稷叶片净光合速率最大(图2)。从氮肥处理来看,柳枝稷叶片净光合速率值均在施中氮(N120)处理下达到峰值,并且总体高于其他氮肥处理,在柳枝稷返青期、拔节期、抽穗期、开花期和灌浆期,净 光 合 速 率 分 别 为23.77、25.57、28.23、33.96 和31.30 μmol·m−2·s−1。在柳枝稷各生育时期,无氮添加(N0)和施低氮(N60)处理之间及施中氮(N120)和高氮(N240)处理之间,叶片净光合速率均没有显著性差异(P>0.05),但施中氮(N120)和高氮(N240)处理显著高于无氮添加(N0)和施低氮(N60)处理(P<0.05)。在柳枝稷整个生育时期内,施低氮(N60)、中氮(N120)和高氮(N240)处理较无氮添加平均提升了4.57%、13.66 %和11.99%。
2.3 柳枝稷叶片气孔导度的变化
总体来看,柳枝稷各生育时期内的气孔导度值均在N120处理下达到峰值,分别为0.095(返青期)、0.116(拔节期)、0.144(抽穗期)、0.178(开花期)和0.164 mol·m−2·s−1(灌浆期),总体上高于其他氮肥处理(图3)。在柳枝稷返青期、拔节期和开花期,无氮添加(N0)和施低氮(N60)处理之间及施中氮(N120)处理和高氮(N240)处理之间,柳枝稷叶片气孔导度均不存在显著性差异(P>0.05),但施中氮(N120)和施高氮(N240)处理显著高于无氮添加(N0)和施低氮(N60)处理(P<0.05)。在柳枝稷抽穗期和灌浆期,无氮添加(N0)和施低氮(N60)处理下柳枝稷叶片气孔导度之间没有显著性差异(P>0.05),但施中氮(N120)处理和高氮(N240)处理下气孔导度之间存在显著性差异(P<0.05)。在柳枝稷整个生育时期内,与无氮添加(N0)相比,施低氮(N60)、中氮(N120)处理和高氮(N240)处理下叶片气孔导度平均提高了5.49%、56.89%和39.52%。
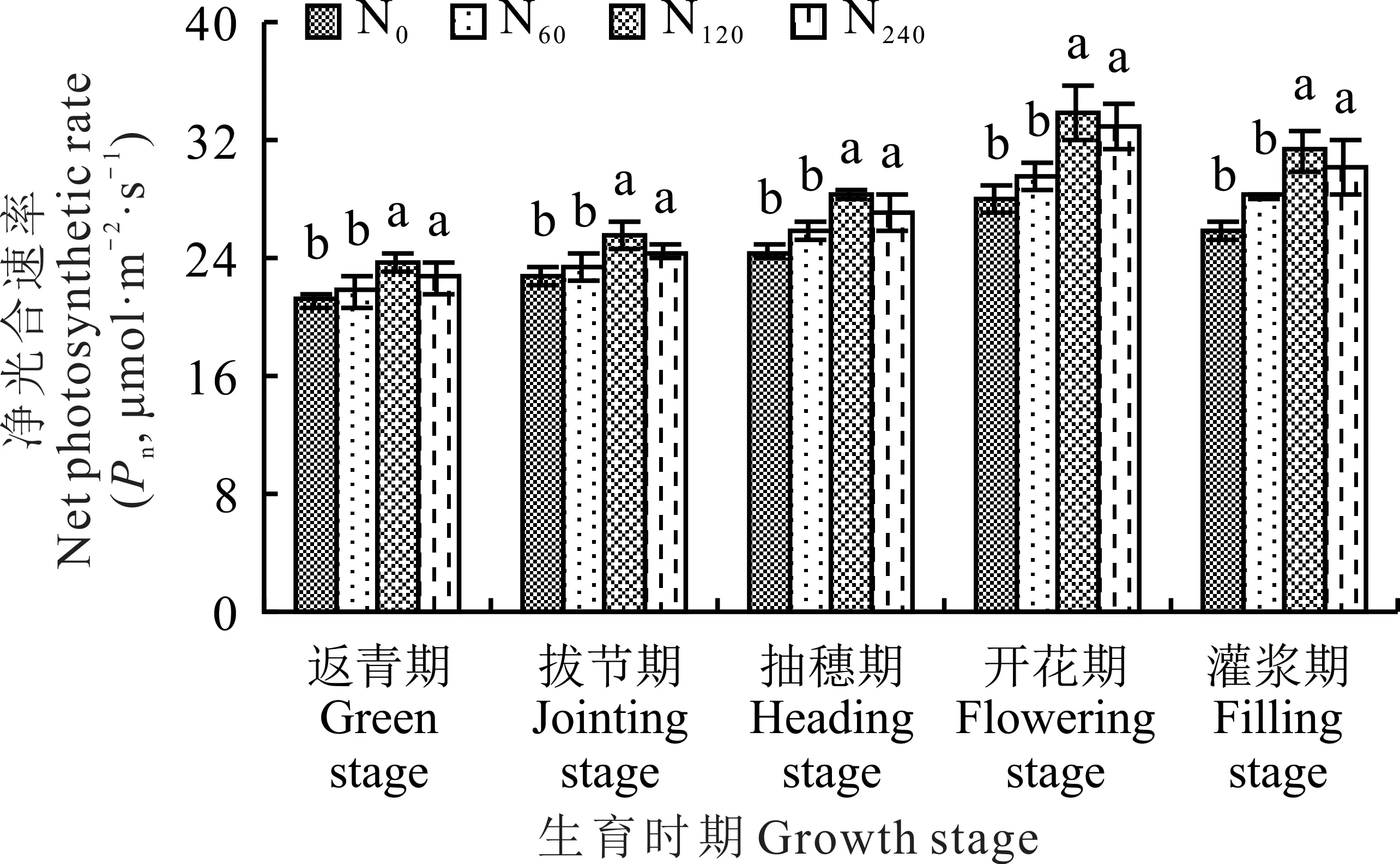
图2 不同施氮水平对柳枝稷叶片净光合速率的影响Fig.2 Effect of different nitrogen levels on net photosynthetic rate of switchgrass leaves
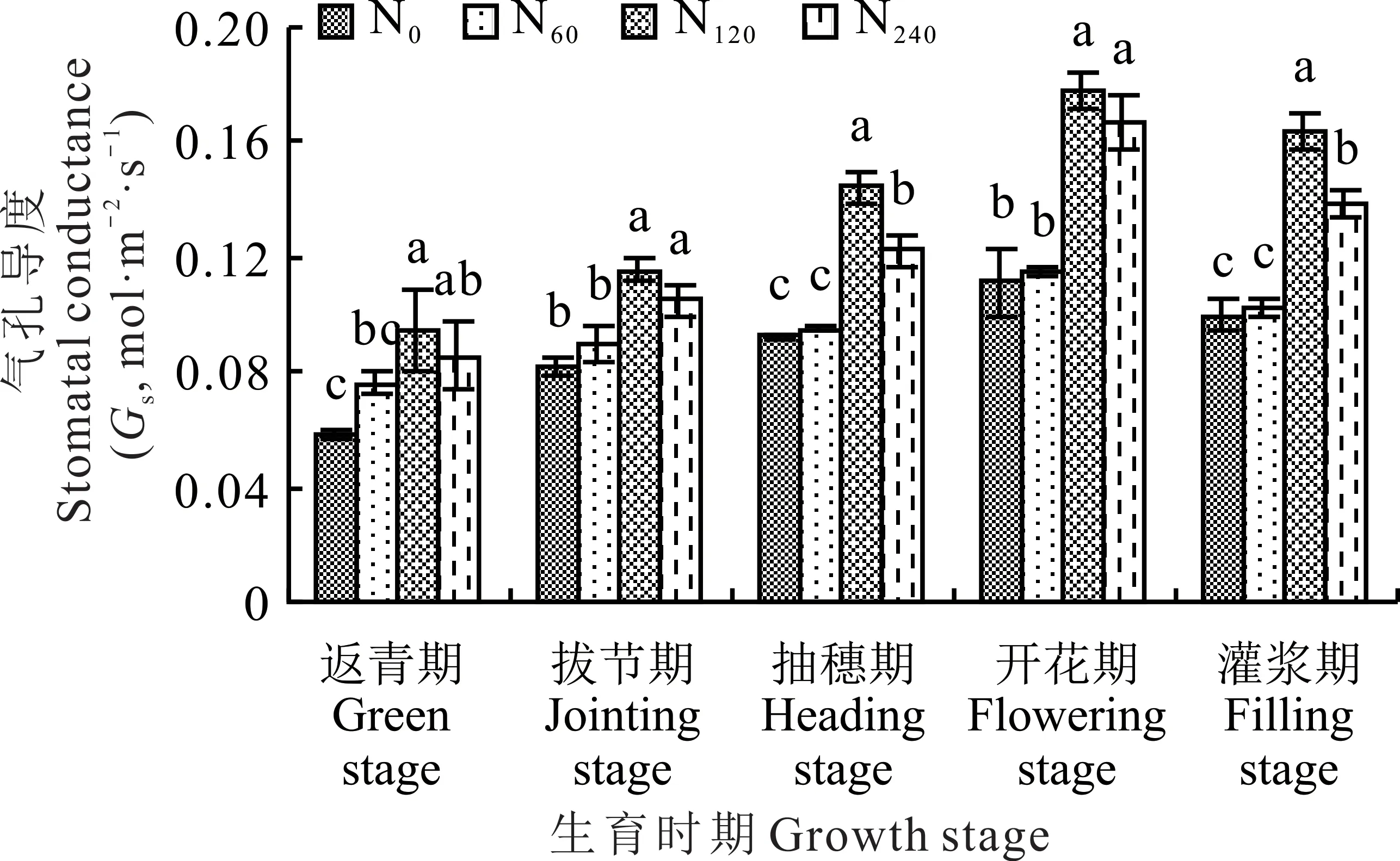
图3 不同施氮水平对柳枝稷叶片气孔导度的影响Fig.3 Effect of different nitrogen levels on stomatal conductance of switchgrass leaves
2.4 柳枝稷叶片细胞间隙CO2浓度的变化
不同施氮水平下柳枝稷各生育时期内叶片细胞间隙CO2浓度呈现出先升后降的总趋势,在整个生育时期内,开花期叶片细胞间隙CO2浓度最大(图4)。从氮肥处理来看,柳枝稷叶片细胞间隙CO2浓度均在施中氮(N120)处理下达到峰值,并且总体高于其他氮肥处理,在柳枝稷返青期、拔节期、抽穗期、开花期和灌浆期,叶片细胞间隙CO2浓度分别为174.61、210.56、219.86、328.53 和265.27 μmol·mol−1。在 拔节期和开花期,施中氮(N120)处理下柳枝稷叶片细胞间隙CO2浓度均显著高于其他氮肥处理(P<0.05)。在柳枝稷整个生育时期内,与无氮添加(N0)相比,施低氮(N60)、中氮(N120)和高氮(N240)处理下叶片细胞间隙CO2浓度平均提高了7.98%、28.97% 和11.44%。
2.5 柳枝稷叶片蒸腾速率的变化
柳枝稷各个生育时期内蒸腾速率均随着不同施氮水平的增加而呈现出递增趋势,其中在施高氮(N240)处理下最大,分别为3.94(返青期)、4.65(拔节期)、5.39(抽穗期)、6.70(开花期)和5.99 mmol·m−2·s−1(灌浆期),总体上高于其他氮肥处理(图5)。在柳枝稷返青期、拔节期和开花期,无氮添加(N0)和施低氮(N60)处理之间及施中氮(N120)处理和高氮(N240)处理之间均没有显著性差异(P>0.05)。在柳枝稷抽穗期和灌浆期,无氮添加(N0)和施低氮(N60)处理之间叶片的蒸腾速率均没有显著性差异(P>0.05),但均显著低于施中氮(N120)和高氮(N240)处理。在柳枝稷整个生育时期内,施低氮(N60)、中氮(N120)和高氮(N240)处理较无氮添加(N0)处理下叶片蒸腾速率平均提高了6.53%、25.66%和39.74%。
2.6 柳枝稷叶片水分利用效率的变化
柳枝稷叶片水分利用效率随着生育时期的推进而呈现出逐渐下降的总趋势(图6)。其中,返青期柳枝稷叶片的水分利用效率相对较大。从施氮水平来看,柳枝稷各个生育时期内的叶片水分利用效率均在施低氮(N60)处理下达到峰值,分别为7.02(返青期)、6.74(拔节期)、6.81(抽穗期)、5.96(开花期)和6.38 mg·g−1(灌浆期),整体上高于其他氮肥处理,并且随着不同施氮水平的增加而逐渐降低。在柳枝稷返青期和开花期,无氮添加(N0)、施低氮(N60)和中氮(N120)处理之间叶片水分利用效率均没有显著性差异(P>0.05)。在柳枝稷拔节期、抽穗期和灌浆期,无氮添加(N0)和施中氮(N120)处理之间叶片水分利用效率也没有显著性差异(P>0.05)。在柳枝稷生育时期内,与无氮添加(N0)相比,施低氮(N60)处理下叶片水分利用效率分别提高了2.63%、8.75%、8.96%、6.05%和8.70%。
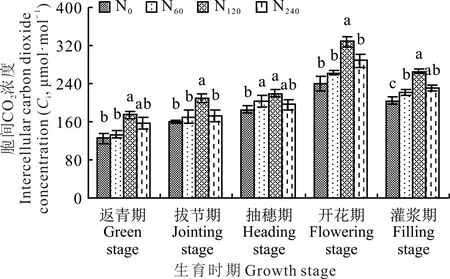
图4 不同施氮水平对柳枝稷叶片细胞间隙CO2浓度的影响Fig.4 Effect of different nitrogen levels on CO2 concentration in the intercellular space of switchgrass leaves
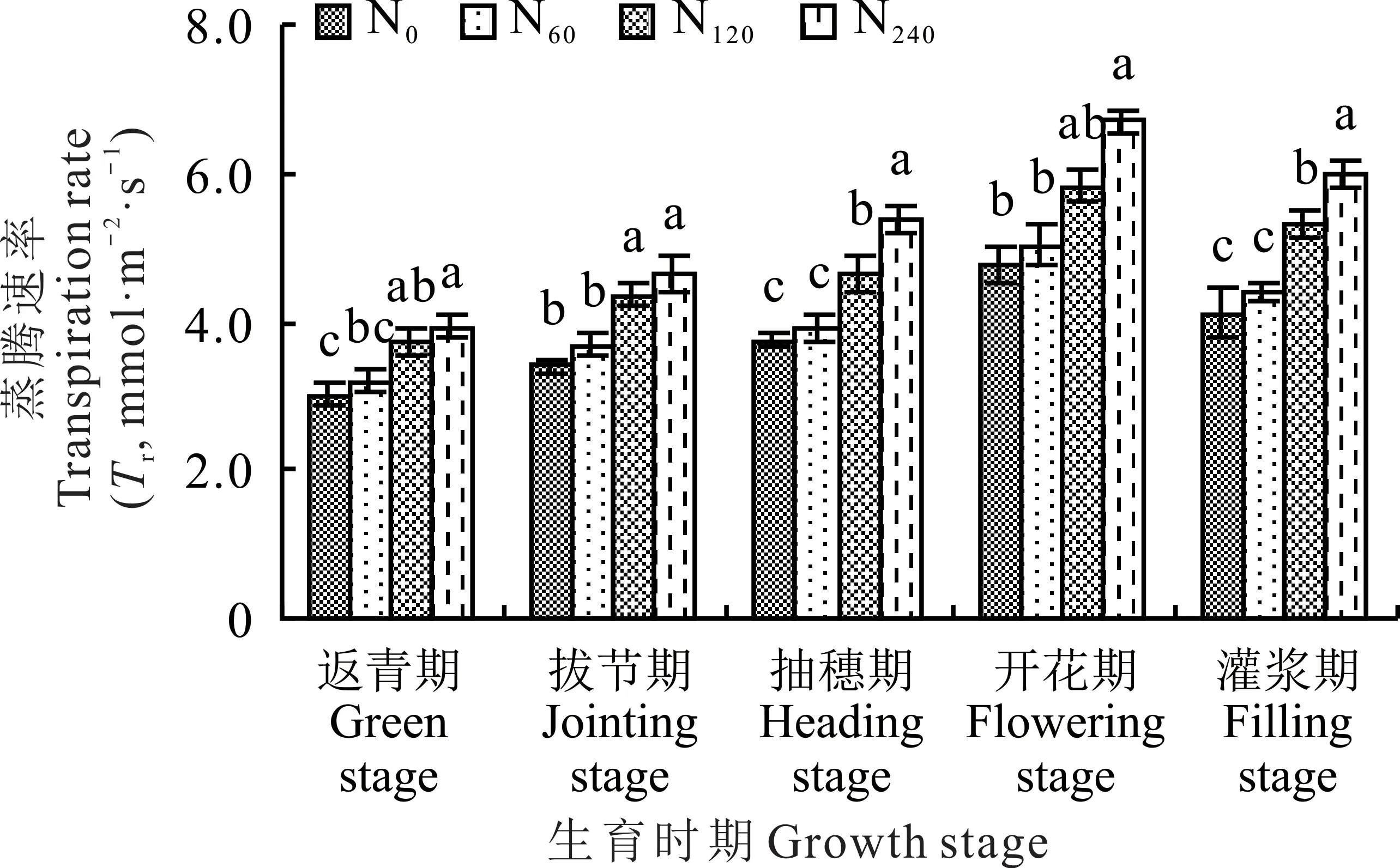
图5 不同施氮水平对柳枝稷叶片蒸腾速率的影响Fig.5 Effect of different nitrogen levels on transpiration rate of switchgrass leaves
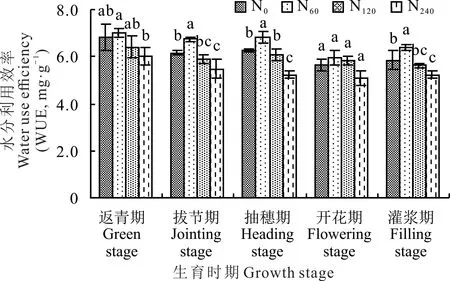
图6 不同施氮水平对柳枝稷叶片水分利用效率的影响Fig.6 Effect of different nitrogen levels on water use efficiency of switchgrass leaves
2.7 不同施氮水平对柳枝稷抗旱性的影响
柳枝稷叶片中主要渗透调节物质及抗氧化酶活性均随着不同施氮水平的增加而不同程度地升高,其中SOD、POD、Pro、MDA、SS 及相对电导率均在N240处理下最大,分别为471.17 U·g−1·min−1、24671.32 U·g−1·min−1、83.55 μg·g−1、0.97 μg·g−1、0.88 mg·g−1和0.87,而CAT 活性在N120处理下最大,为177.08 U·g−1·min−1,之后开始降低(表1)。与N0处理相比,SOD 活性随着不同施氮水平的增加而显著性升高(P<0.05),N60、N120和N240处理分别提高了26.02%、38.18%和48.84%;POD 活性随着不同施氮水平的增加而逐渐升高,且N60和N120处理之间差异不 显著(P>0.05)。N60、N120和N240处理较N0处理POD 活 性分别提高了9.26%、14.64% 和33.79%;CAT 活性随着不同施氮水平的增加而呈现先上升后下降的趋势,且各施氮处理之间差异显著(P<0.05),与N0处理相比,N60、N120和N240处理分别提高了22.52%、47.19%和14.72%;Pro 含量随着不同施氮水平的增加而显著增加,且N0和N60处理之间差异不显著(P>0.05),与N0处理相比,N120和N240处理Pro 含量分别提高了6.61%和18.70%;MDA 和SS 含量均随着不同施氮水平的增加而显著性提高(P<0.05),N60、N120和N240处理较N0处理MDA 含量分别提高了7.59%、11.39%和22.78%,SS 含量分别提高了13.21%、52.83%和66.04%;相对电导率随着不同施氮水平的增加而逐渐升高,其中N60和N120处理之间柳枝稷叶片相对电导率没有显著性差异(P>0.05),与N0处理相比,N60、N120和N240处理柳枝稷叶片相对电导率分别提高了2.50%、3.75%和8.75%。
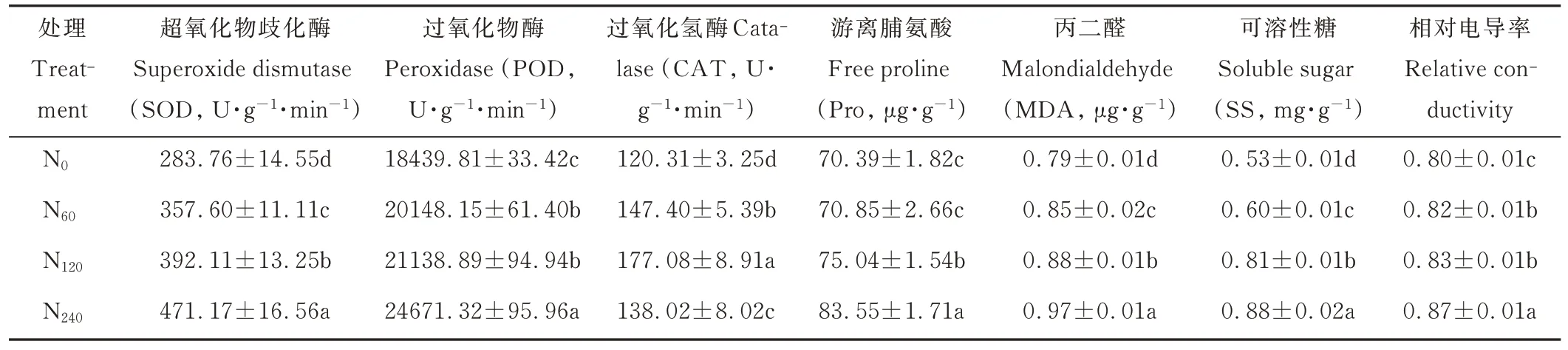
表1 不同施氮水平对柳枝稷主要渗透调节物质和抗氧化酶活性的影响Table 1 Effects of different nitrogen levels on main osmolytes and antioxidant enzyme activities of switchgrass
盐碱条件下柳枝稷各抗旱指标隶属函数值的平均值随着不同施氮水平的增加呈现出先上升后下降的趋势,其中施氮水平为120 kg·hm−2(N120)时,柳枝稷各抗旱指标隶属函数值的平均值最大,为0.510。其余各施氮水平下,柳枝稷各抗旱指标隶属函数值的平均值从大到小依次为:N60>N240>N0,即适当的增施氮肥能在一定程度上提高盐碱地柳枝稷的抗旱性(表2)。
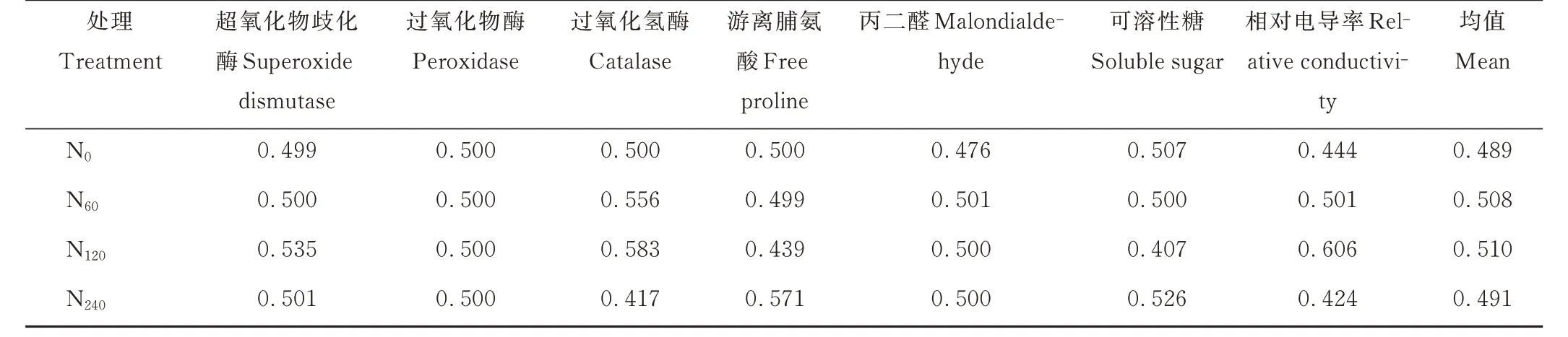
表2 不同施氮水平对柳枝稷各抗旱指标隶属函数值的影响Table 2 Effect of different nitrogen levels amount on the drought resistance index membership function value of switchgrass
3 讨论
光合作用是植物生长的基础,也是绿色植物最主要的生理生化过程之一。盐碱及干旱胁迫条件下,光合性能的强弱直接影响植物的生长发育,也是评价植物耐盐碱及抗旱能力的重要生理指标。氮素直接影响植物叶绿素含量并参与光合作用中与光呼吸相关酶合成与活性,相对叶绿素含量的高低直接影响着植物的光合速率,易受环境条件的改变而改变[20−21]。本研究发现,在柳枝稷各生育时期内,随着施氮量的增加叶片的相对叶绿素含量呈现先增加再降低的变化趋势,中氮(120 kg·hm−2,N120)处理下叶片的SPAD 值达到峰值,高氮(240 kg·hm−2,N240)处理下反而降低。说明在合理的施氮范围内,增施氮肥能有效提高柳枝稷的叶绿素相对含量,而过量的氮供应会使作物叶片呈深绿色,导致营养生长过盛,生殖生长相对减弱,叶片的相对叶绿素含量反而降低。氮肥的合理运筹是当前农业生产中土壤和作物管理的重点,也是大多数农业生态系统的主要限制性因子,氮元素主要参与作物的光合作用,对提高作物光合性能具有重要意义[22−23]。结果表明,施中氮(120 kg·hm−2,N120)时提高了柳枝稷的净光合速率(Pn)、气孔导度(Gs)、细胞间隙CO2浓度(Ci)和蒸腾速率(Tr)等光合指标,进而提高干物质积累。其主要原因是合理的氮肥运筹可以协调柳枝稷生长期的需氮特性,使得柳枝稷生育后期叶片具有较强的光捕获能力。这与王茜等[24]研究不同土壤水分条件下施氮水平对紫花苜蓿(Medicago sativa)苗期光合作用的影响结果基本一致。同时研究发现,柳枝稷叶片的蒸腾速率在施高氮(240 kg·hm−2,N240)处理下达到最大值,其主要原因是作物在光下进行光合作用,经由气孔吸收CO2,气孔必须张开,并可以根据外界环境条件的变化来调节自己的开度而保证获得最高的光合性能。气孔开张又不可避免的发生蒸腾作用,这与王进斌等[25]研究氮肥运筹对陇中旱农区玉米(Zea mays)光合特性及产量的影响结果基本一致。
研究发现,适当地追施氮肥不仅能够显著提高作物的净光合速率,而且可以提高植株叶片光合机构对逆境胁迫的适应能力,同时增加了气孔对逆境胁迫的敏感程度,降低了盐碱地植物对逆境胁迫的敏感程度[26]。施氮对植物抗旱性影响的研究结果各异,有研究认为增施氮肥显著提高小麦(Triticum aestivum)叶片在干旱环境中的保水能力,从而提高其对干旱的适应能力[27−28];张岁歧等[29]认为增施氮肥不能提高春小麦对干旱的适应能力,但是改变了其对干旱的适应机制。当植物受到盐碱及干旱胁迫时,自身可通过合成一些渗透调节物质、提高抗氧化酶活性来抵御不良环境。游离脯氨酸(Pro)、丙二醛(MDA)、可溶性糖(SS)、超氧化物歧化酶(SOD)和过氧化物酶(POD)等都是植物体内常见的渗透调节物质及主要的抗氧化酶,对于维持渗透平衡、保护细胞结构均具有重要意义[30−31]。本研究中4 种施氮水平下柳枝稷叶片中SOD、POD、Pro、MDA 和SS 含量均随着施氮量的增加出现大幅显著升高现象,说明在此胁迫下柳枝稷通过自身合成这些渗透调节物质来抵御逆境胁迫,这与刘兵等[32]对盐碱胁迫下垂丝海棠(Malus halliana)的研究结果一致。同时通过隶属函数分析发现,施中氮(120 kg·hm−2,N120)处理下柳枝稷的抗旱性最强,高氮(240 kg·hm−2,N240)处理下柳枝稷的抗旱性反而会有所降低,说明适当增施氮肥能提高作物总根重和深层土壤根重,改善了根系的水分关系,提高了细胞膜的稳定性,有助于提高作物的抗旱性,而过量的氮供应,又会使作物叶片呈深绿色,植株茎干脆弱,导致营养生长和生殖生长失衡,反而会降低其抗旱性,这与朱毅等[33]研究柳枝稷苗期对氮素胁迫耐受性的综合评价结果基本一致。
4 结论
盐碱条件下,适当的增施氮肥不仅能提高柳枝稷叶片的光合作用,还能在一定程度上提高柳枝稷的抗旱性。随着施氮水平的增加,光合作用和抗旱性整体呈上升趋势,但过量施氮会导致柳枝稷叶片的光合作用和抗旱性相对下降。本试验条件下,施中氮(120 kg·hm−2,N120)处理既可以延缓柳枝稷叶片衰老,提升叶片光合能力,又能在一定程度上提高柳枝稷的抗旱性,是较为理想的施氮水平。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
中国农学通报(2022年29期)2022-11-25
——现代牙刷的由来
童话王国·文学大师班(2022年9期)2022-10-08
小学生必读(低年级版)(2021年4期)2021-07-28
文苑(2020年2期)2020-11-22
文苑·经典美文(2020年2期)2020-04-19
农业环境科学学报(2017年2期)2017-03-20
甘肃林业科技(2016年2期)2016-11-16
河南农业大学学报(2016年2期)2016-09-26
农家顾问(2014年9期)2014-10-20
